

Recent Advances in Anti-Inflammatory Compounds from Marine Microorganisms
 , , ,
, , ,  and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
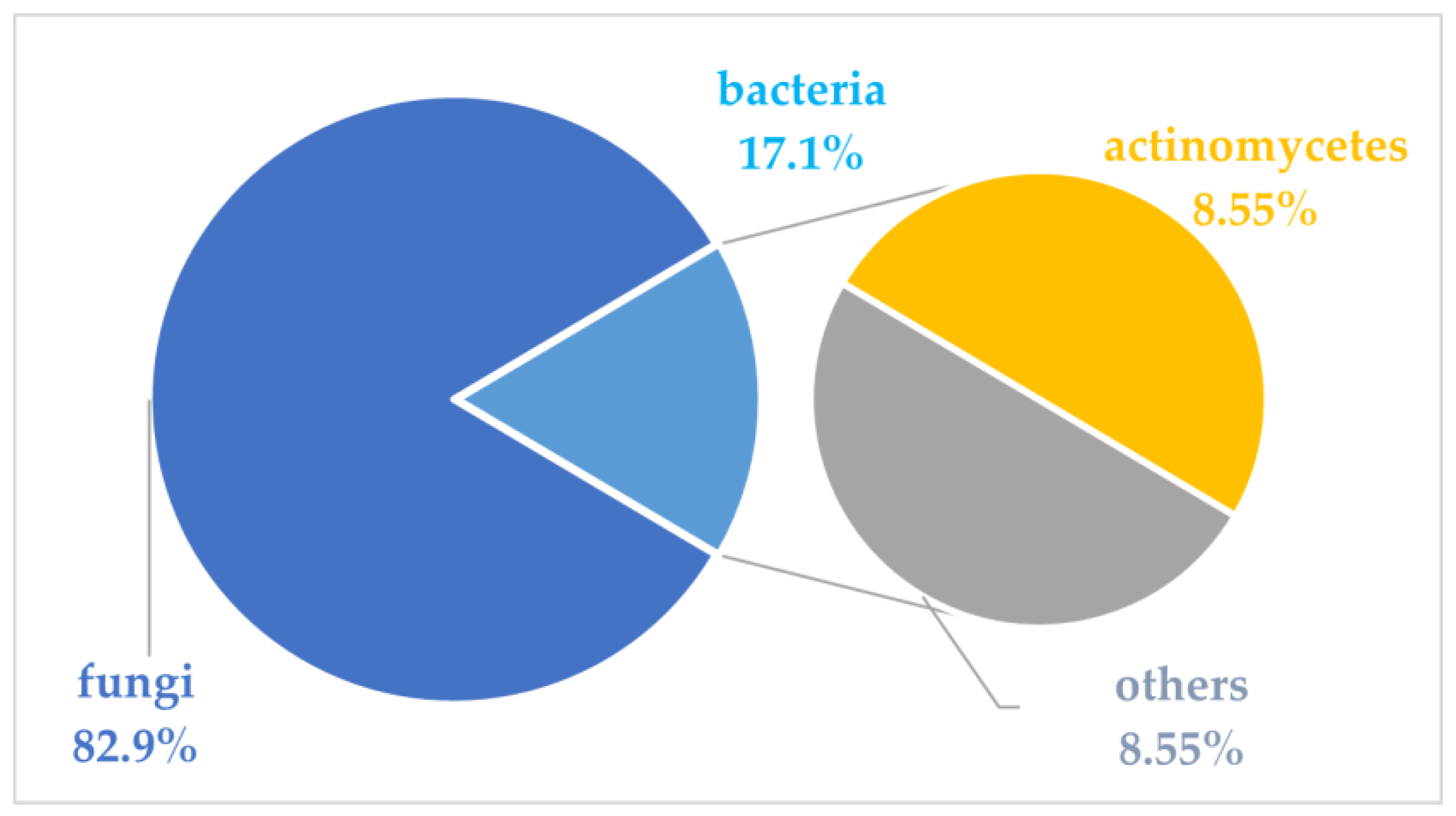
1. Introduction
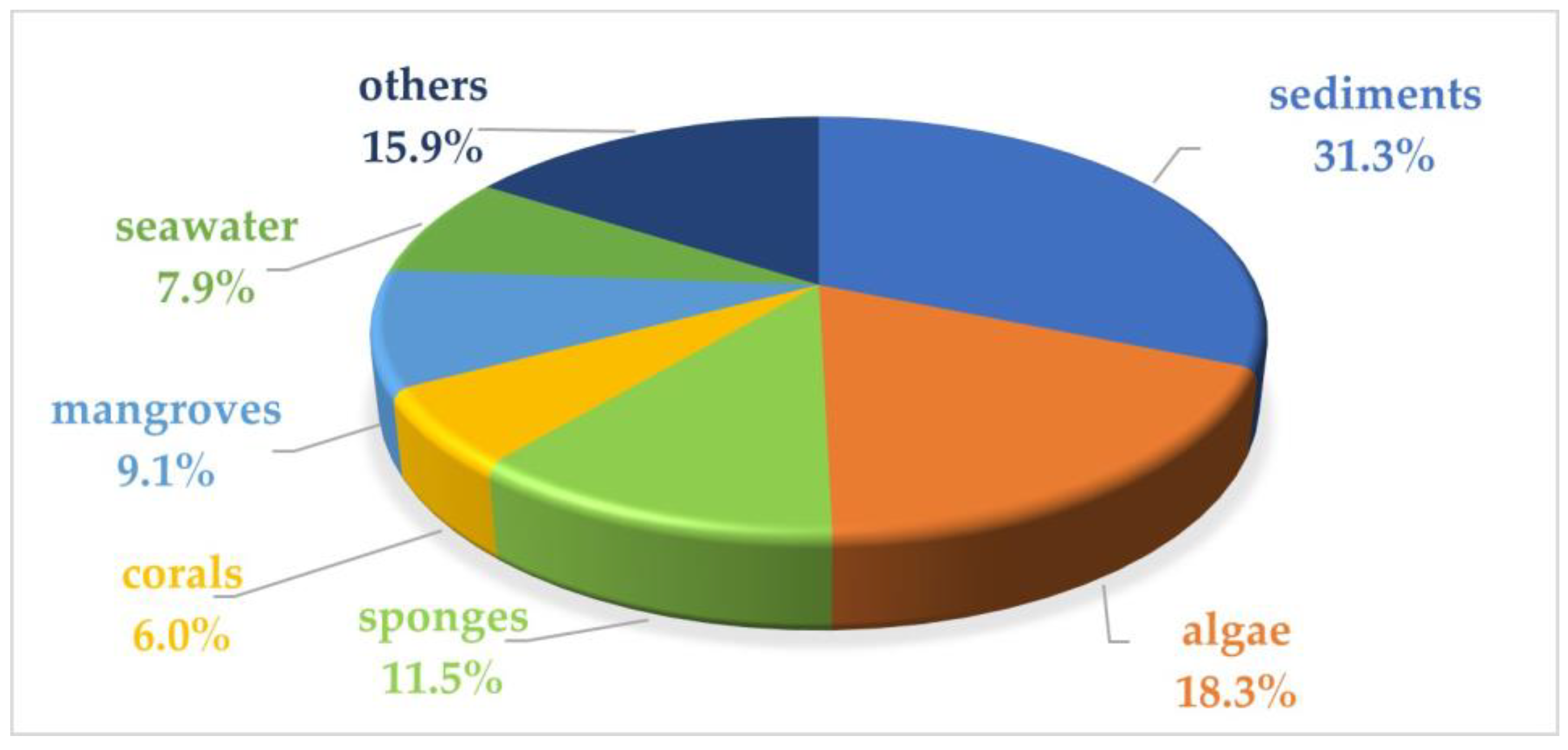
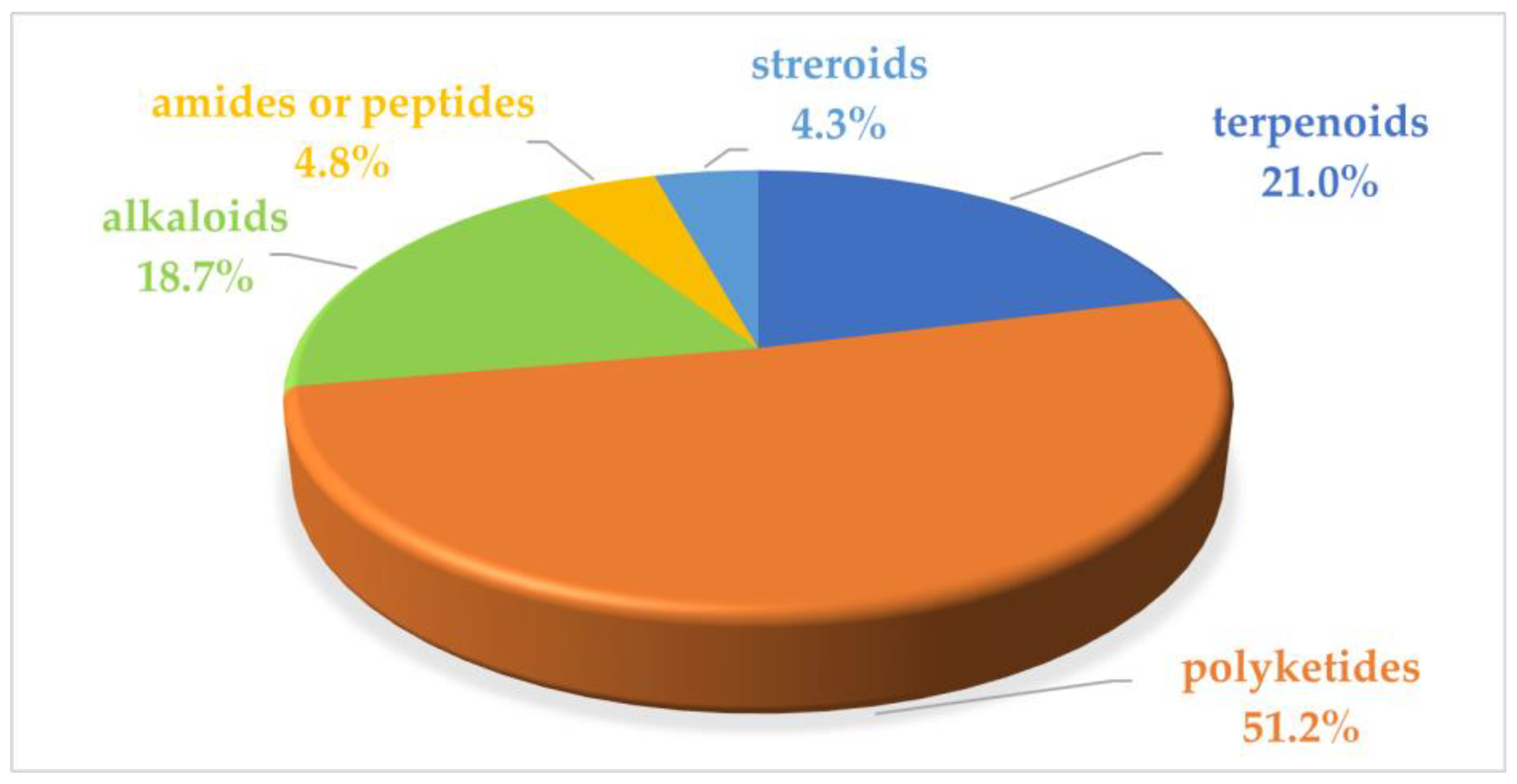
2. Marine Microbial Anti-Inflammatory Compounds
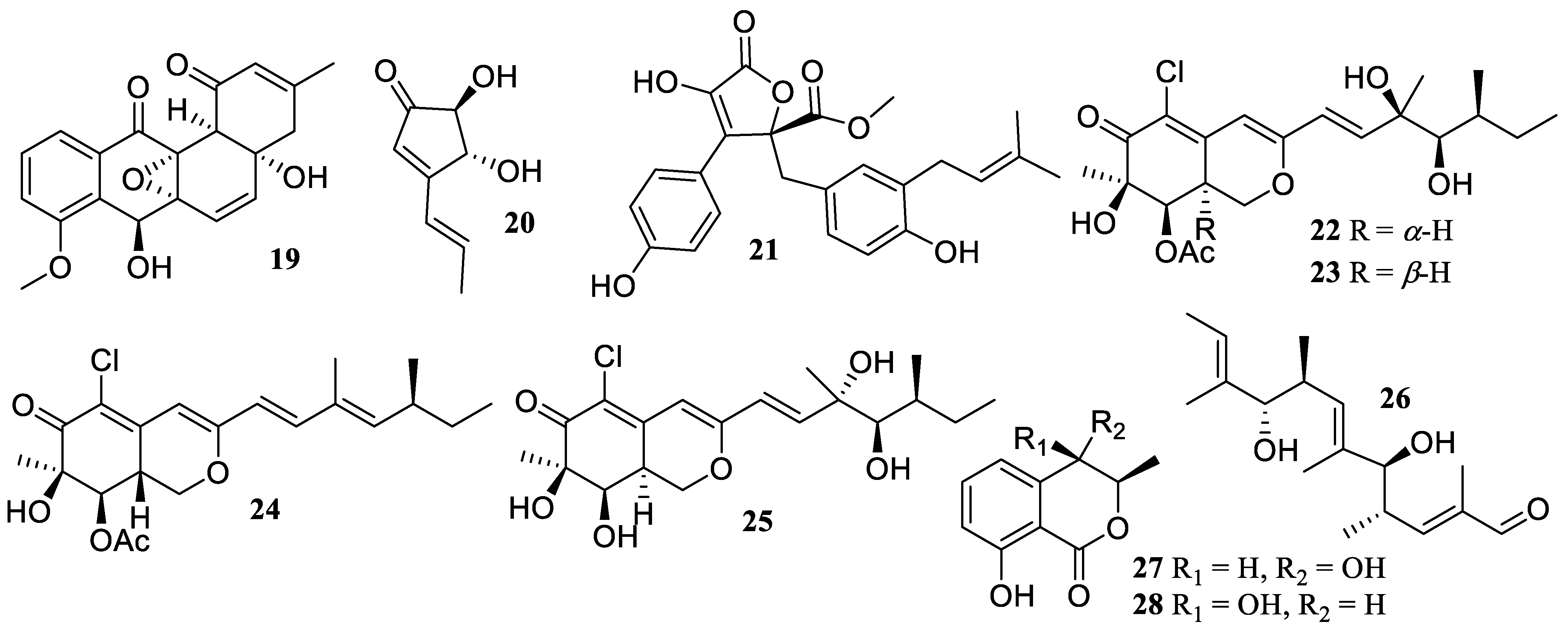
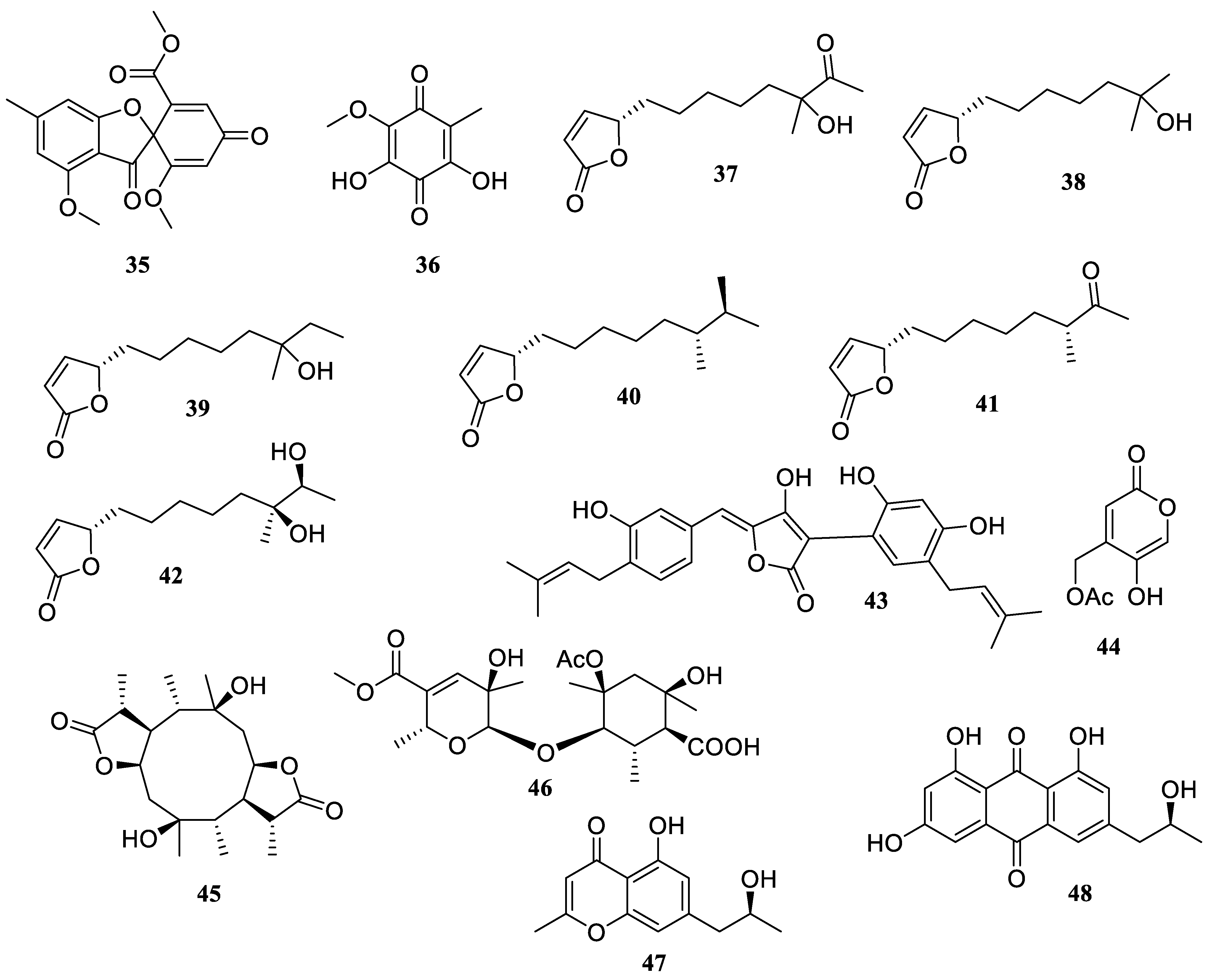
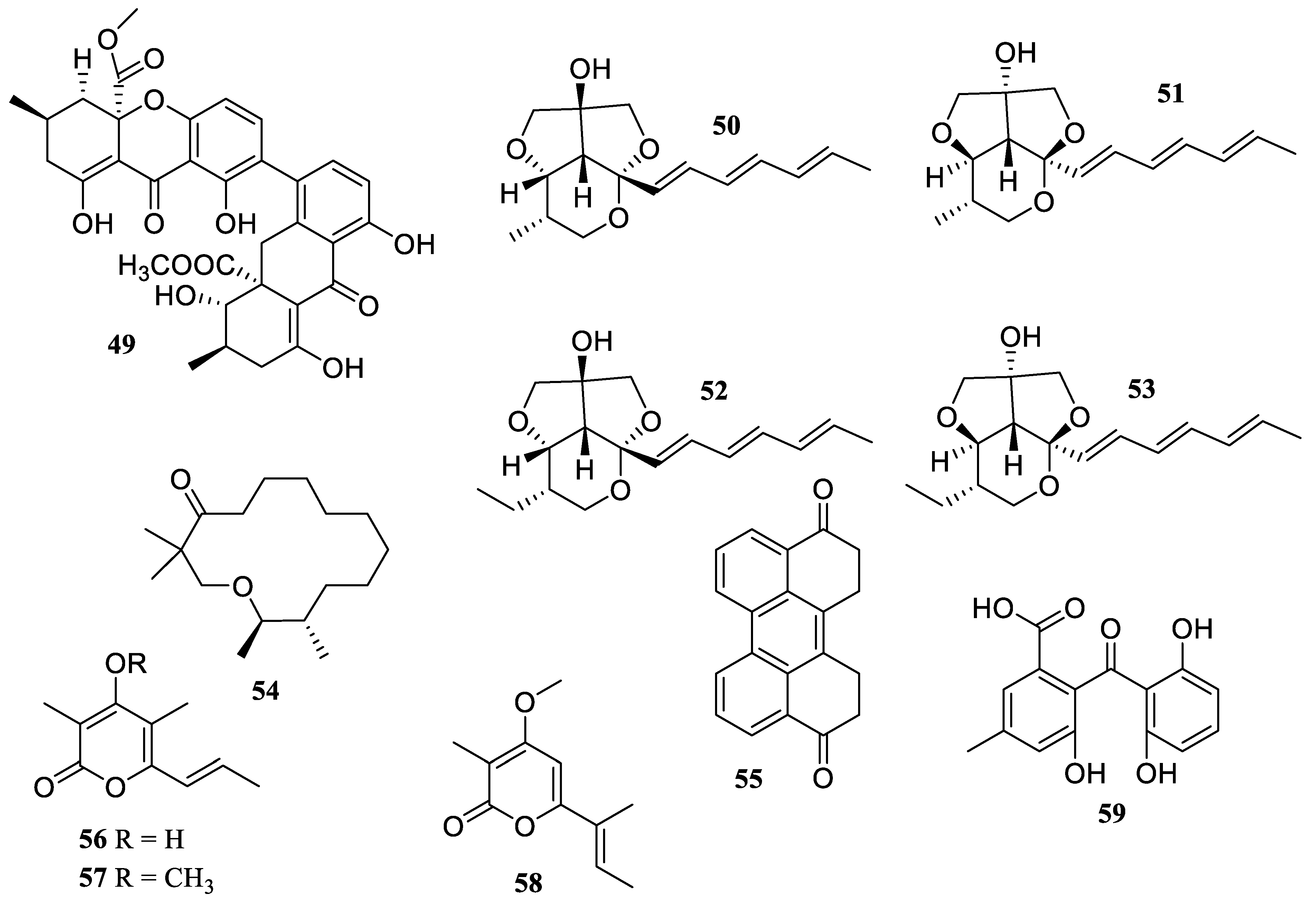
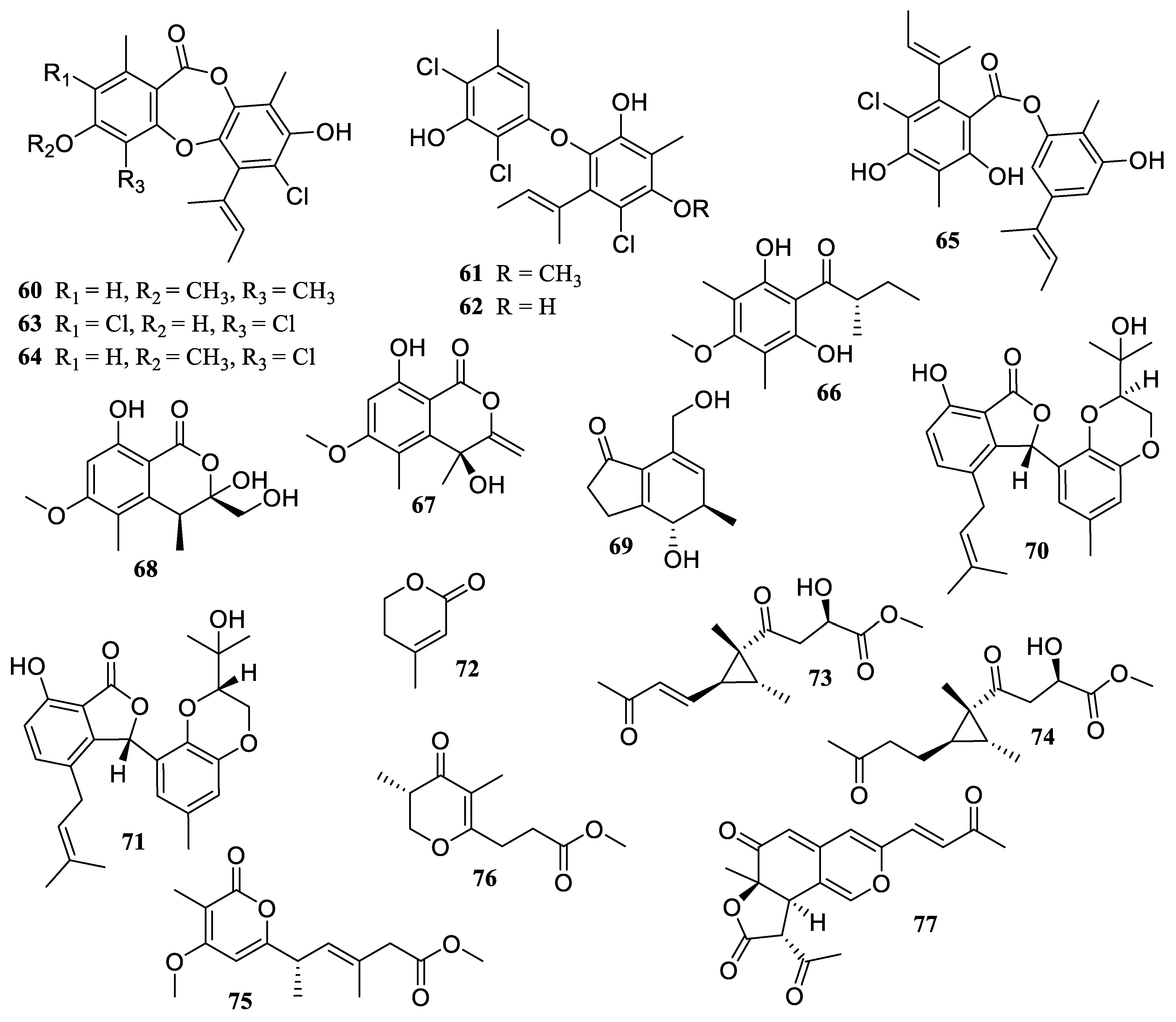
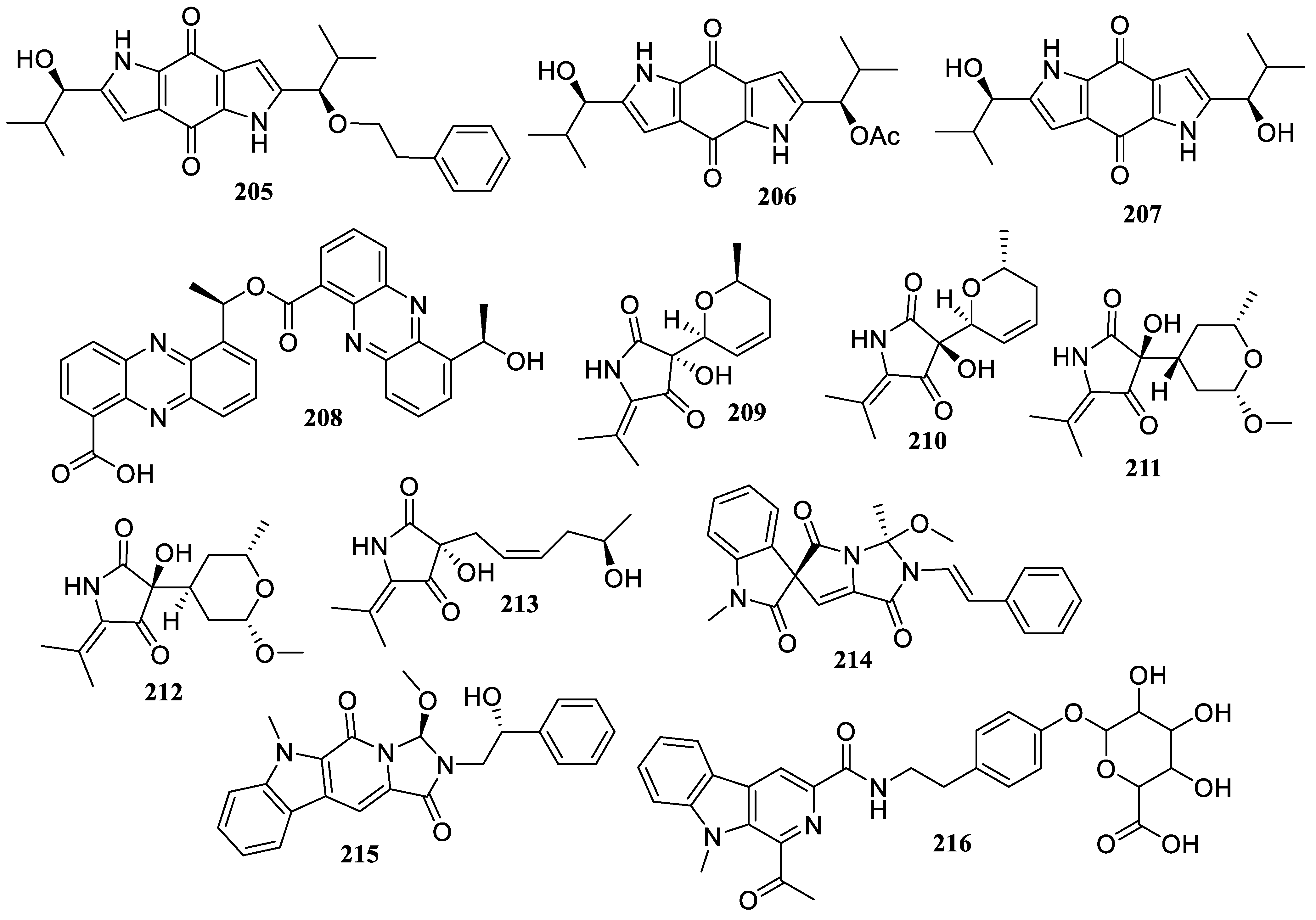
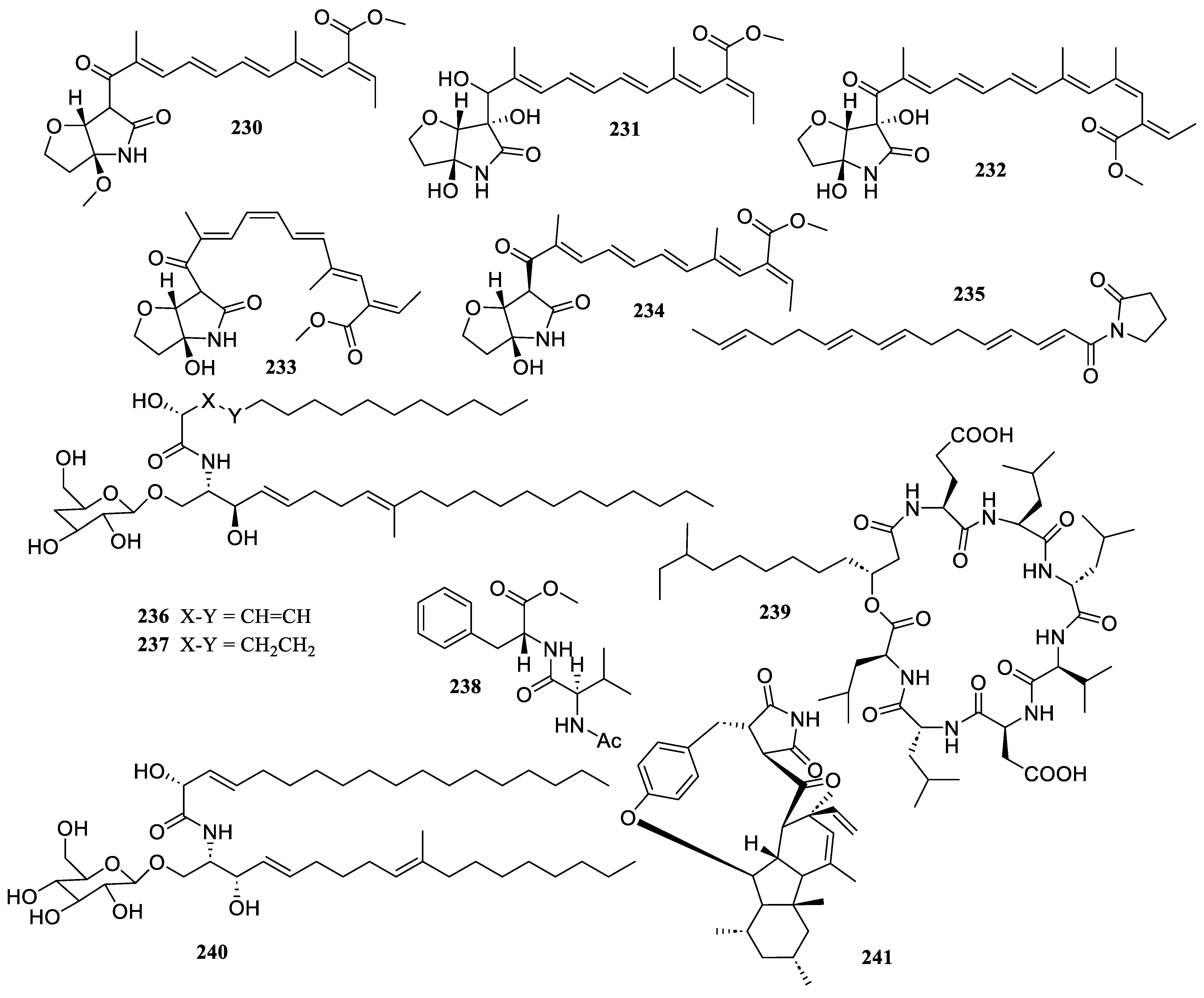
2.1. Polyketides
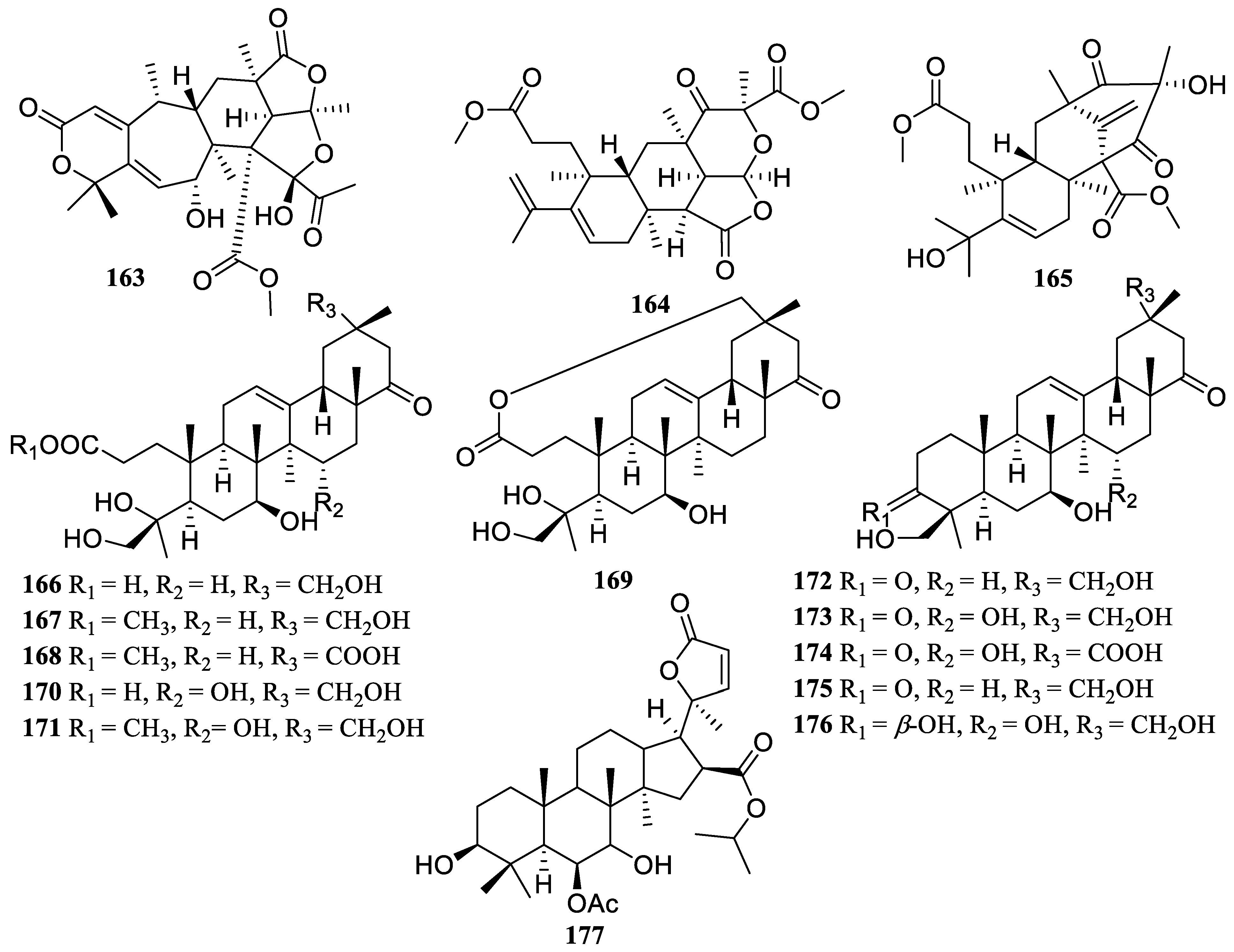
2.2. Terpenoids
2.2.1. Sesquiterpenes
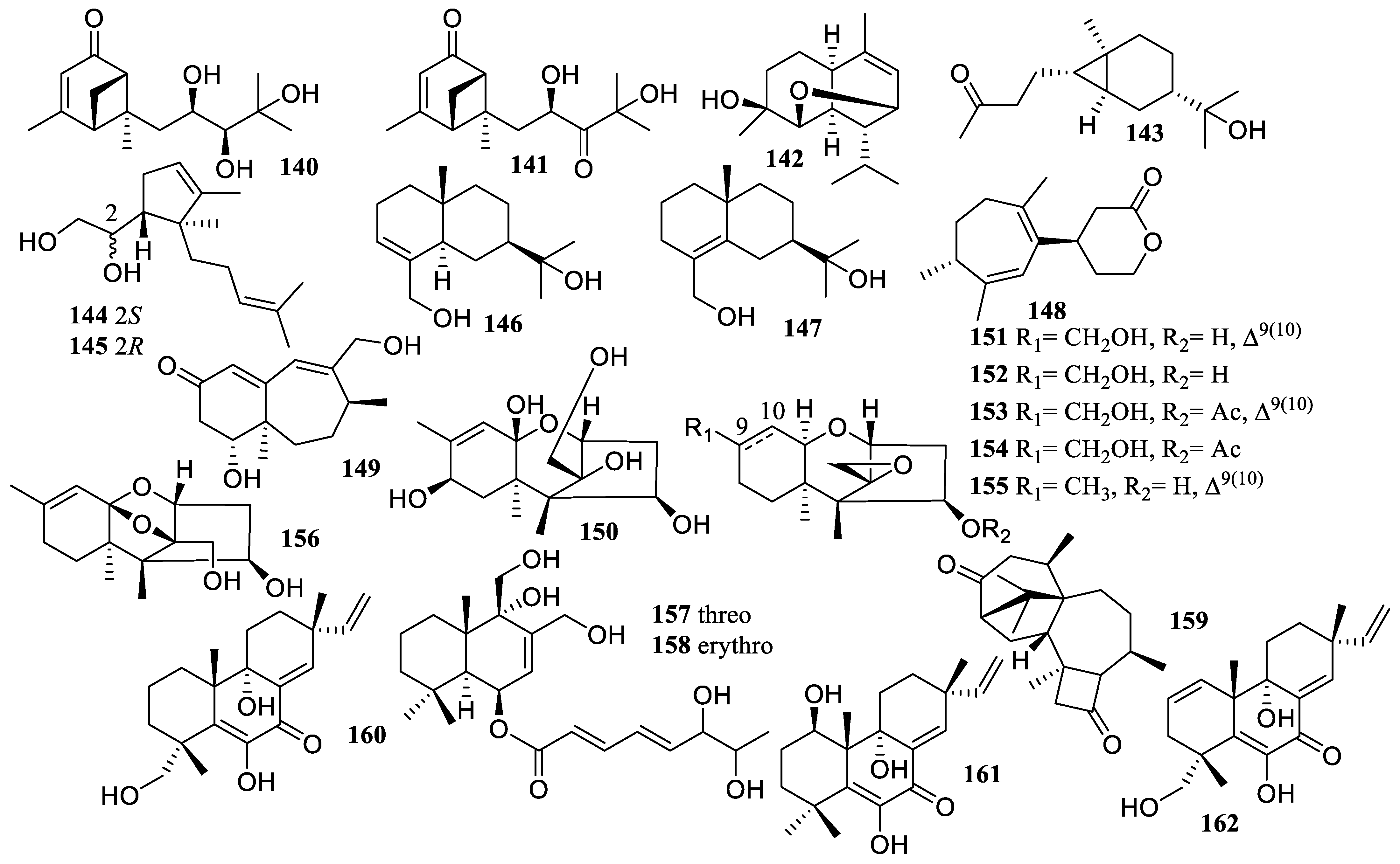
2.2.2. Diterpene
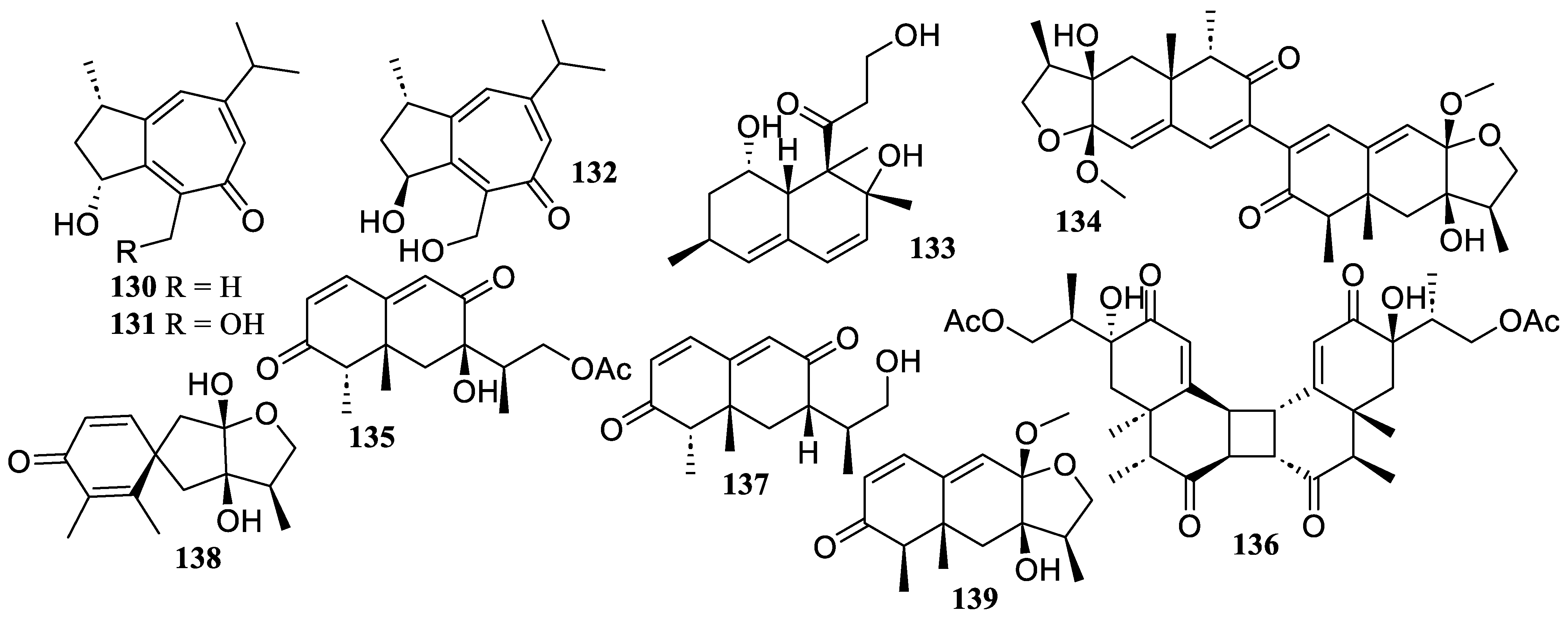
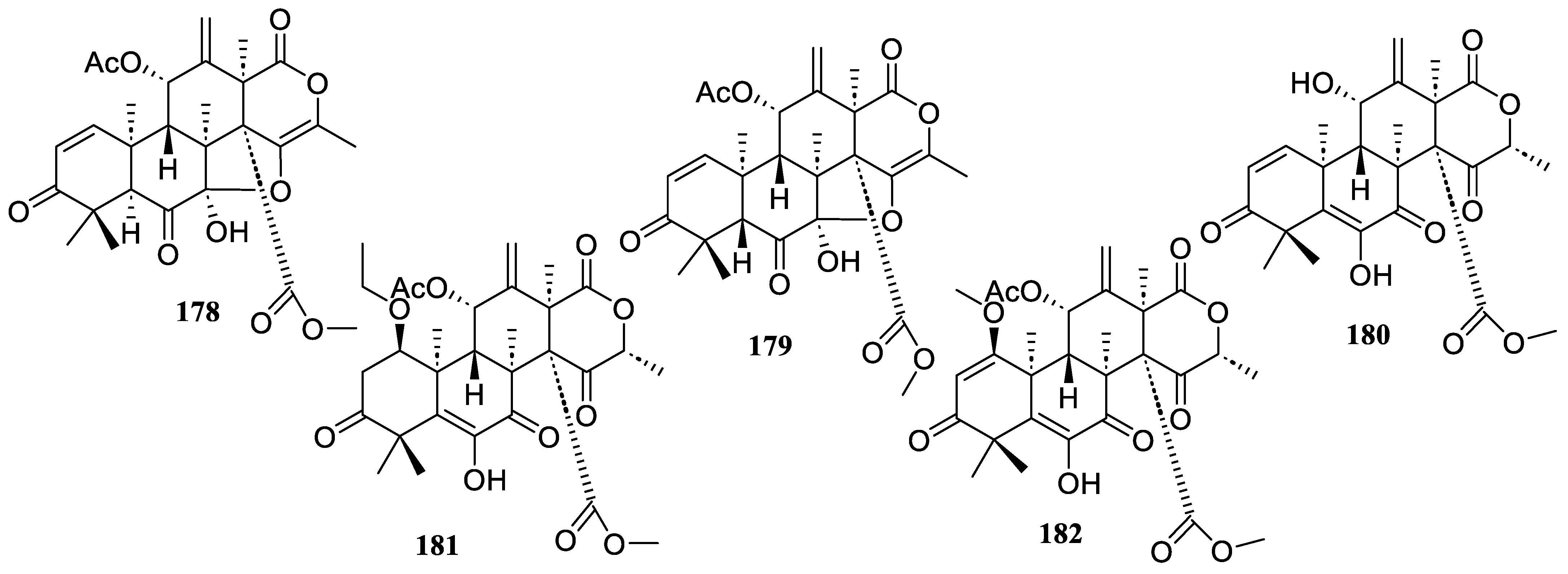
2.2.3. Triterpenoids
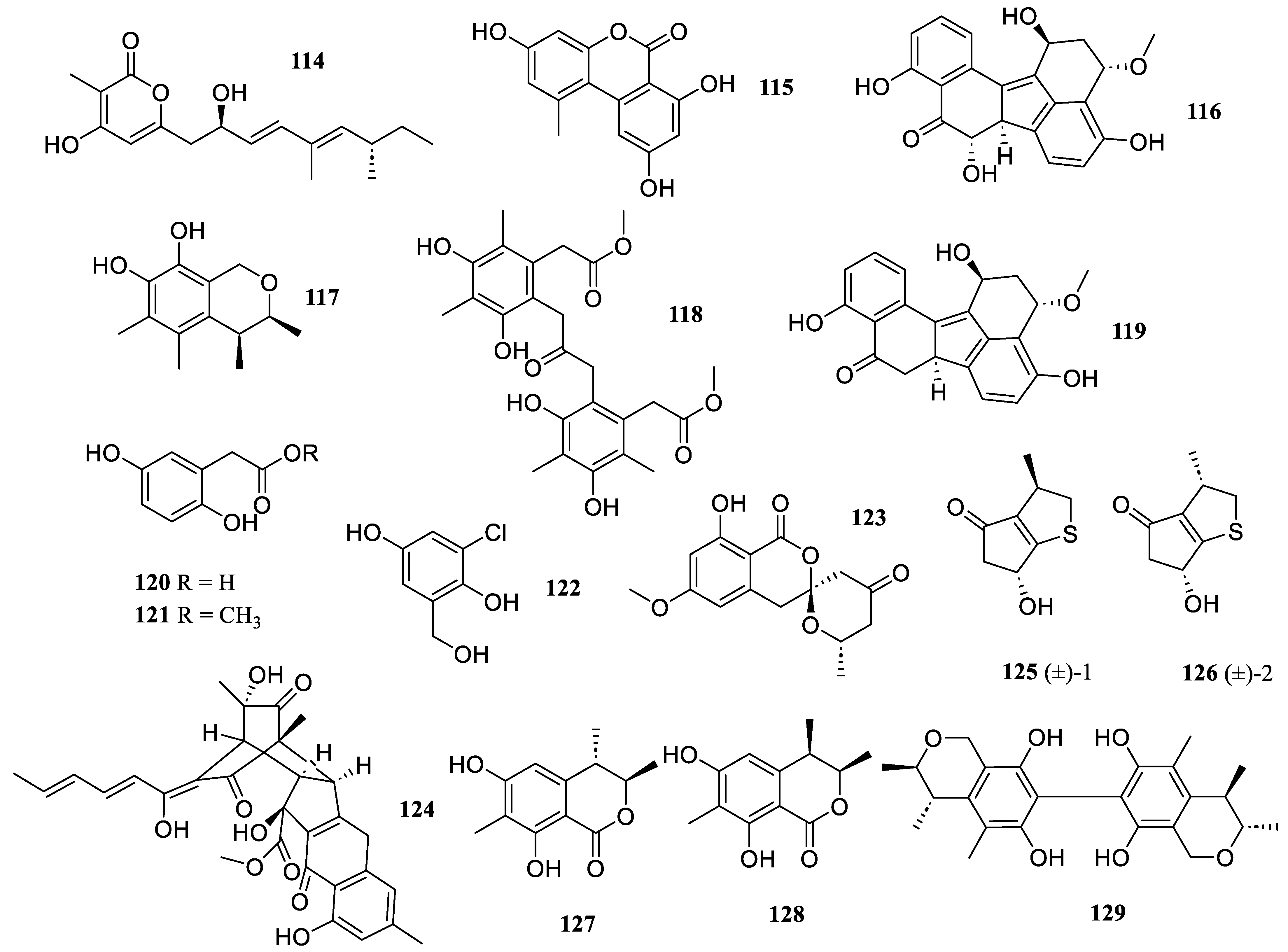
2.2.4. Meroterpenoids
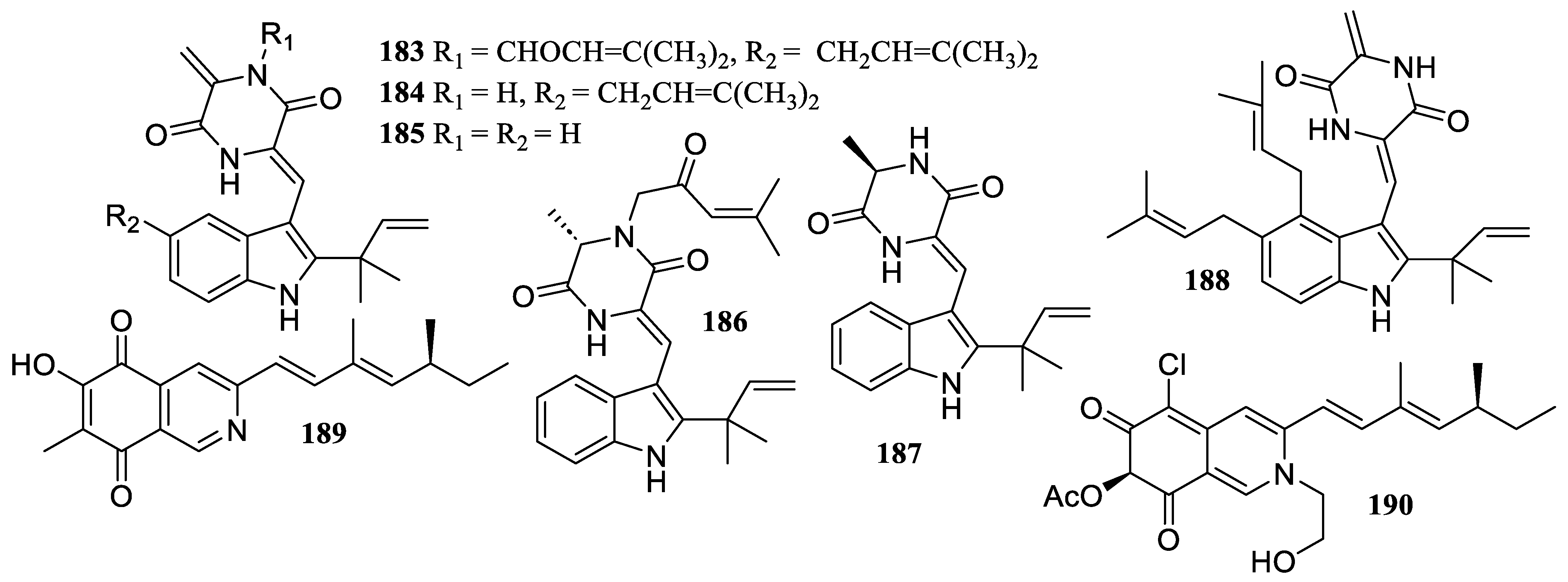
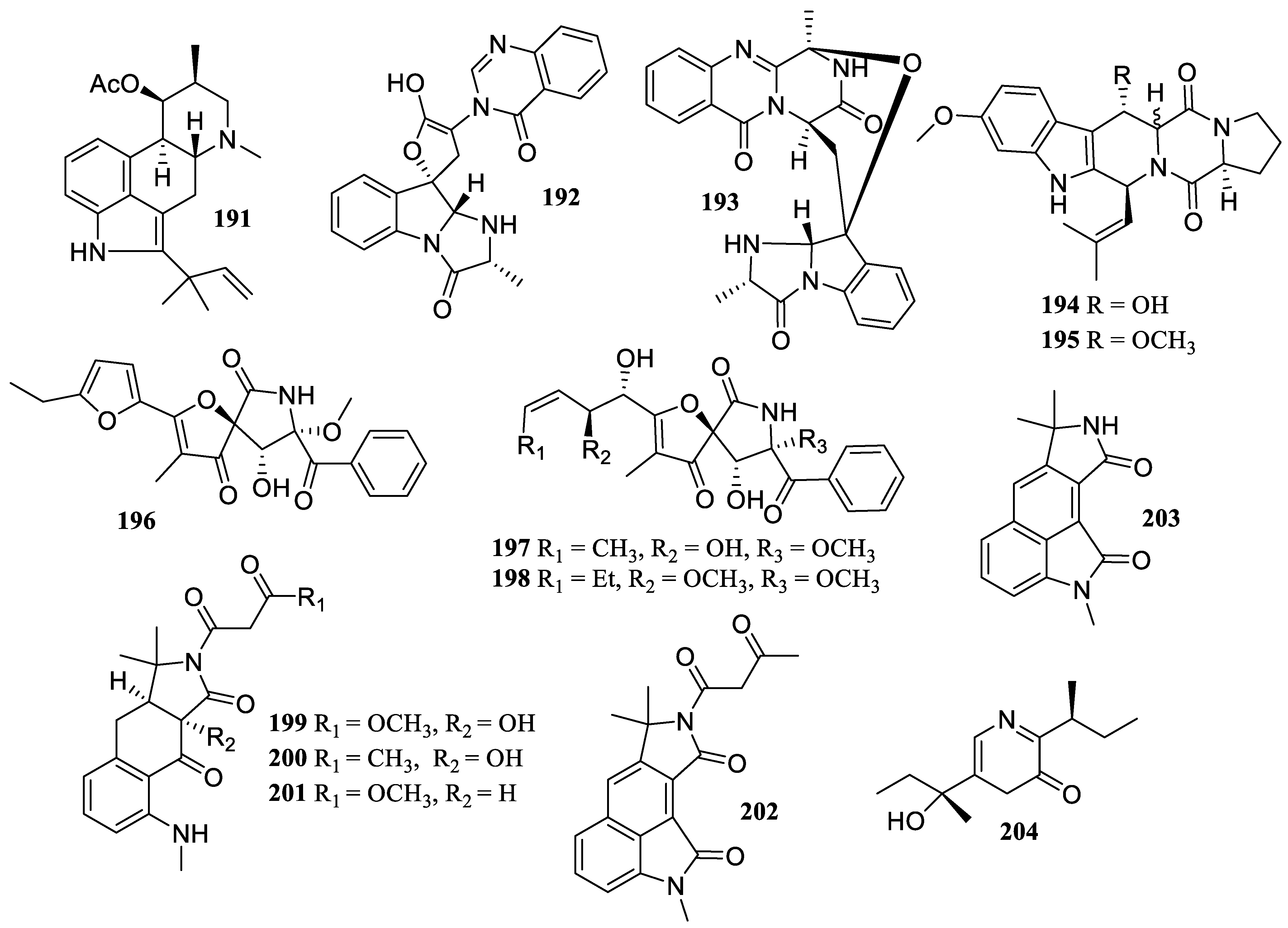
2.3. Alkaloids
2.4. Amides or Peptides
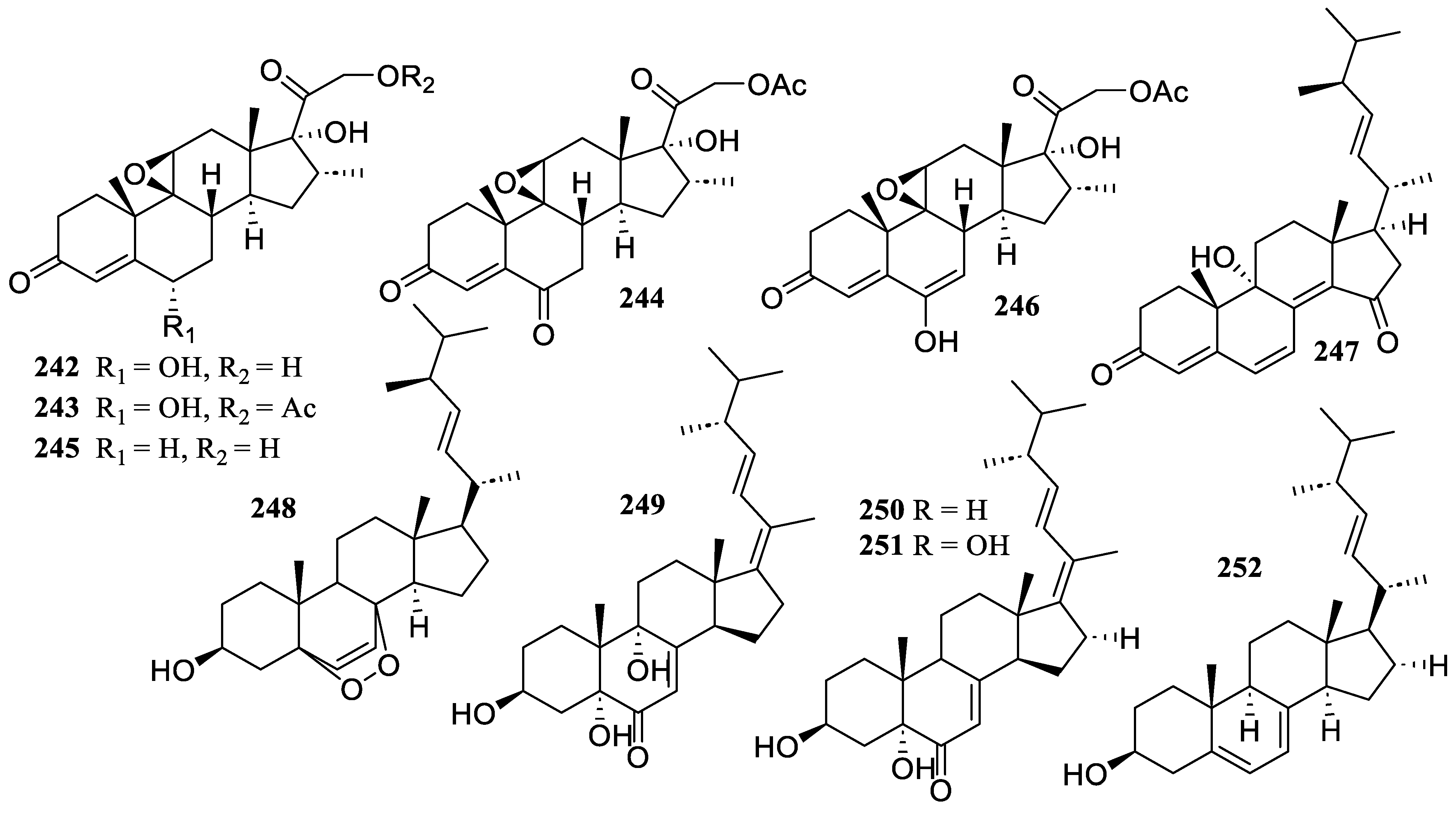
2.5. Steroids
3. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| IC50 | Half maximal inhibitory concentration |
| NF-κB | Nuclear factor kappa-B |
| LPS | Lipopolysaccharide |
| NO | Nitric oxide |
| IL-6 | Interleukine-6 |
| TNF-α | TNF-α |
References
- Almughrbi, A.H.; Crovella, S. Molecular analysis of inflammatory diseases. Exp. Dermatol. 2022, 31, 9–16. [Google Scholar] [CrossRef] [PubMed]
- Deng, L.Y.; He, S.S.; Guo, N.Q.; Tian, W.; Zhang, W.Z.; Luo, L.X. Molecular mechanisms of ferroptosis and relevance to inflammation. Inflamm. Res. 2022, 72, 281–299. [Google Scholar] [CrossRef] [PubMed]
- Lucido, M.J.; Bekhbat, M.; Goldsmith, D.R.; Treadway, M.T.; Haroon, E.; Felger, J.C.; Miller, A.H.; Dantzer, R. Aiding and abetting anhedonia: Impact of inflammation on the brain and pharmacological implications. Pharmacol. Rev. 2021, 73, 1084–1117. [Google Scholar] [CrossRef] [PubMed]
- Coll, R.C.; Schroder, K.; Pelegrín, P. NLRP3 and pyroptosis blockers for treating inflammatory diseases. Trends Pharmacol. Sci. 2022, 43, 653–668. [Google Scholar] [CrossRef]
- Fu, Y.; Xiang, Y.; Li, H.L.; Chen, A.Q.; Dong, Z. Inflammation in kidney repair: Mechanism and therapeutic potential. Pharmacol. Ther. 2022, 237, 108240. [Google Scholar] [CrossRef]
- Squillace, S.; Salvemini, D. Toll-like receptor-mediated neuroinflammation: Relevance for cognitive dysfunctions. Trends Pharmacol. Sci. 2022, 43, 726–739. [Google Scholar] [CrossRef]
- Nicolaes, G.A.F.; Soehnlein, O. Targeting extranuclear histones to alleviate acute and chronic inflammation. Trends Pharmacol. Sci. 2024, 45, 651–662. [Google Scholar] [CrossRef]
- Cryer, B. Nonsteroidal anti-inflammatory drug gastrointestinal toxicity. Curr. Opin. Gastroen. 2001, 17, 503–512. [Google Scholar] [CrossRef]
- Gao, F.; Dai, Z.Q.; Zhang, T.; Gu, Y.H.; Cai, D.S.; Lu, M.J.; Zhang, Z.J.; Zeng, Q.; Shang, B.X.; Xu, B.; et al. Synthesis and biological evaluation of novel sinomenine derivatives as anti-inflammatory and analgesic agent. RSC Adv. 2022, 12, 30001–30007. [Google Scholar] [CrossRef]
- Barzkar, N.; Sukhikh, S.; Babich, O. Study of marine microorganism metabolites: New resources for bioactive natural products. Front. Microbiol. 2024, 14, 1285902. [Google Scholar] [CrossRef]
- Wang, Y.N.; Meng, L.H.; Wang, B.G. Progress in research on bioactive secondary metabolites from deep-sea derived microorganisms. Mar. Drugs 2020, 18, 614. [Google Scholar] [CrossRef] [PubMed]
- Li, C.Q.; Ma, Q.Y.; Gao, X.Z.; Wang, X.; Zhang, B.L. Research progress in anti-inflammatory bioactive substances derived from marine microorganisms, sponges, algae, and corals. Mar. Drugs 2021, 19, 572. [Google Scholar] [CrossRef] [PubMed]
