
Threshold Dose of Three Types of Quantum Dots (QDs) Induces Oxidative Stress Triggers DNA Damage and Apoptosis in Mouse Fibroblast L929 Cells

Abstract
:1. Introduction
2. Experimental Section
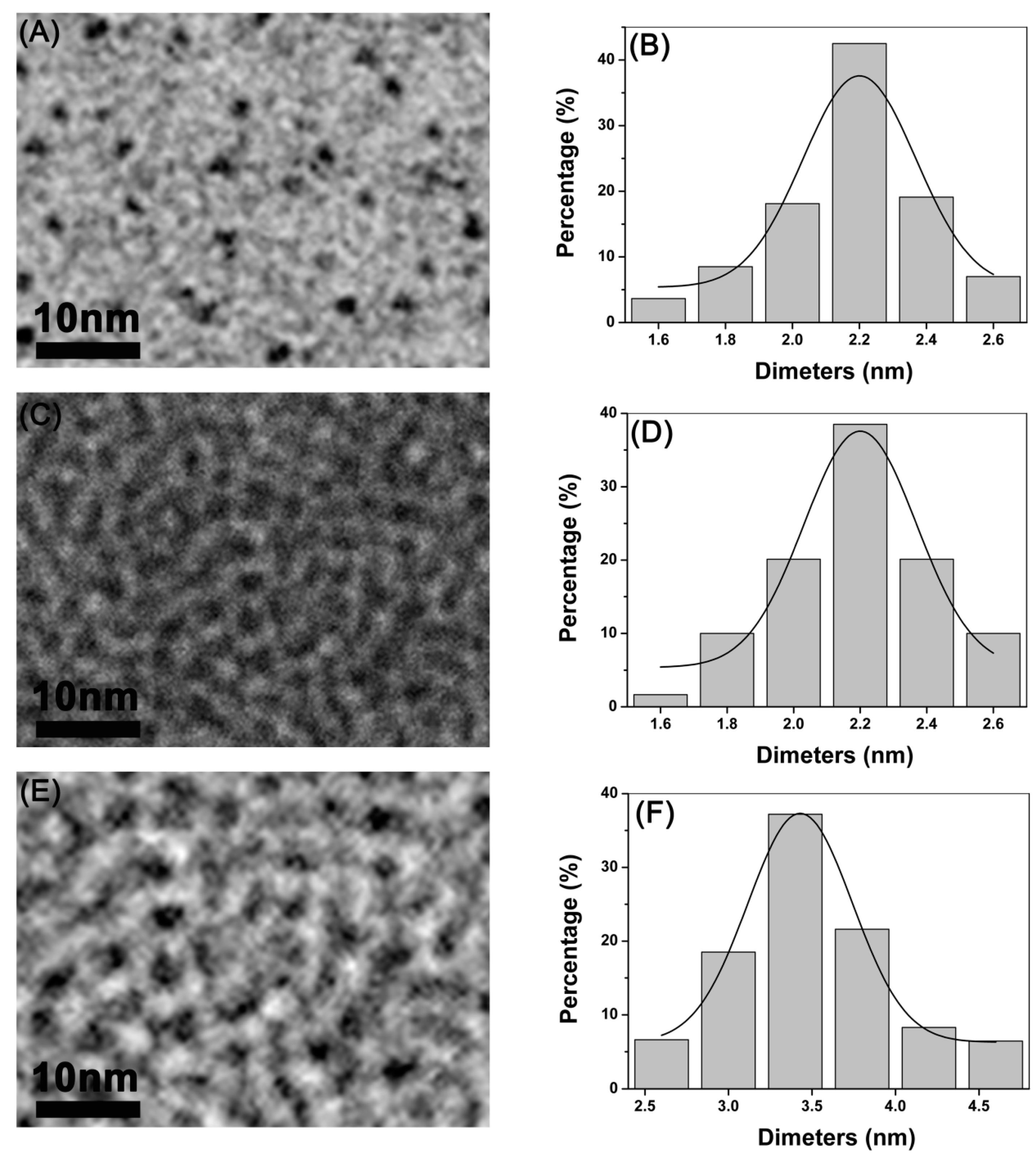
2.1. Preparation and Characterization of CdTe/CdSe QDs
2.2. Hemolysis Test
2.3. Cell Culture and Treatments
2.4. Viability Analysis
2.5. Microscopic Observations
2.6. LDH Release Assay
2.7. Oxidative Damage
2.8. Annexin V-FITC/Propidium Iodide Apoptosis Assay
2.9. Comet Assay
2.10. Statistical Analysis
3. Results
3.1. Synthesis and Characterization of CdTe/CdSe QDs

3.2. Determination of the Hemolytic Activity

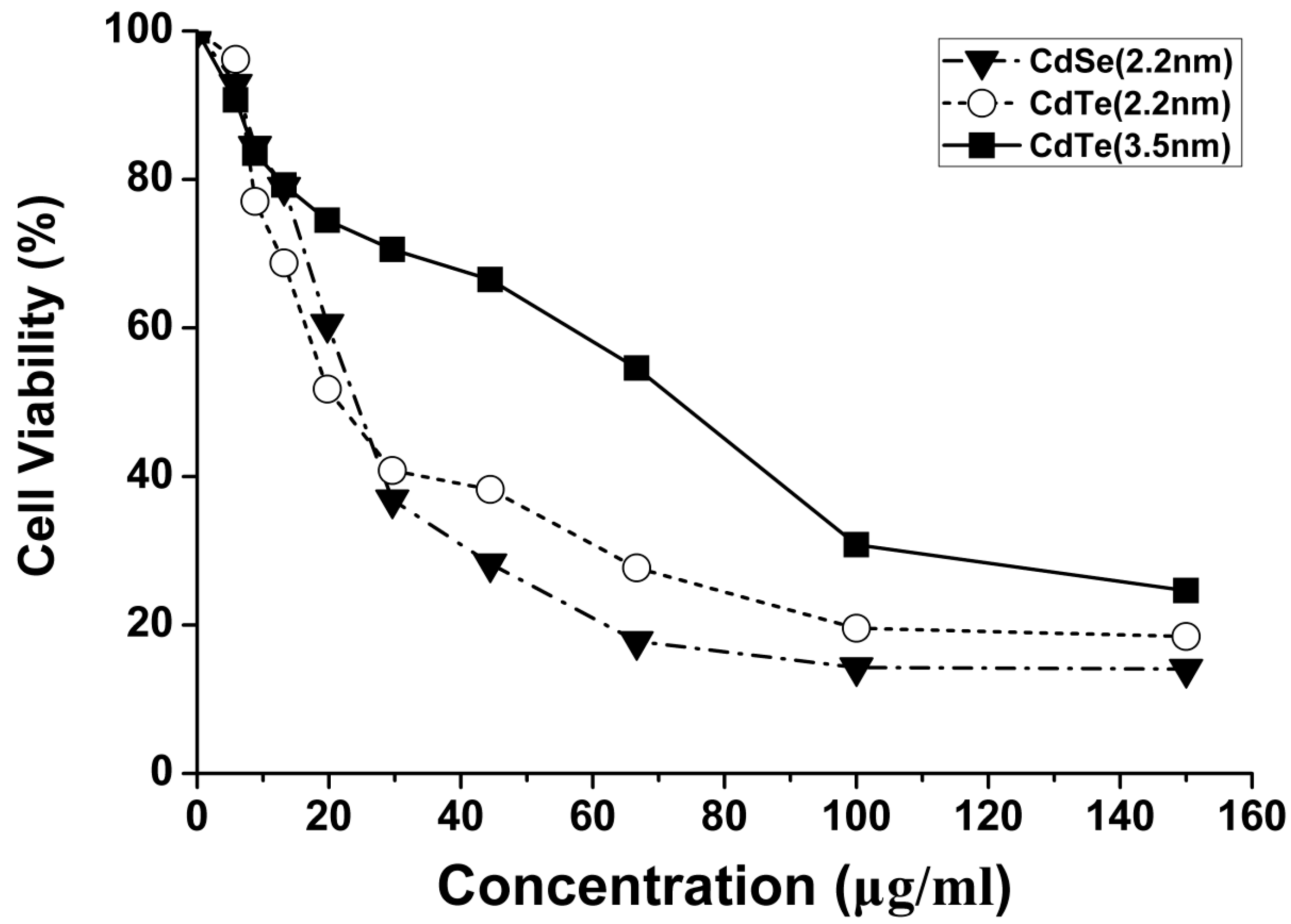
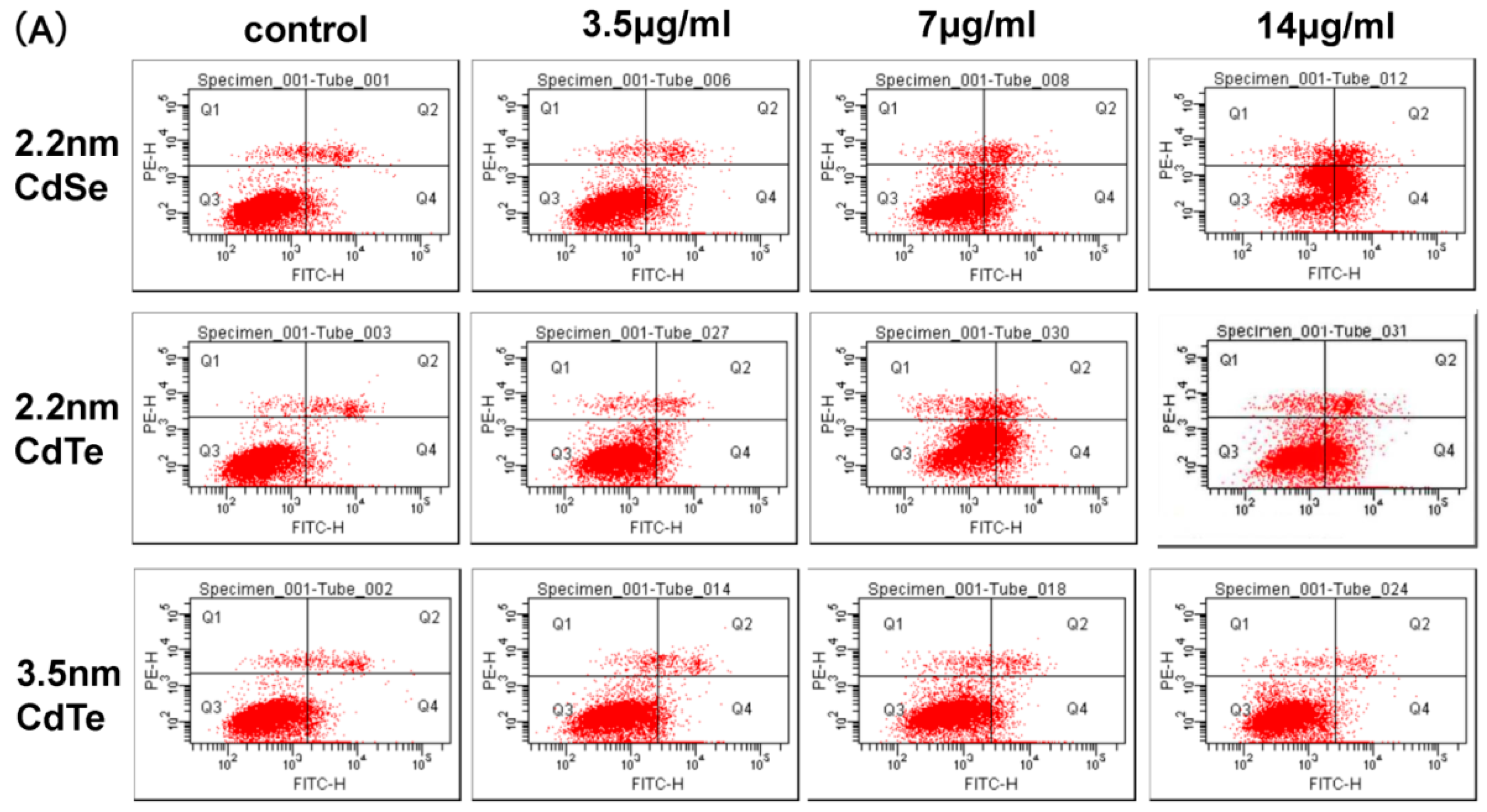
3.3. Concentration- and Size-Dependent Cytotoxicity of QDs Induced in L929 Mouse Fibroblasts


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| QDs | IC20 | IC50 |
|---|---|---|
| 2.2 nm CdSe QDs | 10.84 | 27.16 |
| 2.2 nm CdTe QDs | 7.38 | 25.83 |
| 3.5 nm CdTe QDs | 15.53 | 59.82 |
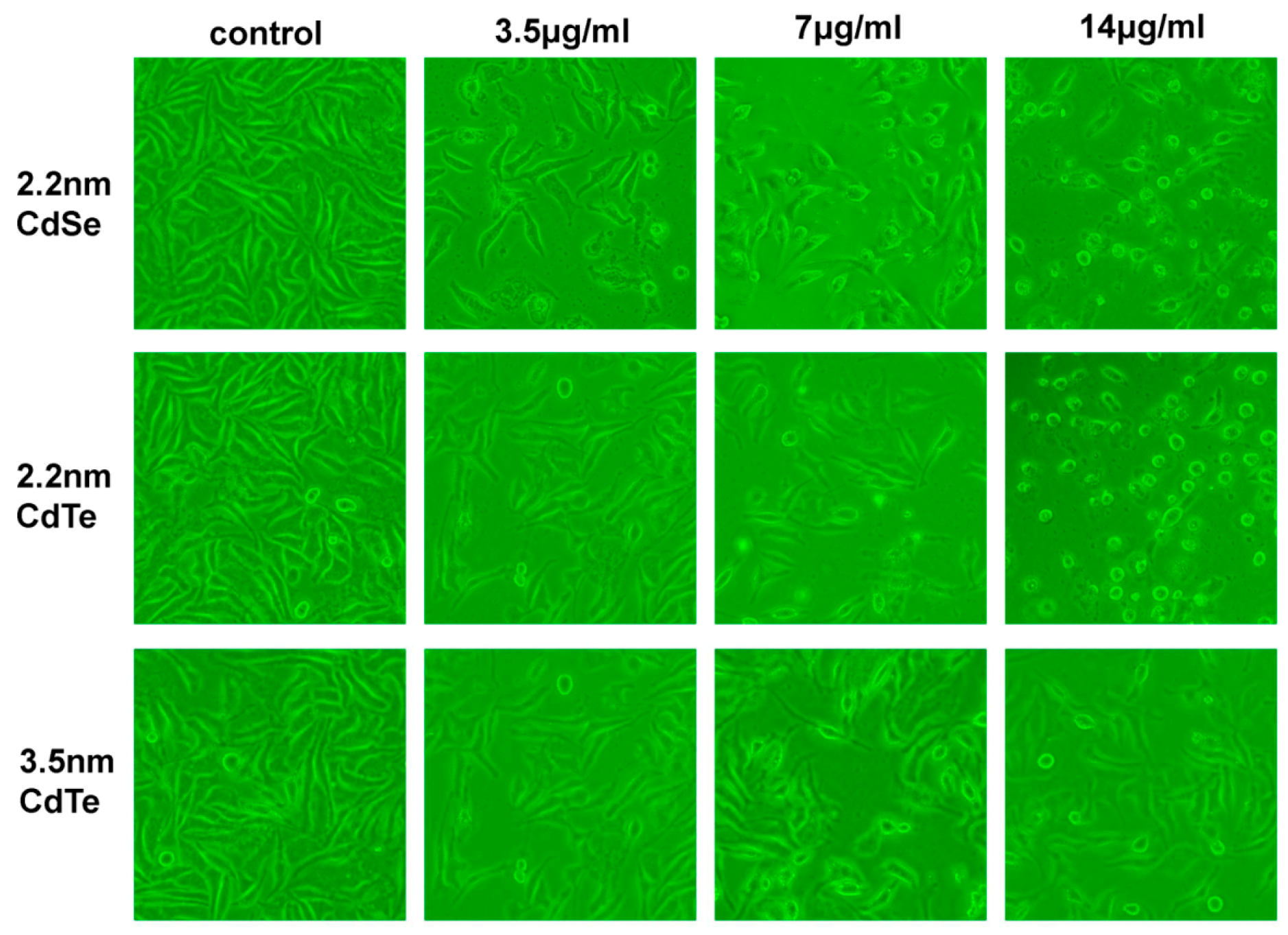
3.4. Change of Cellular Morphology
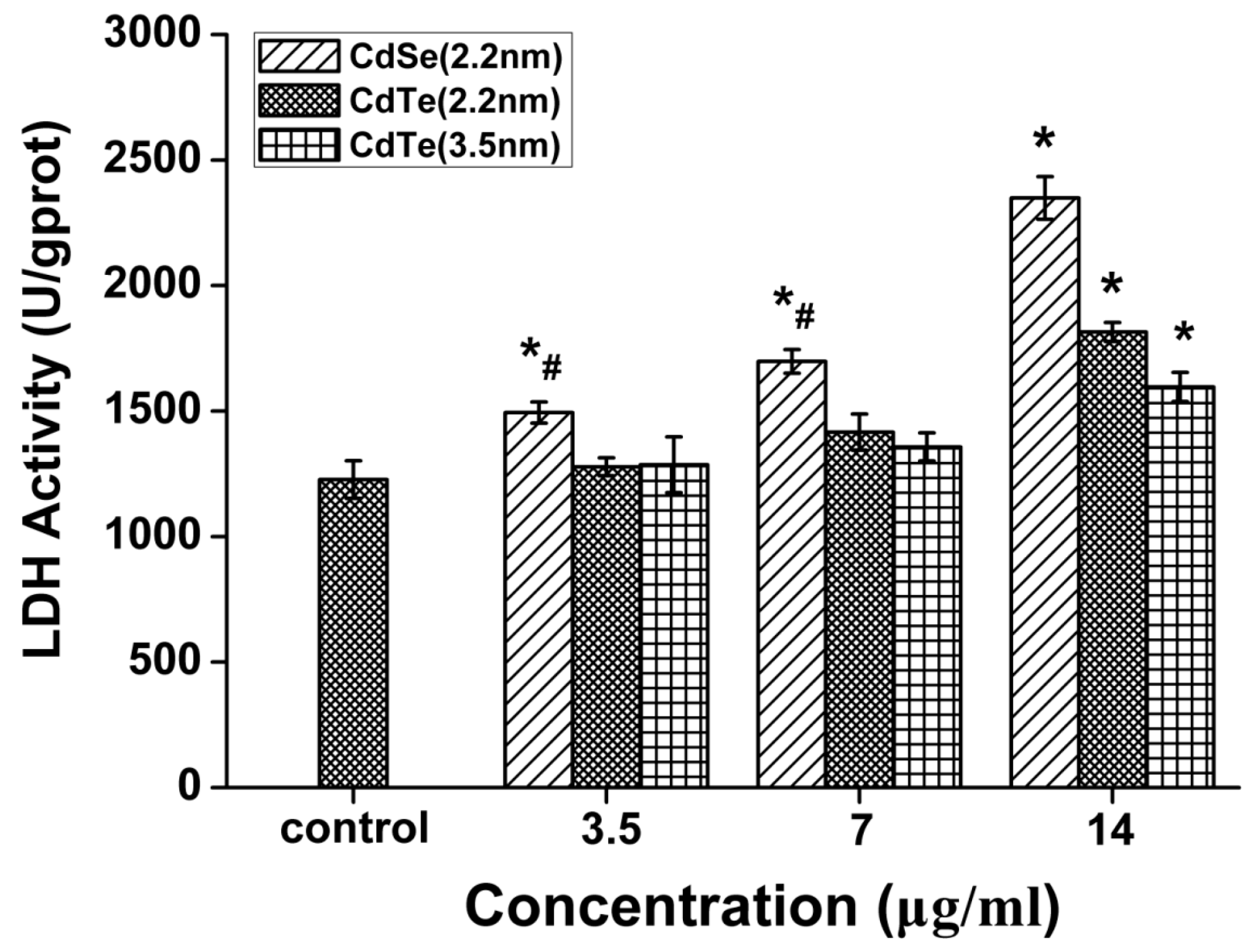
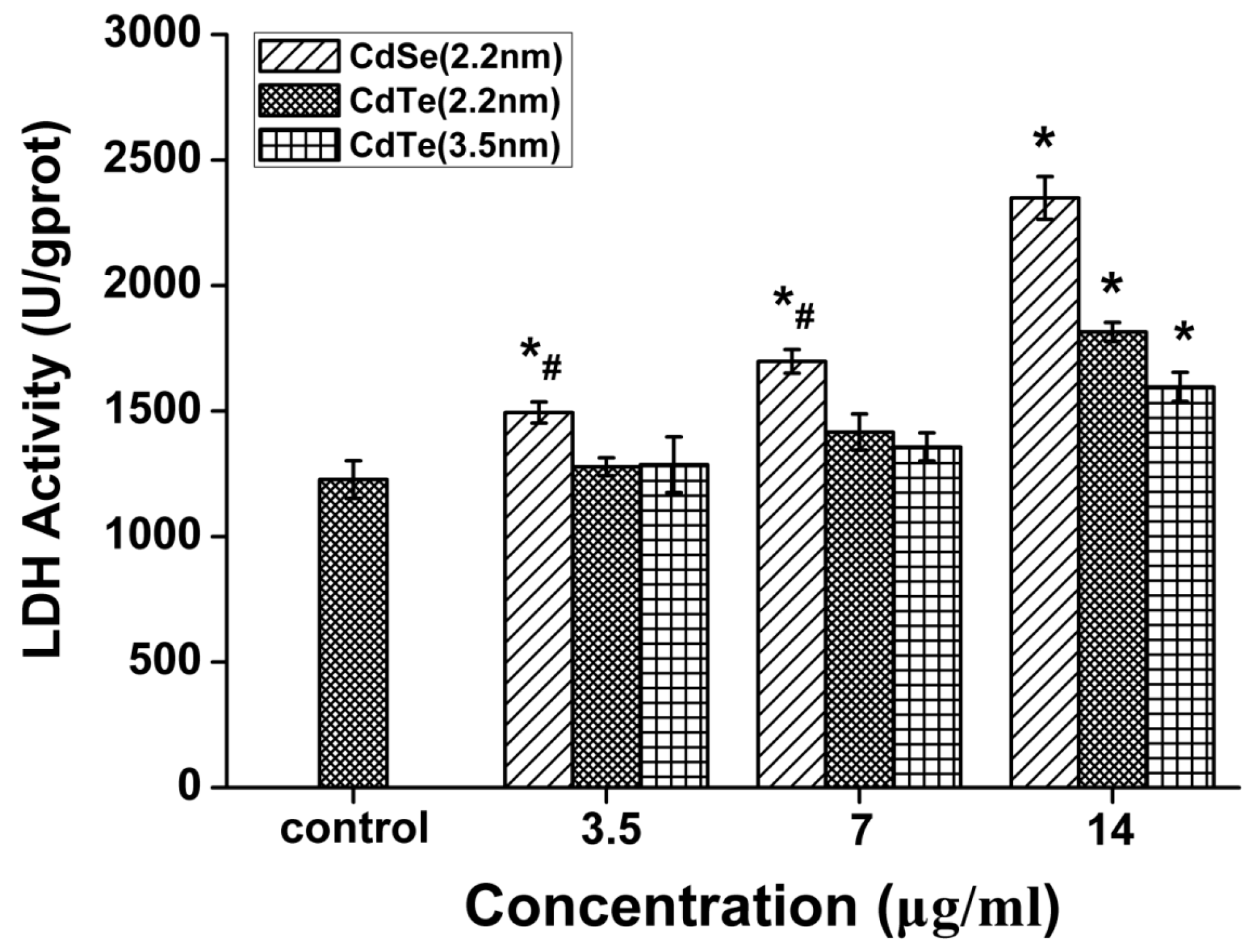
3.5. QDs Caused Membrane Damage in L929 Mouse Fibroblasts Detected by LDH Assay

3.6. Oxidative Stress-Dependent Toxicity of CdTe/CdSe QDs
3.7. CdTe/CdSe QDs Induced Apoptosis



3.8. DNA Damage of L929 Mouse Fibroblasts Induced by QDs
| Group | Dose (μg/mL) | Tailing Rate (%) (n = 200) | Tailing Cell (n = 20) | ||
|---|---|---|---|---|---|
| DNA in Tail (%) | Tail Length | Olive Tail Moment | |||
| negative control | — | 11.34 ± 2.17 | 4.67 ± 4.23 | 32.20 ± 20.18 | 7.27 ± 6.23 |
| positive control | — | 88.25 ± 5.45 a | 24.83 ± 20.17 a | 207.45 ± 84.69 a | 47.56 ± 23.66 a |
| 2.2 nm CdSe | 3.5 | 14.67 ± 3.68 | 6.33 ± 5.02 | 37.64 ± 25.42 | 9.16 ± 7.30 |
| 7 | 17.24 ± 5.58 | 13.53 ± 10.46 | 89.27 ± 40.15 | 20.43 ± 18.33 | |
| 14 | 25.55 ± 5.84 a | 24.00 ± 19.68 a | 138.35 ± 72.64 a | 28.78 ± 20.73 a | |
| 2.2 nm CdTe | 3.5 | 13.22 ± 4.78 | 5.67 ± 4.64 | 38.86 ± 29.95 | 11.41 ± 6.27 |
| 7 | 18.85 ± 5.97 | 14.00 ± 12.94 a | 78.80 ± 45.85 a | 22.28 ± 18.69 | |
