
Staurosporine as a Potential Treatment for Acanthamoeba Keratitis Using Mouse Cornea as an Ex Vivo Model

 ,
,  ,
,  ,
,  , ,
, ,  and
and
Abstract

1. Introduction
2. Results
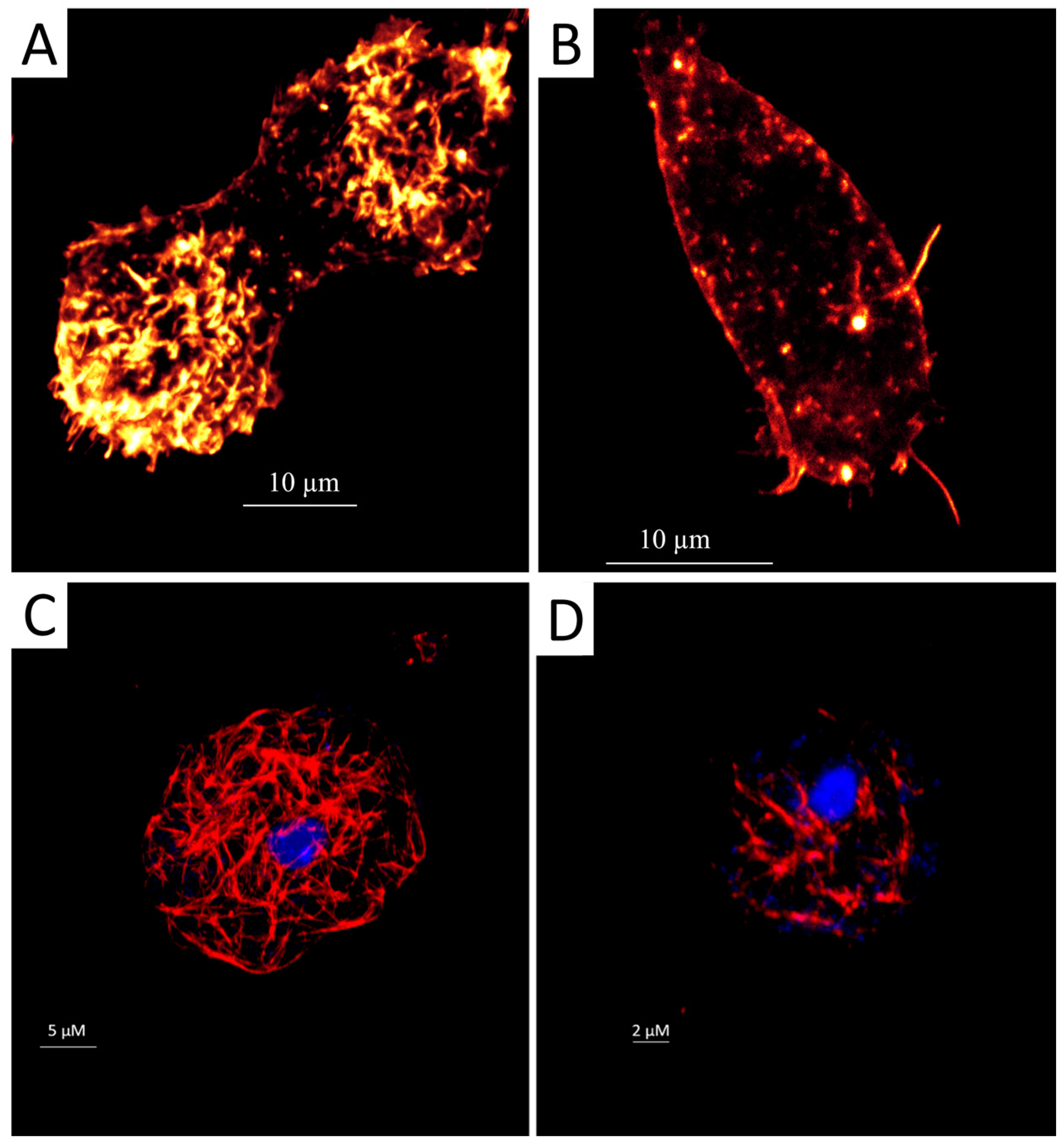
2.1. Staurosporine Induced Structural Damage of the Acanthamoeba Cytoskeleton
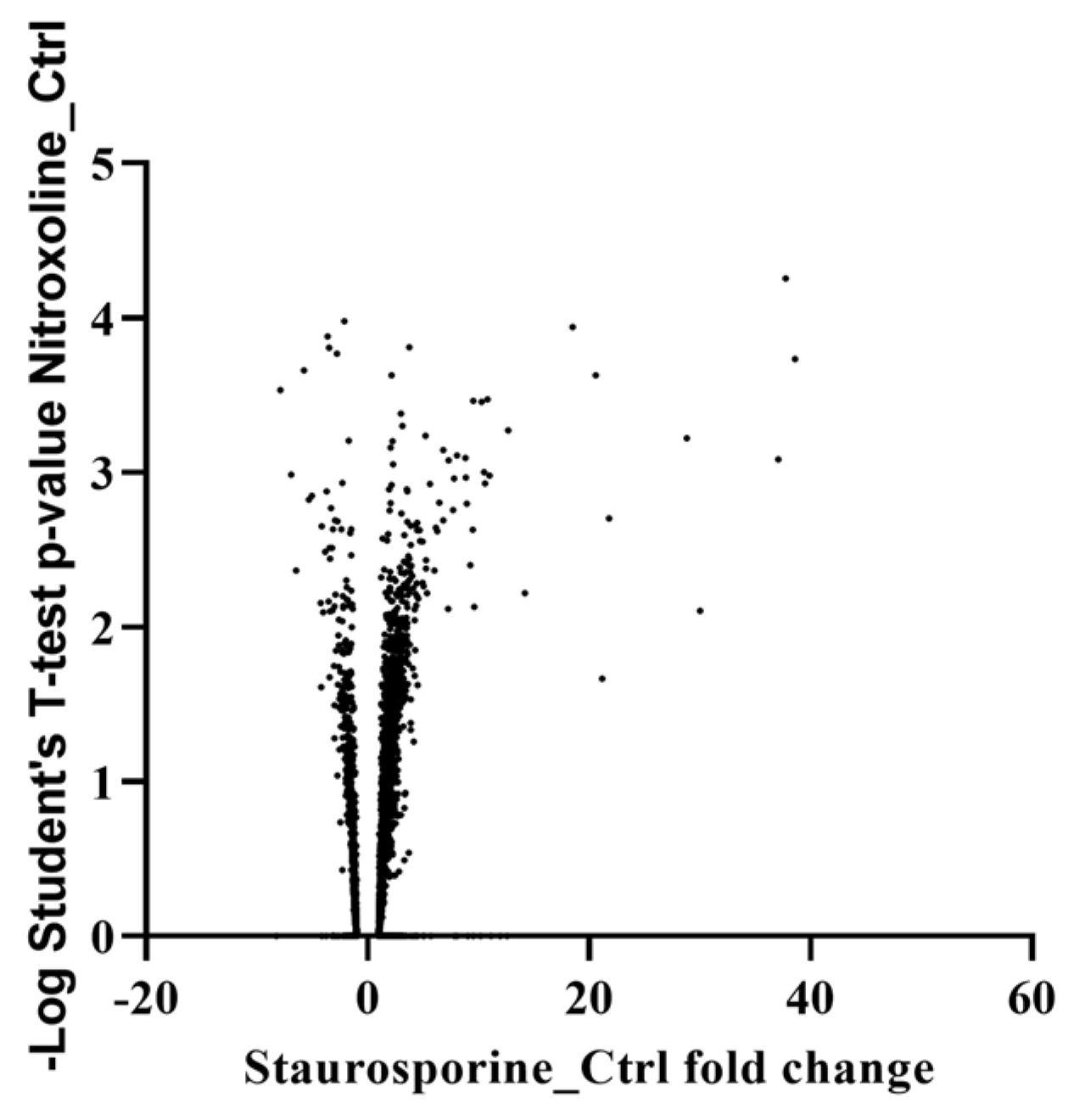
2.2. Proteomic Analysis
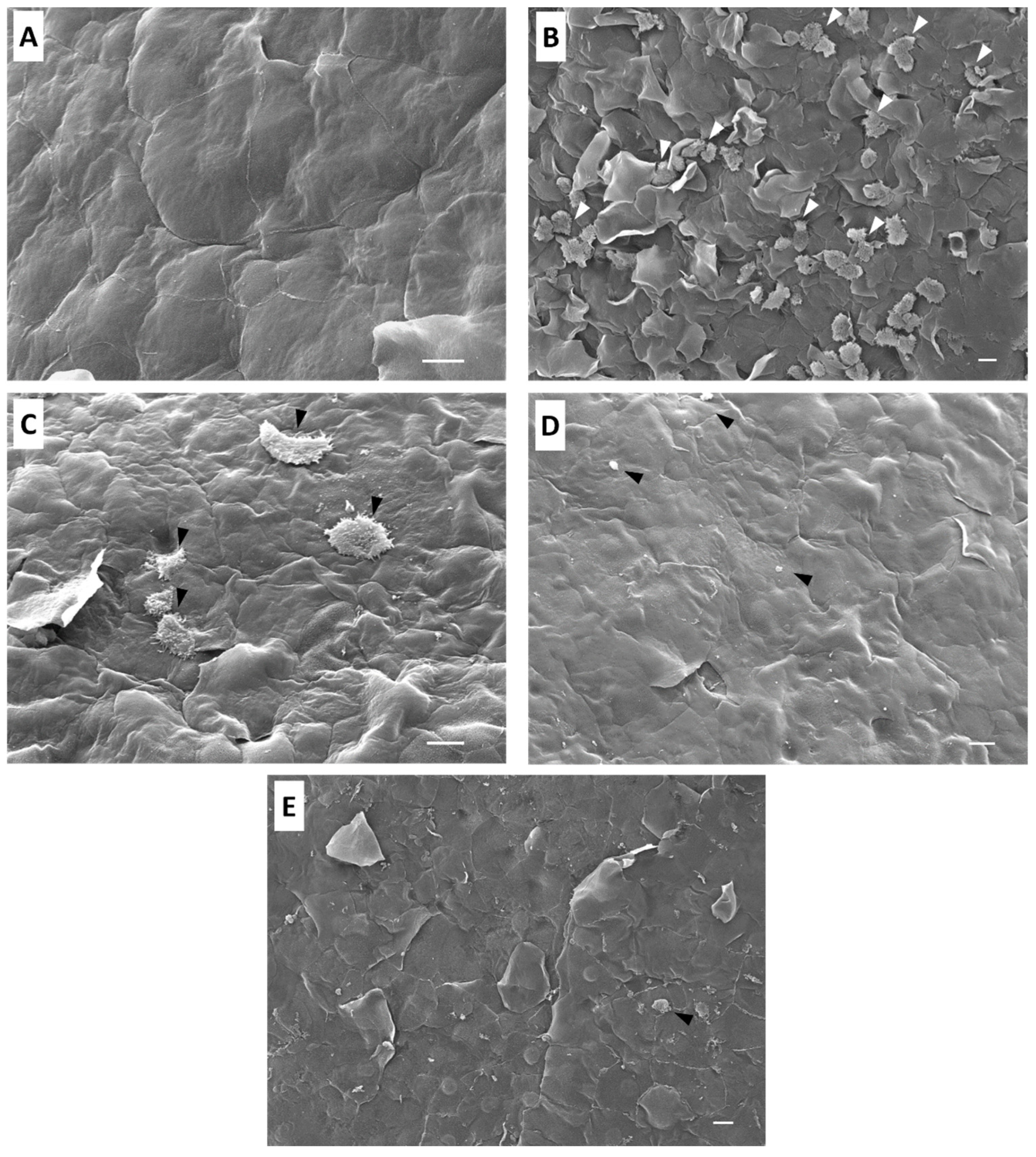
2.3. Ex Vivo Assay in Mouse Corneas Infected with A. castellanii and Treated with Staurosporine
3. Discussion
4. Materials and Methods
4.1. Acanthamoeba Strain
4.2. In Vitro Assay
4.2.1. Fluorescent Staining of Actin Distribution
4.2.2. Immunofluorescence Staining of the Tubulin of Acanthamoeba Trophozoites
4.3. Proteomic Analysis
4.4. Ex Vivo Infection
Co-Incubation and Interaction of A. castellanii Trophozoites with BALB/c Mice Cornea
4.5. Scanning Electron Microscopy
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Correction Statement
References
- Henriquez, F.L. Review of “Acanthamoeba: Biology and Pathogenesis” by Naveed Ahmed Khan. Parasit. Vectors 2009, 2, 16. [Google Scholar] [CrossRef]
- Lorenzo-Morales, J.; Martín-Navarro, C.M.; López-Arencibia, A.; Arnalich-Montiel, F.; Piñero, J.E.; Valladares, B. Acanthamoeba Keratitis: An Emerging Disease Gathering Importance Worldwide? Trends Parasitol. 2013, 29, 181–187. [Google Scholar] [CrossRef] [PubMed]
- List, W.; Glatz, W.; Riedl, R.; Mossboeck, G.; Steinwender, G.; Wedrich, A. Evaluation of Acanthamoeba Keratitis Cases in a Tertiary Medical Care Centre over 21 Years. Sci. Rep. 2021, 11, 1036. [Google Scholar] [CrossRef] [PubMed]
- Aiello, F.; Afflitto, G.G.; Ceccarelli, F.; Turco, M.V.; Han, Y.; Amescua, G.; Dart, J.K.; Nucci, C. Perspectives on the Incidence of Acanthamoeba Keratitis: A Systematic Review and Meta-Analysis. Ophthalmology 2024. [Google Scholar] [CrossRef] [PubMed]