- Ding, Y.; An, F.; Zhu, X.; Yu, H.Y.; Hao, L.L.; Lu, Y.H. Curdepsidones B-G, six depsidones with anti-inflammatory activities from the marine-derived fungus Curvularia sp. IFB-Z10. Mar. Drugs 2019, 17, 266. [Google Scholar] [CrossRef] [PubMed]
- Li, H.Y.; Huang, H.M.; Hou, L.K.; Ju, J.H.; Li, W.L. Discovery of antimycin-type depsipeptides from a wbl gene mutant strain of deepsea-derived Streptomyces somaliensis SCSIO ZH66 and their effects on pro-inflammatory cytokine production. Front. Microbiol. 2017, 8, 678. [Google Scholar] [CrossRef]
- Carroll, A.R.; Copp, B.R.; Davis, R.A.; Keyzers, R.A.; Prinsep, M.R. Marine natural products. Nat. Prod. Rep. 2020, 37, 175–223. [Google Scholar] [CrossRef]
- Carroll, A.R.; Copp, B.R.; Davis, R.A.; Keyzers, R.A.; Prinsep, M.R. Marine natural products. Nat. Prod. Rep. 2021, 38, 362–413. [Google Scholar] [CrossRef]
- Carroll, A.R.; Copp, B.R.; Davis, R.A.; Keyzers, R.A.; Prinsep, M.R. Marine natural products. Nat. Prod. Rep. 2022, 39, 1122–1171. [Google Scholar] [CrossRef]
- Carroll, A.R.; Copp, B.R.; Davis, R.A.; Keyzers, R.A.; Prinsep, M.R. Marine natural products. Nat. Prod. Rep. 2023, 40, 275–325. [Google Scholar] [CrossRef]
- Carroll, A.R.; Copp, B.R.; Grkovic, T.; Keyzers, R.A.; Prinsep, M.R. Marine natural products. Nat. Prod. Rep. 2024, 41, 162–207. [Google Scholar] [CrossRef]
- Alcaraz, M.J.; Paya, M. Marine sponge metabolites for the control of inflammatory diseases. Curr. Opin. Investig. Drugs 2006, 7, 974–979. [Google Scholar]
- Cheung, R.C.F.; Ng, T.B.; Wong, J.H.; Chen, Y.; Chan, W.Y. Marine natural products with anti-inflammatory activity. Appl. Microbiol. Biotechnol. 2015, 100, 1645–1666. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.Z.; Yi, M.Q.; Ding, L.J.; He, S. A review of anti-inflammatory compounds from marine fungi, 2000–2018. Mar. Drugs 2019, 17, 636. [Google Scholar] [CrossRef] [PubMed]
- Souza, C.R.M.; Bezerra, W.P.; Souto, J.T. Marine alkaloids with anti-inflammatory activity: Current knowledge and future perspectives. Mar. Drugs 2020, 18, 147. [Google Scholar] [CrossRef] [PubMed]
- Chu, Y.C.; Chang, C.H.; Liao, H.R.; Cheng, M.J.; Wu, M.D.; Fu, S.L.; Chen, J.J. Rare chromone derivatives from the marine-derived Penicillium citrinum with anti-cancer and anti-inflammatory activities. Mar. Drugs 2021, 19, 25. [Google Scholar] [CrossRef]
- Chu, Y.C.; Chang, C.H.; Liao, H.R.; Fu, S.L.; Chen, J.J. Anti-cancer and anti-inflammatory activities of three new chromone derivatives from the marine-derived Penicillium citrinum. Mar. Drugs 2021, 19, 408. [Google Scholar] [CrossRef] [PubMed]
- Ding, W.J.; Wang, F.F.; Li, Q.W.; Xue, Y.X.; Dong, Z.T.; Tian, D.M.; Chen, M.; Zhang, Y.W.; Hong, K.; Tang, J.S. Isolation and characterization of anti-inflammatory sorbicillinoids from the mangrove-derived fungus Penicillium sp. DM 815. Chem. Biodivers. 2021, 18, e2100229. [Google Scholar]
- Gou, X.S.; Tian, D.M.; Wei, J.H.; Ma, Y.H.; Zhang, Y.X.; Chen, M.; Ding, W.J.; Wu, B.; Tang, J.S. New drimane sesquiterpenes and polyketides from marine-derived fungus Penicillium sp. TW58-16 and their anti-inflammatory and α-glucosidase inhibitory effects. Mar. Drugs 2021, 19, 416. [Google Scholar] [CrossRef]
- Guo, H.; Wu, Q.L.; Chen, D.G.; Jiang, M.H.; Chen, B.; Lu, Y.J.; Li, J.; Liu, L.; Chen, S.H. Absolute configuration of polypropionate derivatives: Decempyrones A-J and their MptpA inhibition and anti-inflammatory activities. Bioorg. Chem. 2021, 115, 105156. [Google Scholar] [CrossRef]
