
Trichloroethylene and Its Oxidative Metabolites Enhance the Activated State and Th1 Cytokine Gene Expression in Jurkat Cells

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals, Antibodies and Supplies
2.2. Cell Culture
2.3. Assessment of Cell Viability
2.4. Measurement of IL-2 Production
2.5. Detection of Activation Markers
2.6. Real-Time PCR Analysis
2.7. Statistical Analysis
3. Results
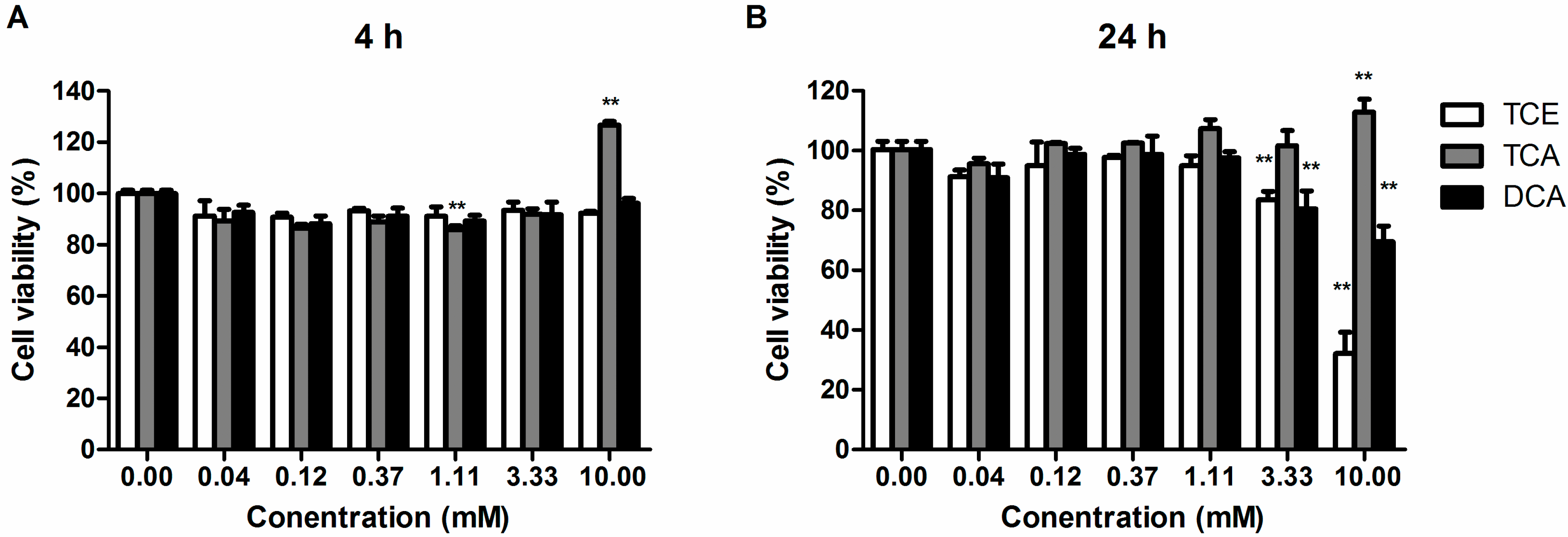
3.1. Effect of TCE, TCA and DCA on the Cell Viability of Jurkat Cells