- Petrillo, F.; Tortori, A.; Vallino, V.; Galdiero, M.; Fea, A.M.; De Sanctis, U.; Reibaldi, M. Understanding Acanthamoeba Keratitis: An In-Depth Review of a Sight-Threatening Eye Infection. Microorganisms 2024, 12, 758. [Google Scholar] [CrossRef]
- Jasim, H.; Grzeda, M.; Foot, B.; Tole, D.; Hoffman, J.J. Incidence of Acanthamoeba Keratitis in the United Kingdom in 2015: A Prospective National Survey. Cornea 2024, 43, 269–276. [Google Scholar] [CrossRef]
- Fanselow, N.; Sirajuddin, N.; Yin, X.-T.; Huang, A.J.W.; Stuart, P.M. Acanthamoeba Keratitis, Pathology, Diagnosis and Treatment. Pathogens 2021, 10, 323. [Google Scholar] [CrossRef]
- Szentmáry, N.; Daas, L.; Shi, L.; Laurik, K.L.; Lepper, S.; Milioti, G.; Seitz, B. Acanthamoeba Keratitis—Clinical Signs, Differential Diagnosis and Treatment. J. Curr. Ophthalmol. 2019, 31, 16–23. [Google Scholar] [CrossRef]
- Lorenzo-Morales, J.; Khan, N.A.; Walochnik, J. An Update on Acanthamoeba Keratitis: Diagnosis, Pathogenesis and Treatment. Parasite 2015, 22, 10. [Google Scholar] [CrossRef]
- Posarelli, M.; Passaro, M.L.; Avolio, F.C.; Costagliola, C.; Semeraro, F.; Romano, V. The Incidence of Severe Complications in Acanthamoeba Keratitis: Qualitative and Quantitative Systematic Assessment. Surv. Ophthalmol. 2024, 69, 769–778. [Google Scholar] [CrossRef]
- Nakano, H.; Ōmura, S. Chemical Biology of Natural Indolocarbazole Products: 30 Years since the Discovery of Staurosporine. J. Antibiot. 2009, 62, 17–26. [Google Scholar] [CrossRef] [PubMed]
- Omura, S.; Iwai, Y.; Hirano, A.; Nakagawa, A.; Awaya, J.; Tsuchiya, H.; Takahashi, Y.; Asuma, R. A New Alkaloid AM-2282 of Streptomyces Origin Taxonomy, Fermentation, Isolation and Preliminary Characterization. J. Antibiot. 1977, 30, 275–282. [Google Scholar] [CrossRef] [PubMed]
- Gani, O.A.B.S.M.; Engh, R.A. Protein Kinase Inhibition of Clinically Important Staurosporine Analogues. Nat. Prod. Rep. 2010, 27, 489. [Google Scholar] [CrossRef] [PubMed]