| 14 | 24.73 ± 4.57 a | 26.67 ± 20.74 a | 123.55 ± 65.73 a | 27.58 ± 16.62 a | |
| 3.5 nm CdTe | 3.5 | 13.35 ± 3.67 | 5.07 ± 4.48 | 38.37 ± 26.84 | 9.81 ± 6.16 |
| 7 | 14.49 ± 4.83 a | 12.67 ± 9.43 | 59.27 ± 30.15 | 18.62 ± 12.99 | |
| 14 | 19.25 ± 4.14 a,b | 23.33 ± 20.27 a,b | 94.68 ± 63.86 a,b | 20.11 ± 16.30 | |
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Hild, W.A.; Breunig, M.; Goepferich, A. Quantum dots—Nano-sized probes for the exploration of cellular and intracellular targeting. Eur. J. Pharm. Biopharm. 2008, 68, 153–168. [Google Scholar] [CrossRef] [PubMed]
- Howarth, M.; Liu, W.; Puthenveetil, S.; Zheng, Y.; Marshall, L.F.; Schmidt, M.M.; Wittrup, K.D.; Bawendi, M.G.; Ting, A.Y. Monovalent, reduced-size quantum dots for imaging receptors on living cells. Nat. Methods 2008, 5, 397–399. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.-L.; Zhang, Z.-L.; Sun, E.-Z.; Peng, J.; Xie, M.; Tian, Z.-Q.; Lin, Y.; Pang, D.-W. Visualizing the endocytic and exocytic processes of wheat germ agglutinin by quantum dot-based single-particle tracking. Biomaterials 2011, 32, 7616–7624. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Liu, S.; Gao, D.; Hu, D.; Gong, P.; Sheng, Z.; Deng, J.; Ma, Y.; Cai, L. Click-functionalized compact quantum dots protected by multidentate-imidazole ligands: Conjugation-ready nanotags for living-virus labeling and imaging. J. Am. Chem. Soc. 2012, 134, 8388–8391. [Google Scholar] [CrossRef] [PubMed]
- Shah, B.S.; Clark, P.A.; Moioli, E.K.; Stroscio, M.A.; Mao, J.J. Labeling of mesenchymal stem cells by bioconjugated quantum dots. Nano Lett. 2007, 7, 3071–3079. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Cui, Y.; Levenson, R.M.; Chung, L.W.K.; Nie, S. In vivo cancer targeting and imaging with semiconductor quantum dots. Nat. Biotechnol. 2004, 22, 969–976. [Google Scholar] [CrossRef] [PubMed]
- Gazouli, M.; Bouziotis, P.; Lyberopoulou, A.; Ikonomopoulos, J.; Papalois, A.; Anagnou, N.P.; Efstathopoulos, E.P. Quantum dots-bevacizumab complexes for in vivo imaging of tumors. In Vivo 2014, 28, 1091–1095. [Google Scholar] [PubMed]