- Lei, H.; Bi, X.X.; Lin, X.P.; She, J.L.; Luo, X.W.; Niu, H.; Zhang, D.; Yang, B. Heterocornols from the sponge-derived fungus Pestalotiopsis heterocornis with anti-inflammatory activity. Mar. Drugs 2021, 19, 585. [Google Scholar] [CrossRef]
- Luo, G.Y.; Li, Z.; Wu, Q.L.; Chen, S.H.; Li, J.; Liu, L. Fusarins G-L with Inhibition of NO in RAW 264.7 from marine-derived fungus Fusarium solani 7227. Mar. Drugs 2021, 19, 305. [Google Scholar] [CrossRef]
- Wang, S.; Zeng, Y.B.; Yin, J.J.; Chang, W.J.; Zhao, X.L.; Mao, Y. Two new azaphilones from the marine-derived fungus Penicillium sclerotiorum E23Y-1A. Phytochem. Lett. 2021, 47, 76–80. [Google Scholar] [CrossRef]
- Shen, Q.Y.; Dai, G.Z.; Li, A.Y.; Liu, Y.; Zhong, G.N.; Li, X.J.; Ren, X.M.; Sui, H.Y.; Fu, J.; Jiao, N.Z.; et al. Genome-guided discovery of highly oxygenated aromatic polyketides, saccharothrixins D-M, from the rare marine actinomycete Saccharothrix sp. D09. J. Nat. Prod. 2021, 84, 2875–2884. [Google Scholar] [CrossRef] [PubMed]
- Tilvi, S.; Parvatkar, R.; Singh, K.S.; Devi, P. Chemical investigation of marine-derived fungus Aspergillus flavipes for potential anti-inflammatory agents. Chem. Biodivers. 2021, 18, e2000956. [Google Scholar] [CrossRef] [PubMed]
- Uras, I.S.; Ebada, S.S.; Korinek, M.; Albohy, A.; Abdulrazik, B.S.; Wang, Y.H.; Chen, B.H.; Horng, J.T.; Lin, W.; Hwang, T.L.; et al. Anti-inflammatory, antiallergic, and COVID-19 main protease (Mpro) inhibitory activities of butenolides from a marine-derived fungus Aspergillus terreus. Molecules 2021, 26, 3354. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.C.; Ke, T.Y.; Ko, Y.C.; Lin, J.J.; Chang, J.S.; Cheng, Y.B. Anti-inflammatory azaphilones from the edible alga-derived fungus Penicillium sclerotiorum. Mar. Drugs 2021, 19, 529. [Google Scholar] [CrossRef]
- Yang, W.C.; Yuan, J.; Tan, Q.; Chen, Y.; Zhu, Y.J.; Jiang, H.M.; Zou, G.; Zang, Z.M.; Wang, B.; She, Z.G. Peniazaphilones A-I, produced by co-culturing of mangrove endophytic fungi, Penicillium sclerotiorum THSH-4 and Penicillium sclerotiorum ZJHJJ-18. Chin. J. Chem. 2021, 39, 3404–3412. [Google Scholar] [CrossRef]
- Chen, C.M.; Ren, X.; Tao, H.M.; Cai, W.T.; Chen, Y.C.; Luo, X.W.; Guo, P.; Liu, Y.H. Anti-inflammatory polyketides from an alga-derived fungus Aspergillus ochraceopetaliformis SCSIO 41020. Mar. Drugs 2022, 20, 295. [Google Scholar] [CrossRef]
- Chen, S.H.; Guo, H.; Jiang, M.H.; Wu, Q.L.; Li, J.; Shen, H.J.; Liu, L. Mono- and dimeric xanthones with anti-glioma and anti-inflammatory activities from the ascidian-derived fungus Diaporthe sp. SYSU-MS 4722. Mar. Drugs 2022, 20, 51. [Google Scholar] [CrossRef]
- Cong, M.J.; Zhang, Y.; Feng, X.Y.; Pang, X.Y.; Liu, Y.H.; Zhang, X.Y.; Yang, Z.Y.; Wang, J.F. Anti-inflammatory alkaloids from the cold-seep-derived fungus Talaromyces helicus SCSIO 41311. 3 Biotech 2022, 12, 161. [Google Scholar] [CrossRef]
- Gao, M.; Lee, S.B.; Lee, J.E.; Kim, G.J.; Moon, J.; Nam, J.W.; Bae, J.S.; Chin, J.; Jeon, Y.H.; Choi, H. Anti-inflammatory butenolides from a marine-derived Streptomyces sp. 13G036. Appl. Sci. 2022, 12, 4510. [Google Scholar] [CrossRef]
- Hsiao, G.; Chi, W.C.; Chang, C.H.; Chiang, Y.R.; Fu, Y.J.; Lee, T.H. Bioactive pulvinones from a marine algicolous fungus Aspergillus terreus NTU243. Phytochemistry 2022, 200, 113229. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.M.; Li, Z.Z.; Zhang, Y.T.; Chen, C.M.; Chen, W.H.; Gao, C.H.; Liu, Y.H.; Tan, Y.H.; Luo, X.W. A new α-cyclopiazonic acid alkaloid identified from the Weizhou Island coral-derived fungus Aspergillus flavus GXIMD 02503. J. Ocean Univ. China 2022, 21, 1307–1312. [Google Scholar] [CrossRef]
