
Construction of Specific Primers for Rapid Detection of South African Exportable Vegetable Macergens



Abstract
:
1. Introduction
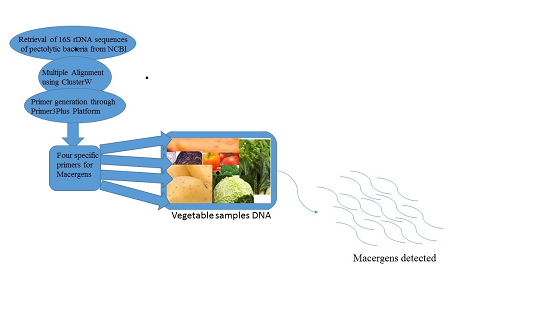
2. Materials and Methods
2.1. Primer Design
2.2. Primer Development
3. Detection of Macergens from Vegetable Samples
3.1. Extraction of Metagenomic DNA from Vegetables
3.2. PCR Amplification
3.3. DNA Sequencing
3.4. Sequence Analysis
3.5. Phylogenetic Analysis
4. Results and Discussions

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Primer Set | Oligonucleotide Sequence | GC % | Tm | Length | Location | Position |
|---|---|---|---|---|---|---|---|
| M101F | Set 1 | CGGACGGGTGAGTAATGTCT | 55 | 56.5 | 20 | 16S | 101-121 |
| M1208R | Set 1 | AAGGGCCATGATGACTTGAC | 50 | 55.1 | 20 | 16S | 1208-1180 |
| M182F | Set 2 | CGATCCCTAGCTGGTCTGAG | 60 | 60.0 | 20 | 16S | 182-202 |
| M1190R | Set 2 | TTATGAGGTCCGCTTGCTCT | 50 | 60.0 | 20 | 16S | 1190-1170 |
| M180F | Set 3 | GACGATCCCTAGCTGGTCTG | 60 | 56.9 | 20 | 16S | 180-200 |
| M1190R | Set 3 | TTATGAGGTCCGCTTGCTCT | 50 | 56.0 | 20 | 16S | 1190-1170 |
| M57F | Set 4 | GAGGAAGAAACCGGCGATAG | 55 | 55.3 | 20 | 16S | 57-77 |
| M296R | Set 4 | GGCGTATCCACCGATGTAAT | 50 | 54.6 | 20 | 16S | 296-279 |



| Lanes | Vegetable Source | Macergens | Clone Codes | Accession Number | References |
|---|---|---|---|---|---|
| 3 | Straight Baby Marrow | Uncultured Kluyvera sp. | M111 | KP114439 | This Study |
| 5 | White Button Mushroom | Uncultured Enterobacter sp. | M112 | KP114440 | This Study |
| 9 | Straight Small Marrow | Uncultured Citrobacter sp. | M113 | KP114441 | This Study |
| 10 | Round Baby Marrow | Uncultured Pseudomonas sp. | M114 | KP114442 | This Study |
| 11 | Red Cabbage | Uncultured Klebsiella sp. | M115 | KP114443 | This Study |
| 15 | Parsley | Uncultured Pantoea sp. | M116 | KP114444 | This Study |
| 16 | Potatoes | Uncultured Pseudomonas sp. | M117 | KP114445 | This Study |
| 21 | Spinach | Uncultured Citrobacter sp. | M118 | KP114446 | This Study |
| 22 | Spring Onions | Uncultured Lelliottia sp. | M119 | KP114447 | This Study |
| 26 | Bell Paper | Uncultured Tatumella sp. | M120 | KP114448 | This Study |
| Lanes | Vegetable Source | Macergens | Clone Codes | Accession Number | References |
|---|---|---|---|---|---|
| 1 | White Cabbage | Uncultured Enterobacter sp. | M20 | KM924134 | This Study |
| 3 | Straight Baby Marrow | Uncultured Enterobacter sp. | M21 | KM924135 | This Study |
| 5 | White Button Mushroom | Uncultured Cedecea sp. | M22 | KM924136 | This Study |
| 9 | Straight Small Marrow | Uncultured Citrobacter sp. | M23 | KM924137 | This Study |
| 10 | Round Baby Marrow | Uncultured Citrobacter sp. | M24 | KM924138 | This Study |
| 11 | Red Cabbage | Uncultured Klebsiella sp. | M25 | KM924139 | This Study |
| 12 | Iceberg Lettuce | Uncultured Pectobacterium sp. | M26 | KM924140 | This Study |
| 13 | Cauliflower | Uncultured Citrobacter sp. | M27 | KM924141 | This Study |
| 15 | Parsley | Uncultured Leclercia sp. | M28 | KM924142 | This Study |
| 16 | Potatoes | Uncultured Rahnella sp. | M29 | KM924143 | This Study |
| 18 | Potatoes | Uncultured Lelliottia sp. | M30 | KM924144 | This Study |
| 26 | Bell Pepper | Uncultured Tatumella sp. | M31 | KM924145 | This Study |