- Jung, H.; Zarlenga, D.; Martin, J.C.; Geldhof, P.; Hallsworth-Pepin, K.; Mitreva, M. The Identification of Small Molecule Inhibitors with Anthelmintic Activities That Target Conserved Proteins among Ruminant Gastrointestinal Nematodes. mBio 2024, 15, e00095-24. [Google Scholar] [CrossRef] [PubMed]
- Cartuche, L.; Sifaoui, I.; López-Arencibia, A.; Bethencourt-Estrella, C.J.; San Nicolás-Hernández, D.; Lorenzo-Morales, J.; Piñero, J.E.; Díaz-Marrero, A.R.; Fernández, J.J. Antikinetoplastid Activity of Indolocarbazoles from Streptomyces Sanyensis. Biomolecules 2020, 10, 657. [Google Scholar] [CrossRef] [PubMed]
- Pimentel-Elardo, S.M.; Kozytska, S.; Bugni, T.S.; Ireland, C.M.; Moll, H.; Hentschel, U. Anti-Parasitic Compounds from Streptomyces Sp. Strains Isolated from Mediterranean Sponges. Mar. Drugs 2010, 8, 373–380. [Google Scholar] [CrossRef]
- Cartuche, L.; Reyes-Batlle, M.; Sifaoui, I.; Arberas-Jiménez, I.; Piñero, J.E.; Fernández, J.J.; Lorenzo-Morales, J.; Díaz-Marrero, A.R. Antiamoebic Activities of Indolocarbazole Metabolites Isolated from Streptomyces Sanyensis Cultures. Mar. Drugs 2019, 17, 588. [Google Scholar] [CrossRef]
- Cartuche, L.; Sifaoui, I.; Cruz, D.; Reyes-Batlle, M.; López-Arencibia, A.; Javier Fernández, J.; Díaz-Marrero, A.R.; Piñero, J.E.; Lorenzo-Morales, J. Staurosporine from Streptomyces Sanyensis Activates Programmed Cell Death in Acanthamoeba via the Mitochondrial Pathway and Presents Low in Vitro Cytotoxicity Levels in a Macrophage Cell Line. Sci. Rep. 2019, 9, 11651. [Google Scholar] [CrossRef]
- Dusinska, M.; Rundén-Pran, E.; Schnekenburger, J.; Kanno, J. Toxicity Tests: In Vitro and In Vivo. In Adverse Effects of Engineered Nanomaterials; Elsevier: Amsterdam, The Netherlands, 2017; pp. 51–82. [Google Scholar]
- Ren, M.; Wu, X. Evaluation of Three Different Methods to Establish Animal Models of Acanthamoeba Keratitis. Yonsei Med. J. 2010, 51, 121. [Google Scholar] [CrossRef]