- Jiang, P.; Luo, J.F.; Jiang, Y.; Zhang, L.P.; Jiang, L.Y.; Teng, B.R.; Niu, H.; Zhang, D.; Lei, H. Anti-inflammatory polyketide derivatives from the sponge-derived fungus Pestalotiopsis sp. SWMU-WZ04-2. Mar. Drugs 2022, 20, 711. [Google Scholar] [CrossRef]
- Kim, D.C.; Tran Hong, Q.; Nguyen Thuy, T.; Kim, K.W.; Kim, Y.C.; Thanh, N.T.; Cuong, N.X.; Nam, H.N.; Oh, H. Anti-neuroinflammatory effect of oxaline, isorhodoptilometrin, and 5-hydroxy-7-(2′-hydroxypropyl)-2-methyl-chromone obtained from the marine fungal strain Penicillium oxalicum CLC-MF05. Arch. Pharm. Res. 2022, 45, 90–104. [Google Scholar] [CrossRef]
- Long, J.Y.; Pang, X.Y.; Lin, X.P.; Liao, S.Y.; Zhou, X.F.; Wang, J.F.; Yang, B.; Liu, Y.H. Asperbenzophenone A and versicolamide C, new fungal metabolites from the soft coral derived Aspergillus sp. SCSIO 41036. Chem. Biodivers. 2022, 19, e202100925. [Google Scholar] [CrossRef]
- Shin, H.J.; Heo, C.S.; Anh, C.V.; Yoon, Y.D.; Kang, J.S. Streptoglycerides E-H, unsaturated polyketides from the marine-derived Bacterium Streptomyces specialis and their anti-inflammatory activity. Mar. Drugs 2022, 20, 44. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.S.; Yin, Z.H.; Wang, S.Y.; Yuan, Y.L.; Chen, Y.; Kang, W.Y. Diversified polyketides with anti-inflammatory activities from mangrove endophytic fungus Daldinia eschscholtzii KBJYZ-1. Front. Microbiol. 2022, 13, 900227. [Google Scholar] [CrossRef]
- Yuan, S.W.; Chen, L.T.; Wu, Q.L.; Jiang, M.H.; Guo, H.; Hu, Z.B.; Chen, S.H.; Liu, L.; Gao, Z.Z. Genome mining of α-pyrone natural products from ascidian-derived fungus Amphichordafelina SYSU-MS 7908. Mar. Drugs 2022, 20, 294. [Google Scholar] [CrossRef]
- Zhai, G.F.; Chen, S.H.; Shen, H.J.; Guo, H.; Jiang, M.H.; Liu, L. Bioactive monoterpenes and polyketides from the ascidian-derived fungus Diaporthe sp. SYSU-MS4722. Mar. Drugs 2022, 20, 553. [Google Scholar] [CrossRef]
- Zhang, Y.T.; Li, Z.Z.; Huang, B.Y.; Liu, K.; Peng, S.; Liu, X.M.; Gao, C.H.; Liu, Y.H.; Tan, Y.H.; Luo, X.W. Anti-osteoclastogenic and antibacterial effects of chlorinated polyketides from the Beibu Gulf coral-derived fungus Aspergillus unguis GXIMD 02505. Mar. Drugs 2022, 20, 178. [Google Scholar] [CrossRef]
- Zhang, Y.H.; Du, H.F.; Gao, W.B.; Li, W.; Cao, F.; Wang, C.Y. Anti-inflammatory polyketides from the marine-derived fungus Eutypella scoparia. Mar. Drugs 2022, 20, 486. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.S.; Nagahawatta, D.P.; Jeon, Y.J.; Lee, M.A.; Heo, C.S.; Park, S.J.; Shin, H.J. Streptinone, a new indanone derivative from a marine-derived Streptomyces massiliensis, inhibits particulate matter-induced inflammation. Mar. Drugs 2023, 21, 640. [Google Scholar] [CrossRef] [PubMed]
- Tan, Z.L.; Chen, Y.C.; Zhang, J.P.; Liu, H.X.; Zhang, W.M.; Yan, H.J. A new secondary metabolite from the marine-derived fungus Phomopsis lithocarpus FS 508. J. Asian Nat. Prod. Res. 2023, 26, 534–540. [Google Scholar] [CrossRef]
- Wu, Z.G.; Guo, H.; Wu, Q.L.; Jiang, M.H.; Chen, J.J.; Chen, B.; Li, H.X.; Liu, L.; Chen, S.H. Absolute configuration of cyclopropanes and the structural revision of pyrones from marine-derived fungus Stagonospora sp. SYSU-MS 7888. Bioorg. Chem. 2023, 136, 106542. [Google Scholar] [CrossRef]
- Zeng, Y.B.; Wang, Z.; Chang, W.J.; Zhao, W.B.; Wang, H.; Chen, H.Q.; Dai, H.F.; Lv, F. New azaphilones from the marine-derived fungus Penicillium sclerotiorum E23Y-1A with their anti-inflammatory and antitumor activities. Mar. Drugs 2023, 21, 75. [Google Scholar] [CrossRef]
- Ren, X.; Chen, C.M.; Ye, Y.X.; Xu, Z.Y.; Zhao, Q.L.; Luo, X.W.; Liu, Y.H.; Guo, P. Anti-inflammatory compounds from the mangrove endophytic fungus Amorosia sp. SCSIO 41026. Front. Microbiol. 2022, 13, 976399. [Google Scholar] [CrossRef] [PubMed]