| Lanes | Vegetable Source | Macergens | Strains Codes | Accession Number | References |
|---|---|---|---|---|---|
| 1 | White Cabbage | Uncultured Pectobacterium carotovorum | M32 | KP792433 | This study |
| 3 | Straight Baby Marrow | Uncultured Acinetobacter calcoaceticus | M33 | KP792434 | This study |
| 4 | Beetroot | Uncultured Cronobacter malonaticus | M34 | KP792435 | This study |
| 11 | Red Cabbage | Uncultured Klebsiella pneumoniae | M35 | KP792436 | This study |
| 12 | Iceberg Lettuce | Uncultured Pectobacterium sp. | M36 | KP792437 | This study |
| 17 | Celery | Uncultured Lelliottia amnigena | M37 | KP792438 | This study |
| 18 | Potatoes | Uncultured Enterobacter sp. | M38 | KP792439 | This study |
| 20 | Potatoes | Uncultured Leclercia adecarboxylata | M39 | KP792440 | This study |
| 21 | Spinach | Uncultured Enterobacter sp. | M40 | KP792441 | This study |


5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviation
| DNA | Deoxy Ribonucleic Acid |
| PCR | Polymerase Chain Reaction |
| RNA | Ribonucleic Acid |
| rDNA | Ribosomal Deoxy Ribonucleic Acid |
| rRNA | Ribosomal Ribonucleic Acid |
References
- Beattie, G.A. Plant-associated bacteria: Survey, molecular phylogeny, genomics and recent advances. In Plant-Associated Bacteria; Gnanamanickam, S.S., Ed.; Springer: Dordrecht, The Netherlands, 2006; pp. 1–56. [Google Scholar]
- Sobiczewski, P. Bacterial diseases of plants: Epidemiology, diagnostics and control. Zemdirbyste 2008, 95, 151–157. [Google Scholar]
- Saxena, N. Studies on Cell Wall Degrading Enzymes in Relation to Fungal Pathogenesis. Ph.D. Thesis, University of Saugar, Sagar, India, 2015. [Google Scholar]
- Mikiciński, A.; Sobiczewski, P.; Sulikowska, M.; Puławska, J.; Treder, J. Pectolytic bacteria associated with soft rot of calla lily (Zantedeschia spp.) tubers. J. Phytopathol. 2010, 158, 201–209. [Google Scholar] [CrossRef]
- Tarkowski, P.; Vereecke, D. Threats and opportunities of plant pathogenic bacteria. Biotechnol. Adv. 2014, 32, 215–229. [Google Scholar] [CrossRef] [PubMed]
- Howard, D.G. Heirloom Flavor: Yesterday’s Best-Tasting Vegetables, Fruits, and Herbs for Today’s Cook; Cool Springs Press: Brentwood, TN, USA, 2013. [Google Scholar]
- Dong, Y.H.; Zhang, X.F.; Xu, J.L.; Zhang, L.H. Insecticidal Bacillus thuringiensis silences Erwinia carotovora virulence by a new form of microbial antagonism, signal interference. Appl. Environ. Microbiol. 2004, 70, 954–960. [Google Scholar] [CrossRef] [PubMed]
- Yeni, F.; Acar, S.; Polat, Ö.; Soyer, Y.; Alpas, H. Rapid and standardized methods for detection of foodborne pathogens and mycotoxins on fresh produce. Food Control 2014, 40, 359–367. [Google Scholar] [CrossRef]
- Hall, T. BioEdit. Ibis Therapeutics. Available online: http://www.mbio.ncsu.edu/BioEdit/bioedit.html (accessed on 18 March 2005).
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; et al. ClustalW and ClustalX version 2.0. Bioinform. Biol. Insights 2007, 23, 2947–2948. [Google Scholar]
- Innis, M.A.; Gelfand, D.H. Optimization of PCRs. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: San Diego, CA, USA, 2012; pp. 3–12. [Google Scholar]
- Technelysium. Chromas. Available online: http://www.technelysium.com.au/chromas.html (accessed on 18 March 2005).