- Bagalkot, V.; Zhang, L.; Levy-Nissenbaum, E.; Jon, S.; Kantoff, P.W.; Langer, R.; Farokhzad, O.C. Quantum dot-aptamer conjugates for synchronous cancer imaging, therapy, and sensing of drug delivery based on bi-fluorescence resonance energy transfer. Nano. Lett. 2007, 7, 3065–3070. [Google Scholar] [CrossRef] [PubMed]
- Kauffer, F.-A.; Merlin, C.; Balan, L.; Schneider, R. Incidence of the core composition on the stability, the ros production and the toxicity of cdse quantum dots. J. Hazard. Mater. 2014, 268, 246–255. [Google Scholar] [CrossRef] [PubMed]
- Katsumiti, A.; Gilliland, D.; Arostegui, I.; Cajaraville, M.P. Cytotoxicity and cellular mechanisms involved in the toxicity of cds quantum dots in hemocytes and gill cells of the mussel mytilus galloprovincialis. Aquat. Toxicol. 2014, 153, 39–52. [Google Scholar] [CrossRef] [PubMed]
- Zhu, M.; Nie, G.; Meng, H.; Xia, T.; Nel, A.; Zhao, Y. Physicochemical properties determine nanomaterial cellular uptake, transport and fate. Acc. Chem. Res. 2013, 46, 622–631. [Google Scholar] [CrossRef] [PubMed]
- Rivera-Gil, P.; Jimenez de Aberasturi, D.; Wulf, V.; Pelaz, B.; del Pino, P.; Zhao, Y.; de la Fuente, J.M.; Ruiz de Larramendi, I.; Rojo, T.; Liang, X.-J.; et al. The challenge to relate the physicochemical properties of colloidal nanoparticles to their cytotoxicity. Acc. Chem. Res. 2013, 46, 743–749. [Google Scholar] [CrossRef] [PubMed]
- Winnik, F.M.; Maysinger, D. Quantum dot cytotoxicity and ways to reduce it. Acc. Chem. Res. 2013, 46, 672–680. [Google Scholar] [CrossRef] [PubMed]
- Peng, L.; He, M.; Chen, B.; Wu, Q.; Zhang, Z.; Pang, D.; Zhu, Y.; Hu, B. Cellular uptake, elimination and toxicity of CdSe/ZnS quantum dots in HepG2 cells. Biomaterials 2013, 34, 9545–9558. [Google Scholar] [CrossRef] [PubMed]
- Derfus, A.M.; Chan, W.C.W.; Bhatia, S.N. Probing the cytotoxicity of semiconductor quantum dots. Nano Lett. 2004, 4, 11–18. [Google Scholar] [CrossRef]
- Bell, I.R.; Ives, J.A.; Jonas, W.B. Nonlinear effects of nanoparticles: Biological variability from hormetic doses, small particle sizes, and dynamic adaptive interactions. Dose Response 2014, 12, 202–232. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Chen, W.; Zhang, J.; Liu, J.; Chen, G.; Pope, C. In vitro and in vivo toxicity of cdte nanoparticles. J. Nanosci. Nanotechnol. 2007, 7, 497–503. [Google Scholar] [CrossRef] [PubMed]