- Lu, H.M.; Tan, Y.H.; Zhang, Y.T.; Li, Z.C.; Chen, J.Y.; Gao, C.H.; Liu, Y.H.; Luo, X.W. Osteoclastogenesis inhibitory phenolic derivatives produced by the Beibu Gulf coral-associated fungus Acremonium sclerotigenum GXIMD 02501. Fitoterapia 2022, 159, 105201. [Google Scholar] [CrossRef]
- Xing, D.X.; Song, X.S.; Pan, W.C.; Cui, H.; Zhao, Z.X. New chromone compounds from the marine derived fungus Diaporthe sp. XW12-1. Fitoterapia 2023, 164, 105384. [Google Scholar] [CrossRef] [PubMed]
- Yan, F.H.; Fang, J.B.; Ding, W.J.; Tang, X.Y.; Chen, X.M.; Ma, Z.J.; Wang, J.H. Structurally diverse metabolites from the marine-derived Streptomyces sp. DS-27 based on two different culture conditions. Chem. Biodivers. 2023, 20, 1017. [Google Scholar] [CrossRef]
- Jiang, M.; Zhang, Y.; Zhang, Y.X.; Ma, Z.J.; Wang, J.H. Suncheonosides E-M and benzothioate derivatives from the marine-derived Streptomyces sp. ZSN77. J. Nat. Prod. 2022, 85, 1771–1778. [Google Scholar] [CrossRef]
- Xie, X.T.; Xiao, S.P.; Liao, H.Y.; Jiang, Q.J.; Chen, G.; Wen, L. A new chloro-containing γ-butyrolactone from the mangrove endophytic fungus Neofusicoccum parvum Y2NBKZG1016. Chem. Nat. Compd. 2023, 59, 424–427. [Google Scholar] [CrossRef]
- Shin, H.J.; Anh, C.V.; Cho, D.Y.; Choi, D.K.; Kang, J.S.; Trinh, P.T.H.; Choi, B.-K.; Lee, H.-S. New polyenes from the marine-derived fungus Talaromyces cyanescens with anti-neuroinflammatory and cytotoxic activities. Molecules 2021, 26, 836. [Google Scholar] [CrossRef]
- He, J.L.; Wu, X.; Huang, S.H.; Wang, J.; Niu, S.W.; Chen, M.X.; Zhang, G.Y.; Cai, S.Y.; Wu, J.N.; Hong, B.H. Phenolic metabolites from a deep-sea-derived fungus Aspergillus puniceus A2 and their Nrf2-dependent anti-inflammatory effects. Mar. Drugs 2022, 20, 575. [Google Scholar] [CrossRef] [PubMed]
- Su, Y.S.; Cheng, M.J.; Wu, M.D.; Chai, C.Y.; Kwan, A.L.; Su, S.H.; Kuo, Y.H. Chemical constituents from a mangrove-derived Actinobacteria Isoptericola chiayiensis BCRC 16888 and evaluation of their anti-NO activity. Chem. Biodivers. 2021, 18, e2100211. [Google Scholar] [CrossRef]
- Chen, Y.; Zou, G.; Yang, W.C.; Zhao, Y.Y.; Tan, Q.; Chen, L.; Wang, J.M.; Ma, C.Y.; Kang, W.Y.; She, Z.G. Metabolites with anti-inflammatory activity from the mangrove endophytic fungus diaporthe sp. QYM12. Mar. Drugs 2021, 19, 56. [Google Scholar] [CrossRef]
- Dong, L.X.; Kim, H.J.; Cao, T.Q.; Liu, Z.M.; Lee, H.; Ko, W.; Kim, Y.C.; Sohn, J.H.; Kim, T.K.; Yim, J.H.; et al. Anti-inflammatory effects of metabolites from antarctic fungal strain Pleosporales sp. SF-7343 in HaCaT human keratinocytes. Int. J. Mol. Sci. 2021, 22, 9674. [Google Scholar] [CrossRef]
- Qin, X.Y.; Huang, J.G.; Zhou, D.X.; Zhang, W.X.; Zhang, Y.J.; Li, J.; Yang, R.Y.; Huang, X.S. Polyketide derivatives, guhypoxylonols A–D from a mangrove endophytic fungus Aspergillus sp. GXNU-Y45 that inhibit nitric oxide production. Mar. Drugs 2021, 20, 5. [Google Scholar] [CrossRef]
- Quang, T.H.; Vien, L.T.; Anh, L.N.; Ngan, N.T.T.; Hanh, T.T.H.; Cuong, N.X.; Nam, N.H.; Van Minh, C. Anti-inflammatory metabolites from a marine sponge-associated fungus Aspergillus sp. IMBC-FP2.05. Vietnam. J. Chem. 2021, 59, 52–56. [Google Scholar] [CrossRef]
- Chen, C.; Ye, G.T.; Tang, J.; Li, J.L.; Liu, W.B.; Wu, L.; Long, Y.H. New polyketides from mangrove endophytic fungus Penicillium sp. BJR-P2 and their anti-inflammatory activity. Mar. Drugs 2022, 20, 583. [Google Scholar] [CrossRef]