- Dwia Pertiwi, Y.; Chikama, T.; Sueoka, K.; Ko, J.-A.; Kiuchi, Y.; Onodera, M.; Sakaguchi, T. Efficacy of Photodynamic Anti-Microbial Chemotherapy for Acanthamoeba Keratitis In Vivo. Lasers Surg. Med. 2021, 53, 695–702. [Google Scholar] [CrossRef]
- González-Robles, A.; Omaña-Molina, M.; Salazar-Villatoro, L.; Flores-Maldonado, C.; Lorenzo-Morales, J.; Reyes-Batlle, M.; Arnalich-Montiel, F.; Martínez-Palomo, A. Acanthamoeba culbertsoni Isolated from a Clinical Case with Intraocular Dissemination: Structure and In Vitro Analysis of the Interaction with Hamster Cornea and MDCK Epithelial Cell Monolayers. Exp. Parasitol. 2017, 183, 245–253. [Google Scholar] [CrossRef] [PubMed]
- Omaña-Molina, M.; Navarro-García, F.; González-Robles, A.; Serrano-Luna, J.d.J.; Campos-Rodríguez, R.; Martínez-Palomo, A.; Tsutsumi, V.; Shibayama, M. Induction of Morphological and Electrophysiological Changes in Hamster Cornea after in Vitro Interaction with Trophozoites of Acanthamoeba spp. Infect. Immun. 2004, 72, 3245–3251. [Google Scholar] [CrossRef] [PubMed]
- Omaña-Molina, M.; González-Robles, A.; Iliana Salazar-Villatoro, L.; Lorenzo-Morales, J.; Cristóbal-Ramos, A.R.; Hernández-Ramírez, V.I.; Talamás-Rohana, P.; Méndez Cruz, A.R.; Martínez-Palomo, A. Reevaluating the Role of Acanthamoeba Proteases in Tissue Invasion: Observation of Cytopathogenic Mechanisms on MDCK Cell Monolayers and Hamster Corneal Cells. Biomed. Res. Int. 2013, 2013, 461329. [Google Scholar] [CrossRef]
- Salazar-Villatoro, L.; Chávez-Munguía, B.; Guevara-Estrada, C.E.; Lagunes-Guillén, A.; Hernández-Martínez, D.; Castelan-Ramírez, I.; Omaña-Molina, M. Taurine, a Component of the Tear Film, Exacerbates the Pathogenic Mechanisms of Acanthamoeba castellanii in the Ex Vivo Amoebic Keratitis Model. Pathogens 2023, 12, 1049. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.-J.; Chen, C.-H.; Chen, J.-W.; Lin, W.-C. Commensals Serve as Natural Barriers to Mammalian Cells during Acanthamoeba castellanii Invasion. Microbiol. Spectr. 2021, 9, e00512-21. [Google Scholar] [CrossRef] [PubMed]