- Lovrić, J.; Cho, S.J.; Winnik, F.M.; Maysinger, D. Unmodified cadmium telluride quantum dots induce reactive oxygen species formation leading to multiple organelle damage and cell death. Chem. Biol. 2005, 12, 1227–1234. [Google Scholar] [CrossRef] [PubMed]
- Lovrić, J.; Bazzi, H.; Cuie, Y.; Fortin, G.A.; Winnik, F.; Maysinger, D. Differences in subcellular distribution and toxicity of green and red emitting cdte quantum dots. J. Mol. Med. 2005, 83, 377–385. [Google Scholar] [CrossRef] [PubMed]
- Munari, M.; Sturve, J.; Frenzilli, G.; Sanders, M.B.; Brunelli, A.; Marcomini, A.; Nigro, M.; Lyons, B.P. Genotoxic effects of CdS quantum dots and Ag2s nanoparticles in fish cell lines (RTG-2). Mutat. Res./Genet. Toxicol. Environ. Mutagen. 2014, 775–776, 89–93. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Feng, D.; Shi, W.; Li, X.; Ma, H. Parallel comparative studies on the toxic effects of unmodified cdte quantum dots, gold nanoparticles, and carbon nanodots on live cells as well as green gram sprouts. Talanta 2013, 116, 237–244. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.W.; Zhang, Y.; Ge, C.W.; Jin, Y.L.; Hu, S.L.; Gu, N. Preparation and characterization of highly luminescent water-soluble cdte quantum dots as optical temperature probes. J. Nanosci. Nanotechnol. 2010, 10, 7578–7582. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.W.; Zhang, Y.; Ge, C.W.; Jin, Y.L.; Hu, S.L.; Gu, N. Temperature-dependent photoluminescence of highly luminescent water-soluble cdte quantum dots. Chin. Chem. Lett. 2009, 20, 977–980. [Google Scholar] [CrossRef]
- Wang, Y.; Niu, S.H.; Zhang, Z.J.; Wang, H.T.; Yuan, C.W.; Fu, D.G. Silica coating of water-soluble cdte/cds core-shell nanocrystals by microemulsion method. Chin. J. Chem. Phys. 2007, 20, 685–689. [Google Scholar] [CrossRef]
- Dobrovolskaia, M.A.; Clogston, J.D.; Neun, B.W.; Hall, J.B.; Patri, A.K.; McNeil, S.E. Method for analysis of nanoparticle hemolytic properties in vitro. Nano Lett. 2008, 8, 2180–2187. [Google Scholar] [CrossRef] [PubMed]