- Pang, X.Y.; Wang, P.; Liao, S.R.; Zhou, X.F.; Lin, X.P.; Yang, B.; Tian, X.P.; Wang, J.F.; Liu, Y.H. Three unusual hybrid sorbicillinoids with anti-inflammatory activities from the deep-sea derived fungus Penicillium sp. SCSIO 06868. Phytochemistry 2022, 202, 113311. [Google Scholar] [CrossRef]
- Cong, M.J.; Ren, X.; Song, Y.; Pang, X.Y.; Tian, X.P.; Liu, Y.H.; Guo, P.; Wang, J.F. Ochrathinols A and B, two pairs of sulfur-containing racemates from an antarctic fungus Aspergillus ochraceopetaliformis SCSIO 05702 inhibit LPS-induced pro-inflammatory cytokines and NO production. Phytochemistry 2023, 208, 113593. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Xie, C.L.; Wang, Y.; He, X.W.; Xie, M.M.; Li, Y.; Zhang, K.; Zou, Z.B.; Yang, L.H.; Xu, R.; et al. Penidihydrocitrinins A–C: New polyketides from the deep-sea-derived Penicillium citrinum W17 and their anti-inflammatory and anti-osteoporotic bioactivities. Mar. Drugs 2023, 21, 538. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Meng, Q.Y.; Niu, S.W.; Liu, J.; Guo, X.C.; Sun, Z.L.; Liu, D.; Gu, Y.C.; Huang, J.; Fan, A.L.; et al. Epigenetic manipulation to trigger production of guaiane-type sesquiterpenes from a marine-derived Spiromastix sp. fungus with anti-neuroinflammatory effects. J. Nat. Prod. 2021, 84, 1993–2003. [Google Scholar] [CrossRef] [PubMed]
- Li, P.H.; Zhang, M.Q.; Li, H.N.; Wang, R.C.; Hou, H.R.; Li, X.B.; Liu, K.C.; Chen, H. New prenylated indole homodimeric and pteridine alkaloids from the marine-derived fungus Aspergillus austroafricanus Y32-2. Mar. Drugs 2021, 19, 98. [Google Scholar] [CrossRef] [PubMed]
- Sun, B.Y.; Wang, D.D.; Ren, J.W.; Wang, C.J.; Yan, P.C.; Gustafson, K.R.; Jiang, W. Paraconulones A-G: Eremophilane sesquiterpenoids from the marine-derived fungus Paraconiothyrium sporulosum DL-16. J. Nat. Prod. 2023, 86, 1360–1369. [Google Scholar] [CrossRef]
- Niu, S.W.; Liu, D.; Shao, Z.Z.; Liu, J.R.; Fan, A.L.; Lin, W.H. Chemical epigenetic manipulation triggers the production of sesquiterpenes from the deep-sea derived Eutypella fungus. Phytochemistry 2021, 192, 112978. [Google Scholar] [CrossRef]
- Shang, R.Y.; Cui, J.; Li, J.X.; Miao, X.X.; Zhang, L.; Xie, D.D.; Zhang, L.; Lin, H.W.; Jiao, W.H. Nigerin and ochracenes J−L, new sesquiterpenoids from the marine sponge symbiotic fungus Aspergillus niger. Tetrahedron 2021, 104, 132599. [Google Scholar] [CrossRef]
- Safwan, S.; Wang, S.W.; Hsiao, G.; Hsiao, S.W.; Hsu, S.J.; Lee, T.H.; Lee, C.K. New trichothecenes isolated from the marine algicolous fungus Trichoderma brevicompactum. Mar. Drugs 2022, 20, 80. [Google Scholar] [CrossRef]
- Hu, Z.B.; Chen, J.J.; Liu, Q.Q.; Wu, Q.L.; Chen, S.H.; Wang, J.J.; Li, J.; Liu, L.; Gao, Z.Z. Cyclohexenone derivative and drimane sesquiterpenes from the seagrass-derived fungus Aspergillus insuetus. Chem. Biodivers. 2023, 20, e202300424. [Google Scholar] [CrossRef]
- Li, H.X.; Liu, X.Y.; Li, X.F.; Hu, Z.L.; Wang, L.Y. Novel harziane diterpenes from deep-sea sediment fungus Trichoderma sp. SCSIOW 21 and their potential anti-inflammatory effects. Mar. Drugs 2021, 19, 689. [Google Scholar] [CrossRef]
- Ning, Y.D.; Zhang, S.; Zheng, T.; Xu, Y.; Li, S.; Zhang, J.P.; Jiao, B.H.; Zhang, Y.; Ma, Z.L.; Lu, X.L. Pimarane-type diterpenes with anti-inflammatory activity from arctic-derived fungus Eutypella sp. D-1. Mar. Drugs 2023, 21, 541. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.; Chen, X.C.; Pan, W.C.; Liu, X.; Tan, S.L.; Cui, H.; Zhao, Z.X. Meroterpenoids from the fungus Penicillium sclerotiorum GZU-XW03-2 and their anti-inflammatory activity. Phytochemistry 2022, 202, 113307. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Zhao, M.; Chen, J.; Pan, W.C.; Tan, S.L.; Cui, H.; Zhao, Z.X. Seven new meroterpenoids from the fungus Penicillium sclerotiorum GZU-XW03-2. Fitoterapia 2023, 165, 105428. [Google Scholar] [CrossRef]