- Teuchner, B.; Wibmer, I.D.; Schaumann, P.; Seifarth, C.; Walochnik, J.; Nagl, M. N-Chlorotaurine Inactivates Acanthamoeba and Candida Albicans in the Porcine Ex Vivo Corneal Infection Model. Cornea 2019, 38, 1011–1016. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Xu, S.; Wan, J.; Wang, D.; Pang, X.; Gao, Y.; Ni, N.; Chen, D.; Sun, X. Disturbing Cytoskeleton by Engineered Nanomaterials for Enhanced Cancer Therapeutics. Bioact. Mater. 2023, 29, 50–71. [Google Scholar] [CrossRef]
- Henriquez, F.L.; Ingram, P.R.; Muench, S.P.; Rice, D.W.; Roberts, C.W. Molecular Basis for Resistance of Acanthamoeba Tubulins to All Major Classes of Antitubulin Compounds. Antimicrob. Agents Chemother. 2008, 52, 1133–1135. [Google Scholar] [CrossRef]
- Omura, S.; Sasaki, Y.; Iwai, Y.; Takeshima, H. Staurosporine, a Potentially Important Gift from a Microorganism. J. Antibiot. 1995, 48, 535–548. [Google Scholar] [CrossRef]
- Hedberg, K.K.; Birrell, G.B.; Habliston, D.L.; Griffith, O.H. Staurosporine Induces Dissolution of Microfilament Bundles by a Protein Kinase C-Independent Pathway. Exp. Cell Res. 1990, 188, 199–208. [Google Scholar] [CrossRef]
- Xie, J.L.; O’Meara, T.R.; Polvi, E.J.; Robbins, N.; Cowen, L.E. Staurosporine Induces Filamentation in the Human Fungal Pathogen Candida Albicans via Signaling through Cyr1 and Protein Kinase A. mSphere 2017, 2, e00056-17. [Google Scholar] [CrossRef] [PubMed]
- Olguín-Albuerne, M.; Domínguez, G.; Morán, J. Effect of Staurosporine in the Morphology and Viability of Cerebellar Astrocytes: Role of Reactive Oxygen Species and NADPH Oxidase. Oxid. Med. Cell Longev. 2014, 2014, 678371. [Google Scholar] [CrossRef] [PubMed]
- Hong, Y.; Kang, J.-M.; Joo, S.-Y.; Song, S.-M.; Lê, H.G.; Thái, T.L.; Lee, J.; Goo, Y.-K.; Chung, D.-I.; Sohn, W.-M.; et al. Molecular and Biochemical Properties of a Cysteine Protease of Acanthamoeba castellanii. Korean J. Parasitol. 2018, 56, 409–418. [Google Scholar] [CrossRef] [PubMed]