- Yeh, T.K.; Wu, J.P.; Chang, L.W.; Tsai, M.H.; Chang, W.H.; Tsai, H.T.; Yang, C.S.; Lin, P. Comparative tissue distributions of cadmium chloride and cadmium-based quantum dot 705 in mice: Safety implications and applications. Nanotoxicology 2011, 5, 91–97. [Google Scholar] [CrossRef] [PubMed]
- Lieberman, H.R. Estimating LD50 using the probit technique: A basic computer program. Drug Chem. Toxicol. 1983, 6, 111–116. [Google Scholar] [CrossRef] [PubMed]
- Yu, T.; Malugin, A.; Ghandehari, H. Impact of silica nanoparticle design on cellular toxicity and hemolytic activity. ACS Nano 2011, 5, 5717–5728. [Google Scholar] [CrossRef] [PubMed]
- Jumaa, M.; Furkert, F.H.; Müller, B.W. A new lipid emulsion formulation with high antimicrobial efficacy using chitosan. Eur. J. Pharm. Biopharm. 2002, 53, 115–123. [Google Scholar] [CrossRef]
- Lin, Y.-S.; Haynes, C.L. Impacts of mesoporous silica nanoparticle size, pore ordering, and pore integrity on hemolytic activity. J. Am. Chem. Soc. 2010, 132, 4834–4842. [Google Scholar] [CrossRef] [PubMed]
- Napierska, D.; Thomassen, L.C.J.; Rabolli, V.; Lison, D.; Gonzalez, L.; Kirsch-Volders, M.; Martens, J.A.; Hoet, P.H. Size-dependent cytotoxicity of monodisperse silica nanoparticles in human endothelial cells. Small 2009, 5, 846–853. [Google Scholar] [CrossRef] [PubMed]
- Liao, K.-H.; Lin, Y.-S.; Macosko, C.W.; Haynes, C.L. Cytotoxicity of graphene oxide and graphene in human erythrocytes and skin fibroblasts. ACS Appl. Mater. Interfaces 2011, 3, 2607–2615. [Google Scholar] [CrossRef] [PubMed]
- Mayer, A.; Vadon, M.; Rinner, B.; Novak, A.; Wintersteiger, R.; Fröhlich, E. The role of nanoparticle size in hemocompatibility. Toxicology 2009, 258, 139–147. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.-W.; Powers, K.; Baney, R. Physicochemical properties and blood compatibility of acylated chitosan nanoparticles. Carbohydr. Polymers 2004, 58, 371–377. [Google Scholar] [CrossRef]