- Yu, H.; Chen, Y.H.; Cheng, Z.; Li, H.J.; Bian, H.H.; Yang, X.; Lv, J.; Liu, W.; Su, L.; Sun, P. Anti-inflammatory oleanane-type triterpenoids produced by Nonomuraea sp. MYH 522 through microbial transformation. J. Agric. Food Chem. 2023, 71, 3777–3789. [Google Scholar] [CrossRef] [PubMed]
- Thambi, A.; Chakraborty, K. Anti-inflammatory decurrencyclics A–B, two undescribed nor-dammarane triterpenes from triangular sea bell Turbinaria decurrens. Nat. Prod. Rep. 2022, 37, 713–724. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.Q.; Chen, X.C.; Zhou, Y.W.; Zhao, M.; He, J.X.; Liu, Y.N.; Chen, G.Y.; Zhao, Z.X.; Cui, H. Furanaspermeroterpenes A and B, two unusual meroterpenoids with a unique 6/6/6/5/5 pentacyclic skeleton from the marine-derived fungus Aspergillus terreus GZU-31-1. Bioorg. Chem. 2021, 114, 105111. [Google Scholar] [CrossRef]
- Liu, Z.M.; Chen, Y.C.; Li, S.N.; Hu, C.Y.; Liu, H.X.; Zhang, W.M. Indole diketopiperazine alkaloids from the deep-sea-derived fungus Aspergillus sp. FS 445. Nat. Prod. Res. 2021, 36, 5213–5221. [Google Scholar] [CrossRef]
- Niveditha, L.; Fu, P.; Leao, T.F.; Li, T.; Wang, T.; Poulin, R.X.; Gaspar, L.R.; Naman, C.B.; Puthiyedathu, T.S. Targeted isolation of two new anti-inflammatory and UV-a protective dipyrroloquinones from the sponge-associated fungus Aspergillus tamarii MCCF 102. Planta Med. 2022, 88, 774–782. [Google Scholar]
- Lee, H.S.; Kang, J.S.; Cho, D.Y.; Choi, D.K.; Shin, H.J. Isolation, structure determination, and semisynthesis of diphenazine compounds from a deep-sea-derived strain of the fungus Cystobasidium laryngis and their biological activities. J. Nat. Prod. 2022, 85, 857–865. [Google Scholar] [CrossRef]
- Xu, X.Y.; Tan, Y.H.; Gao, C.H.; Liu, K.; Tang, Z.Z.; Lu, C.J.; Li, H.Y.; Zhang, X.Y.; Liu, Y.H. New 3-acyl tetramic acid derivatives from the deep-sea-derived fungus Lecanicillium fusisporum. Mar. Drugs 2022, 20, 255. [Google Scholar] [CrossRef]
- Zhang, D.; Xu, J.; Qin, Q.; An, F.L.; Wang, S.P.; Li, L.; Lin, H.W. Marinacarboline glucuronide, a new member of β-carboline alkaloids from sponge-derived actinomycete Actinoalloteichus cyanogriseus LHW 52806. J. Antibiot. 2022, 75, 523–525. [Google Scholar] [CrossRef] [PubMed]
- Yao, G.S.; Ma, Z.L.; Zheng, Y.Y.; Lv, L.; Mao, J.Q.; Wang, C.Y. Bioactive alkaloids from the marine-derived fungus Metarhizium sp. P2100. J. Fungi 2022, 8, 1218. [Google Scholar] [CrossRef] [PubMed]
- Meng, Q.Y.; Guo, X.; Wu, J.S.; Liu, D.; Gu, Y.C.; Huang, J.; Fan, A.L.; Lin, W.H. Prenylated notoamide-type alkaloids isolated from the fungus Aspergillus sclerotiorum and their inhibition of NLRP3 inflammasome activation and antibacterial activities. Phytochemistry 2022, 203, 113424. [Google Scholar] [CrossRef]
- Xu, L.; Guo, F.W.; Zhang, X.Q.; Zhou, T.Y.; Wang, C.J.; Wei, M.Y.; Gu, Y.C.; Wang, C.Y.; Shao, C.L. Discovery, total syntheses and potent anti-inflammatory activity of pyrrolinone-fused benzoazepine alkaloids asperazepanones A and B from Aspergillus candidus. Commun. Chem. 2022, 5, 80. [Google Scholar] [CrossRef]
- Yang, J.; Gong, L.Z.; Guo, M.M.; Jiang, Y.; Ding, Y.; Wang, Z.J.; Xin, X.J.; An, F.L. Bioactive indole diketopiperazine alkaloids from the marine endophytic fungus Aspergillus sp. YJ191021. Mar. Drugs 2021, 19, 157. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Li, Y.M.; Chen, M.Q.; Liu, Y.Y.; Liang, J.Y.; Zhang, Y.; Qian, Z.J. Mechanism of two alkaloids isolated from coral endophytic fungus for suppressing angiogenesis in atherosclerotic plaque in HUVEC. Int. Immunopharmacol. 2022, 109, 108931. [Google Scholar] [CrossRef]