- Saurav, S.; Manna, S.K. Profilin Upregulation Induces Autophagy through Stabilization of AMP-activated Protein Kinase. FEBS Lett. 2022, 596, 1765–1777. [Google Scholar] [CrossRef]
- Pernier, J.; Shekhar, S.; Jegou, A.; Guichard, B.; Carlier, M.-F. Profilin Interaction with Actin Filament Barbed End Controls Dynamic Instability, Capping, Branching, and Motility. Dev. Cell 2016, 36, 201–214. [Google Scholar] [CrossRef]
- Omaña-Molina, M.; González-Robles, A.; Salazar-Villatoro, L.I.; Cristóbal-Ramos, A.R.; González-Lázaro, M.; Salinas-Moreno, E.; Méndez-Cruz, R.; Sánchez-Cornejo, M.; De la Torre-González, E.; Martínez-Palomo, A. Acanthamoeba castellanii: Morphological Analysis of the Interaction with Human Cornea. Exp. Parasitol. 2010, 126, 73–78. [Google Scholar] [CrossRef]
- Shing, B.; Balen, M.; McKerrow, J.H.; Debnath, A. Acanthamoeba Keratitis: An Update on Amebicidal and Cysticidal Drug Screening Methodologies and Potential Treatment with Azole Drugs. Expert. Rev. Anti Infect. Ther. 2021, 19, 1427–1441. [Google Scholar] [CrossRef]
- Megha, K.; Sharma, M.; Sharma, C.; Gupta, A.; Sehgal, R.; Khurana, S. Evaluation of in Vitro Activity of Five Antimicrobial Agents on Acanthamoeba Isolates and Their Toxicity on Human Corneal Epithelium. Eye 2022, 36, 1911–1917. [Google Scholar] [CrossRef]
- Wang, Y.; Jiang, L.; Zhao, Y.; Ju, X.; Wang, L.; Jin, L.; Fine, R.D.; Li, M. Biological Characteristics and Pathogenicity of Acanthamoeba. Front. Microbiol. 2023, 14, 1147077. [Google Scholar] [CrossRef]
- Liu, J.X.; Werner, J.; Kirsch, T.; Zuckerman, J.D.; Virk, M.S. Cytotoxicity Evaluation of Chlorhexidine Gluconate on Human Fibroblasts, Myoblasts, and Osteoblasts. J. Bone Jt. Infect. 2018, 3, 165–172. [Google Scholar] [CrossRef]