- Duan, J.; Yu, Y.; Li, Y.; Yu, Y.; Li, Y.; Zhou, X.; Huang, P.; Sun, Z. Toxic effect of silica nanoparticles on endothelial cells through DNA damage response via chk1-dependent G2/M checkpoint. PLoS ONE 2013, 8. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Liu, C.; Yang, D.; Zhang, H.; Xi, Z. Comparative study of cytotoxicity, oxidative stress and genotoxicity induced by four typical nanomaterials: The role of particle size, shape and composition. J. Appl. Toxicol. 2009, 29, 69–78. [Google Scholar] [CrossRef] [PubMed]
- Fotakis, G.; Timbrell, J.A. In vitro cytotoxicity assays: Comparison of LDH, neutral red, MTT and protein assay in hepatoma cell lines following exposure to cadmium chloride. Toxicol. Lett. 2006, 160, 171–177. [Google Scholar] [CrossRef] [PubMed]
- Geiser, M.; Rothen-Rutishauser, B.; Kapp, N.; Schürch, S.; Kreyling, W.; Schulz, H.; Semmler, M.; Hof, V.I.; Heyder, J.; Gehr, P. Ultrafine particles cross cellular membranes by nonphagocytic mechanisms in lungs and in cultured cells. Environ. Health Perspect. 2005, 113, 1555–1560. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Xia, T.; Nel, A.E. The role of oxidative stress in ambient particulate matter-induced lung diseases and its implications in the toxicity of engineered nanoparticles. Free Radic. Biol. Med. 2008, 44, 1689–1699. [Google Scholar] [CrossRef] [PubMed]
- Xia, T.; Kovochich, M.; Liong, M.; Mädler, L.; Gilbert, B.; Shi, H.; Yeh, J.I.; Zink, J.I.; Nel, A.E. Comparison of the mechanism of toxicity of zinc oxide and cerium oxide nanoparticles based on dissolution and oxidative stress properties. ACS Nano 2008, 2, 2121–2134. [Google Scholar] [PubMed]
- Liu, X.; Tang, M.; Zhang, T.; Hu, Y.; Zhang, S.; Kong, L.; Xue, Y. Determination of a threshold dose to reduce or eliminate CdTe-induced toxicity in L929 cells by controlling the exposure dose. PLoS ONE 2013, 8. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Hu, Y.; Tang, M.; Kong, L.; Ying, J.; Wu, T.; Xue, Y.; Pu, Y. Liver toxicity of cadmium telluride quantum dots (CdTe QDs) due to oxidative stress in vitro and in vivo. Int. J. Mol. Sci. 2015, 16, 23279. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Pandey, A.K.; Singh, S.S.; Shanker, R.; Dhawan, A. Engineered ZnO and TiO2 nanoparticles induce oxidative stress and DNA damage leading to reduced viability of Escherichia coli. Free Radic. Biol. Med. 2011, 51, 1872–1881. [Google Scholar] [CrossRef] [PubMed]