- Altschul, S.F.; Madden, T.L.; Schäffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Toh, H. Parallelization of the MAFFT multiple sequence alignment program. Bioinformatics 2010, 26, 1899–1900. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed]
- Jill Harrison, C.; Langdale, J.A. A step by step guide to phylogeny reconstruction. Plant J. 2006, 45, 561–572. [Google Scholar] [CrossRef] [PubMed]
- Clarridge, J.E. Impact of 16S rRNA gene sequence analysis for identification of bacteria on clinical microbiology and infectious diseases. Clin. Microbiol. Rev. 2004, 17, 840–862. [Google Scholar] [CrossRef] [PubMed]
- Drancourt, M.; Bollet, C.; Carlioz, A.; Martelin, R.; Gayral, J.-P.; Raoult, D. 16S ribosomal DNA sequence analysis of a large collection of environmental and clinical unidentifiable bacterial isolates. J. Clin. Microbiol. 2000, 38, 3623–3630. [Google Scholar] [PubMed]
- Konstantinidis, K.T.; Stackebrandt, E. Defining taxonomic ranks. In The Prokaryotes, 4th ed.; Rosenberg, E., DeLong, E.F., Stackebrandt, E., Lory, S., Thompson, F., Eds.; Springer Verlag: Berlin, Germany; Heidelberg, Germany, 2013; Volume 2, pp. 229–254. [Google Scholar]
- Tindall, B.; Rossello-Mora, R.; Busse, H.; Ludwig, W.; Kampfer, P. Notes on the characterization of prokaryote strains for taxonomic purposes. Int. J. Syst. Evol. Microbiol. 2010, 60, 249–266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nabhan, S.; Wydra, K.; Linde, M.; Debener, T. The use of two complementary DNA assays, AFLP and MLSA, for epidemic and phylogenetic studies of pectolytic enterobacterial strains with focus on the heterogeneous species Pectobacterium carotovorum. Plant Pathol. 2012, 61, 498–508. [Google Scholar] [CrossRef]
- Brady, C.L.; Cleenwerck, I.; Denman, S.; Venter, S.N.; Rodríguez-Palenzuela, P.; Coutinho, T.A.; De Vos, P. Proposal to reclassify Brenneria quercina (Hildebrand and Schroth 1967) Hauben et al. 1999 into a new genus, Lonsdalea gen. nov., as Lonsdalea quercina comb. nov., descriptions of Lonsdalea quercina subsp. quercina comb. nov., Lonsdalea quercina subsp. iberica subsp. nov., and Lonsdalea quercina subsp. britannica subsp. nov., emendation of the description of the genus Brenneria, reclassification of Dickeya dieffenbachiae as Dickeya dadantii subsp. dieffenbachiae comb. nov., and emendation of the description of Dickeya dadantii. Int. J. Syst. Evol. Microbiol. 2012, 62, 1592–1602. [Google Scholar]
- Wayne, L.; Brenner, D.; Colwell, R.; Grimont, P.; Kandler, O.; Krichevsky, M.; Moore, L.; Moore, W.; Murray, R.; Stackebrandt, E.; et al. International committee on systematic bacteriology: Report of the ad hoc committee on reconciliation of approaches to bacterial systematics. Int. J. Syst. Evol. Microbiol. 1987, 37, 463–464. [Google Scholar]
- Stackebrandt, E.; Frederiksen, W.; Garrity, G.M.; Grimont, P.A.D.; Kämpfer, P.; Maiden, M.C.J.; Nesme, X.; Rosselló-Mora, R.; Swings, J.; Trüper, H.G. Report of the ad hoc committee for the re-evaluation of the species definition in bacteriology. Int. J. Syst. Evol. Microbiol. 2002, 52, 1043–1047. [Google Scholar] [CrossRef] [PubMed]
- Togashi, J.; Koushi, U.; Namai, T. Overwintering of Erwinia carotovora subsp. carotovora in diseased tissues in soil and its role as inoculum for soft rot of chinese cabbage (Brassica campestris, Pekinensis group). J. Gen. Plant Pathol. 2001, 67, 45–50. [Google Scholar]
- Kimura, M. A simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef] [PubMed]
- Kalia, V.C.; Mukherjee, T.; Bhushan, A.; Joshi, J.; Shankar, P.; Huma, N. Analysis of the unexplored features of rrs (16S rDNA) of the genus Clostridium. BMC Genom. 2011, 12. [Google Scholar] [CrossRef] [PubMed]
- Naushad, H.S.; Lee, B.; Gupta, R.S. Conserved signature indels and signature proteins as novel tools for understanding microbial phylogeny and systematics: Identification of molecular signatures that are specific for the phytopathogenic genera Dickeya, Pectobacterium and Brenneria. Int. J. Syst. Evol. Microbiol. 2014, 64, 366–383. [Google Scholar] [CrossRef] [PubMed]
- Stackebrandt, E.; Goebel, B. Taxonomic note: A place for DNA-DNA reassociation and 16S rRNA sequence analysis in the present species definition in bacteriology. Int. J. Syst. Bacteriol. 1994, 44, 846–849. [Google Scholar] [CrossRef]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aremu, B.R.; Babalola, O.O. Construction of Specific Primers for Rapid Detection of South African Exportable Vegetable Macergens. Int. J. Environ. Res. Public Health 2015, 12, 12356-12370. https://doi.org/10.3390/ijerph121012356
Aremu BR, Babalola OO. Construction of Specific Primers for Rapid Detection of South African Exportable Vegetable Macergens. International Journal of Environmental Research and Public Health. 2015; 12(10):12356-12370. https://doi.org/10.3390/ijerph121012356
Chicago/Turabian StyleAremu, Bukola Rhoda, and Olubukola Oluranti Babalola. 2015. "Construction of Specific Primers for Rapid Detection of South African Exportable Vegetable Macergens" International Journal of Environmental Research and Public Health 12, no. 10: 12356-12370. https://doi.org/10.3390/ijerph121012356
APA StyleAremu, B. R., & Babalola, O. O. (2015). Construction of Specific Primers for Rapid Detection of South African Exportable Vegetable Macergens. International Journal of Environmental Research and Public Health, 12(10), 12356-12370. https://doi.org/10.3390/ijerph121012356