- Härtel, S.; Zorn-Kruppa, M.; Tykhonova, S.; Alajuuma, P.; Engelke, M.; Diehl, H.A. Staurosporine-induced Apoptosis in Human Cornea Epithelial Cells In Vitro. Cytom. Part A 2003, 55, 15–23. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Jasso, M.; Hernández-Martínez, D.; Avila-Acevedo, J.G.; del Carmen Benítez-Flores, C.; Gallegos-Hernández, I.A.; García-Bores, A.M.; Espinosa-González, A.M.; Villamar-Duque, T.E.; Castelan-Ramírez, I.; del Rosario González-Valle, M.; et al. Morphological Description of the Early Events during the Invasion of Acanthamoeba castellanii Trophozoites in a Murine Model of Skin Irradiated under UV-B Light. Pathogens 2020, 9, 794. [Google Scholar] [CrossRef] [PubMed]
- Omaña-Molina, M.; Hernandez-Martinez, D.; Sanchez-Rocha, R.; Cardenas-Lemus, U.; Salinas-Lara, C.; Mendez-Cruz, A.R.; Colin-Barenque, L.; Aley-Medina, P.; Espinosa-Villanueva, J.; Moreno-Fierros, L.; et al. In Vivo CNS Infection Model of Acanthamoeba Genotype T4: The Early Stages of Infection Lack Presence of Host Inflammatory Response and Are a Slow and Contact-Dependent Process. Parasitol. Res. 2017, 116, 725–733. [Google Scholar] [CrossRef] [PubMed]
- Culbertson, C.G.; Smith, J.W.; Cohen, H.K.; Minner, J.R. Experimental Infection of Mice and Monkeys by Acanthamoeba. Am. J. Pathol. 1959, 35, 185–197. [Google Scholar]
- Rodríguez-Expósito, R.L.; Sifaoui, I.; Reyes-Batlle, M.; Fuchs, F.; Scheid, P.L.; Piñero, J.E.; Sutak, R.; Lorenzo-Morales, J. Induction of Programmed Cell Death in Acanthamoeba culbertsoni by the Repurposed Compound Nitroxoline. Antioxidants 2023, 12, 2081. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene ID | Product | Fold Change Staurosporine/Control |
|---|---|---|
| ACA1_376810 | Universal stress domain containing protein | N.d. *. in control |
| ACA1_146430 | Catalase | 8.87 |
| ACA1_392960 | Universal stress domain containing protein | 3.44 |
| ACA1_387720 | Universal stress domain containing protein | 2.92 |
| ACA1_076370 | Profilin | 2.69 |
| ACA1_265580 | Manganese and iron superoxide dismutase | 2.67 |
| ACA1_031660 | Catalase | 2.59 |
| ACA1_062010 | Universal stress domain containing protein | 2.41 |