- Setyawati, M.I.; Tay, C.Y.; Leong, D.T. Mechanistic investigation of the biological effects of SiO2, TiO2, and zno nanoparticles on intestinal cells. Small 2015, 11, 3458–3468. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Cui, H.; Zhou, J.; Li, F.; Wang, J.; Chen, M.; Liu, Q. Cytotoxicity, DNA damage, and apoptosis induced by titanium dioxide nanoparticles in human non-small cell lung cancer A549 cells. Environ. Sci. Pollut. Res. 2015, 22, 5519–5530. [Google Scholar] [CrossRef] [PubMed]
- Inayat-Hussain, S.H.; Chan, K.M.; Rajab, N.F.; Din, L.B.; Chow, S.C.; Kizilors, A.; Farzaneh, F.; Williams, G.T. Goniothalamin-induced oxidative stress, DNA damage and apoptosis via caspase-2 independent and Bcl-2 independent pathways in jurkat T-cells. Toxicol. Lett. 2010, 193, 108–114. [Google Scholar] [CrossRef] [PubMed]
- Verma, A.; Stellacci, F. Effect of surface properties on nanoparticle-cell interactions. Small 2010, 6, 12–21. [Google Scholar] [CrossRef] [PubMed]
- Beddoes, C.M.; Case, C.P.; Briscoe, W.H. Understanding nanoparticle cellular entry: A physicochemical perspective. Adv. Colloid Interface Sci. 2015, 218, 48–68. [Google Scholar] [CrossRef] [PubMed]
- Benameur, L.; Auffan, M.; Cassien, M.; Liu, W.; Culcasi, M.; Rahmouni, H.; Stocker, P.; Tassistro, V.; Bottero, J.-Y.; Rose, J.; et al. DNA damage and oxidative stress induced by CeO2 nanoparticles in human dermal fibroblasts: Evidence of a clastogenic effect as a mechanism of genotoxicity. Nanotoxicology 2014, 9, 696–705. [Google Scholar] [CrossRef] [PubMed]
- Siddiqui, M.A.; Saquib, Q.; Ahamed, M.; Farshori, N.N.; Ahmad, J.; Wahab, R.; Khan, S.T.; Alhadlaq, H.A.; Musarrat, J.; Al-Khedhairy, A.A.; et al. Molybdenum nanoparticles-induced cytotoxicity, oxidative stress, G2/M arrest, and DNA damage in mouse skin fibroblast cells (L929). Colloids Surf. B 2015, 125, 73–81. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Gao, X.; Su, X. Study the damage of DNA molecules induced by three kinds of aqueous nanoparticles. Talanta 2010, 80, 1228–1233. [Google Scholar] [CrossRef] [PubMed]