- Li, S.H.; Ma, Y.H.; Wang, L.X.; Lan, D.H.; Fu, L.L.; Wu, B. Two new alkaloids from the marine-derived fungus Penicillium sp. LSH-3-1. Chem. Biodivers. 2022, 19, e202200310. [Google Scholar] [CrossRef]
- Elhady, S.S.; Goda, M.S.; Mehanna, E.T.; Elfaky, M.A.; Koshak, A.E.; Noor, A.O.; Bogari, H.A.; Malatani, R.T.; Abdelhameed, R.F.A.; Wahba, A.S. Meleagrin isolated from the red sea fungus Penicillium chrysogenum protects against bleomycin-induced pulmonary fibrosis in mice. Biomedicines 2022, 10, 1164. [Google Scholar] [CrossRef] [PubMed]
- Anh, C.V.; Yoon, Y.D.; Kang, J.S.; Lee, H.S.; Heo, C.S.; Shin, H.J. Nitrogen-containing secondary metabolites from a deep-sea fungus Aspergillus unguis and their anti-inflammatory activity. Mar. Drugs 2022, 20, 217. [Google Scholar] [CrossRef]
- Chen, Y.H.; Wu, X.N.; Xu, L.; El-Shazly, M.; Ma, C.W.; Yuan, S.J.; Wang, P.P.; Luo, L.Z. Two new cerebroside metabolites from the marine fungus Hortaea werneckii. Chem. Biodivers. 2022, 19, e202200008. [Google Scholar] [CrossRef]
- Yang, S.C.; Wang, Y.H.; Ho, C.M.; Tsai, Y.F.; Sung, P.J.; Lin, T.E.; Hwang, T.L. Targeting formyl peptide receptor 1 with anteiso-C13-surfactin for neutrophil-dominant acute respiratory distress syndrome. Br. J. Pharmacol. 2023, 180, 2120–2139. [Google Scholar] [CrossRef] [PubMed]
- Chingizova, E.A.; Menchinskaya, E.S.; Chingizov, A.R.; Pislyagin, E.A.; Girich, E.V.; Yurchenko, A.N.; Guzhova, I.V.; Mikhailov, V.V.; Aminin, D.L.; Yurchenko, E.A. Marine fungal cerebroside flavuside B protects HaCaT keratinocytes against staphylococcus aureus induced damage. Mar. Drugs 2021, 19, 553. [Google Scholar] [CrossRef] [PubMed]
- Li, J.X.; Tao, H.M.; Lei, X.X.; Zhang, H.; Zhou, X.F.; Liu, Y.H.; Li, Y.Q.; Yang, B. Arthriniumsteroids A-D, four new steroids from the soft coral-derived fungus Simplicillium lanosoniveum SCSIO 41212. Steroids 2021, 171, 108831. [Google Scholar] [CrossRef] [PubMed]
- Hoang, C.K.; Le, C.H.; Nguyen, D.T.; Tran, H.T.N.; Luu, C.V.; Le, H.M.; Tran, H.T.H. Steroid components of marine-derived fungal strain Penicillium levitum N33.2 and their biological activities. Mycobiology 2023, 51, 246–255. [Google Scholar] [CrossRef]
- Cao, V.A.; Kwon, J.H.; Kang, J.S.; Lee, H.S.; Heo, C.S.; Shin, H.J. Aspersterols A–D, ergostane-type sterols with an unusual unsaturated side chain from the deep-sea-derived fungus Aspergillus unguis. J. Nat. Prod. 2022, 85, 2177–2183. [Google Scholar] [CrossRef]
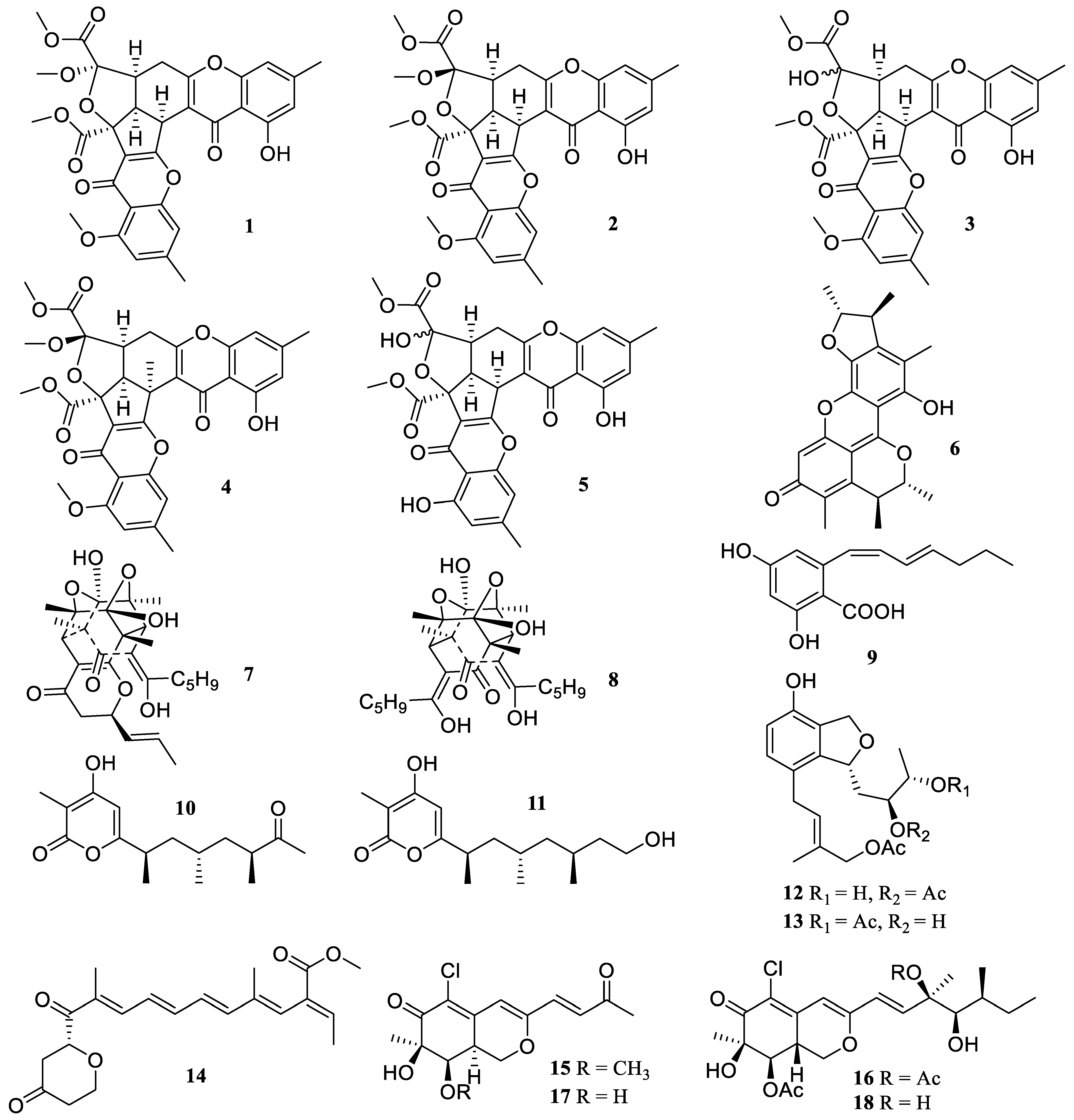




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, G.; Lin, M.; Kaliaperumal, K.; Lu, Y.; Qi, X.; Jiang, X.; Xu, X.; Gao, C.; Liu, Y.; Luo, X. Recent Advances in Anti-Inflammatory Compounds from Marine Microorganisms. Mar. Drugs 2024, 22, 424. https://doi.org/10.3390/md22090424
Yang G, Lin M, Kaliaperumal K, Lu Y, Qi X, Jiang X, Xu X, Gao C, Liu Y, Luo X. Recent Advances in Anti-Inflammatory Compounds from Marine Microorganisms. Marine Drugs. 2024; 22(9):424. https://doi.org/10.3390/md22090424
Chicago/Turabian StyleYang, Guihua, Miaoping Lin, Kumaravel Kaliaperumal, Yaqi Lu, Xin Qi, Xiaodong Jiang, Xinya Xu, Chenghai Gao, Yonghong Liu, and Xiaowei Luo. 2024. "Recent Advances in Anti-Inflammatory Compounds from Marine Microorganisms" Marine Drugs 22, no. 9: 424. https://doi.org/10.3390/md22090424
APA StyleYang, G., Lin, M., Kaliaperumal, K., Lu, Y., Qi, X., Jiang, X., Xu, X., Gao, C., Liu, Y., & Luo, X. (2024). Recent Advances in Anti-Inflammatory Compounds from Marine Microorganisms. Marine Drugs, 22(9), 424. https://doi.org/10.3390/md22090424