| ACA1_236670 | Universal stress protein (USP) family protein | 2.38 |
| ACA1_045080 | Thioredoxin, putative | 2.28 |
| ACA1_054650 | Glutathione transferase family protein | 2.03 |
| ACA1_095580 | Serine/threonine kinase | −2.16 |
| ACA1_288400 | Dual specificity protein kinase shkB, putative | −3.73 |
| ACA1_159080 | Papain family cysteine protease subfamily protein | N.d. in Staurosporine |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rodríguez-Expósito, R.L.; Sifaoui, I.; Salazar-Villatoro, L.; Bethencourt-Estrella, C.J.; Fernández, J.J.; Díaz-Marrero, A.R.; Sutak, R.; Omaña-Molina, M.; Piñero, J.E.; Lorenzo-Morales, J. Staurosporine as a Potential Treatment for Acanthamoeba Keratitis Using Mouse Cornea as an Ex Vivo Model. Mar. Drugs 2024, 22, 423. https://doi.org/10.3390/md22090423
Rodríguez-Expósito RL, Sifaoui I, Salazar-Villatoro L, Bethencourt-Estrella CJ, Fernández JJ, Díaz-Marrero AR, Sutak R, Omaña-Molina M, Piñero JE, Lorenzo-Morales J. Staurosporine as a Potential Treatment for Acanthamoeba Keratitis Using Mouse Cornea as an Ex Vivo Model. Marine Drugs. 2024; 22(9):423. https://doi.org/10.3390/md22090423
Chicago/Turabian StyleRodríguez-Expósito, Rubén L., Ines Sifaoui, Lizbeth Salazar-Villatoro, Carlos J. Bethencourt-Estrella, José J. Fernández, Ana R. Díaz-Marrero, Robert Sutak, Maritza Omaña-Molina, José E. Piñero, and Jacob Lorenzo-Morales. 2024. "Staurosporine as a Potential Treatment for Acanthamoeba Keratitis Using Mouse Cornea as an Ex Vivo Model" Marine Drugs 22, no. 9: 423. https://doi.org/10.3390/md22090423
APA StyleRodríguez-Expósito, R. L., Sifaoui, I., Salazar-Villatoro, L., Bethencourt-Estrella, C. J., Fernández, J. J., Díaz-Marrero, A. R., Sutak, R., Omaña-Molina, M., Piñero, J. E., & Lorenzo-Morales, J. (2024). Staurosporine as a Potential Treatment for Acanthamoeba Keratitis Using Mouse Cornea as an Ex Vivo Model. Marine Drugs, 22(9), 423. https://doi.org/10.3390/md22090423