- Prasad, K.S.; Amin, T.; Katuva, S.; Kumari, M.; Selvaraj, K. Synthesis of water soluble CDS nanoparticles and study of their DNA damage activity. Arabian J. Chem. 2014. [Google Scholar] [CrossRef]
- Choi, Y.J.; Kim, Y.J.; Lee, J.W.; Lee, Y.; Lim, Y.B.; Chung, H.W. Cyto-/genotoxic effect of CdSe/ZnS quantum dots in human lung adenocarcinoma cells for potential photodynamic UV therapy applications. J. Nanosci. Nanotechnol. 2012, 12, 2160–2168. [Google Scholar] [CrossRef] [PubMed]
- Nagy, A.; Hollingsworth, J.A.; Hu, B.; Steinbrück, A.; Stark, P.C.; Rios Valdez, C.; Vuyisich, M.; Stewart, M.H.; Atha, D.H.; Nelson, B.C.; et al. Functionalization-dependent induction of cellular survival pathways by CdSe quantum dots in primary normal human bronchial epithelial cells. ACS Nano 2013, 7, 8397–8411. [Google Scholar] [CrossRef] [PubMed]
- Valko, M.; Izakovic, M.; Mazur, M.; Rhodes, C.J.; Telser, J. Role of oxygen radicals in DNA damage and cancer incidence. Mol. Cell. Biochem. 2004, 266, 37–56. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, T.; Wang, Y.; Kong, L.; Xue, Y.; Tang, M. Threshold Dose of Three Types of Quantum Dots (QDs) Induces Oxidative Stress Triggers DNA Damage and Apoptosis in Mouse Fibroblast L929 Cells. Int. J. Environ. Res. Public Health 2015, 12, 13435-13454. https://doi.org/10.3390/ijerph121013435
Zhang T, Wang Y, Kong L, Xue Y, Tang M. Threshold Dose of Three Types of Quantum Dots (QDs) Induces Oxidative Stress Triggers DNA Damage and Apoptosis in Mouse Fibroblast L929 Cells. International Journal of Environmental Research and Public Health. 2015; 12(10):13435-13454. https://doi.org/10.3390/ijerph121013435
Chicago/Turabian StyleZhang, Ting, Yiqing Wang, Lu Kong, Yuying Xue, and Meng Tang. 2015. "Threshold Dose of Three Types of Quantum Dots (QDs) Induces Oxidative Stress Triggers DNA Damage and Apoptosis in Mouse Fibroblast L929 Cells" International Journal of Environmental Research and Public Health 12, no. 10: 13435-13454. https://doi.org/10.3390/ijerph121013435