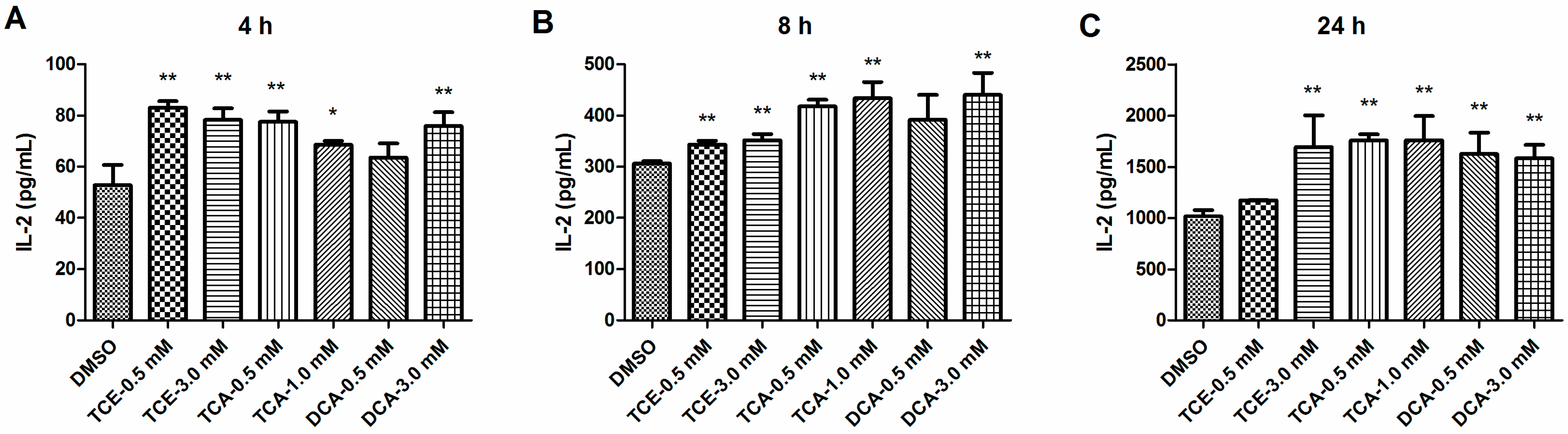
3.2. TCE, TCA and DCA Enhanced the Production of IL-2 in Jurkat Cells

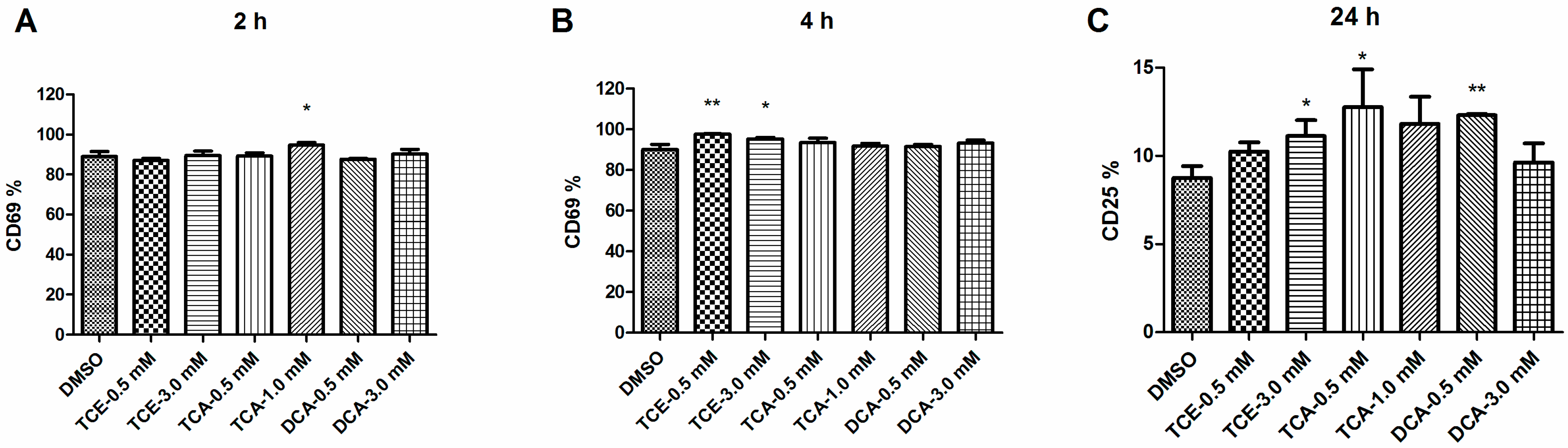
3.3. Influence of TCE, TCA and DCA on JurkatCells Activation Status
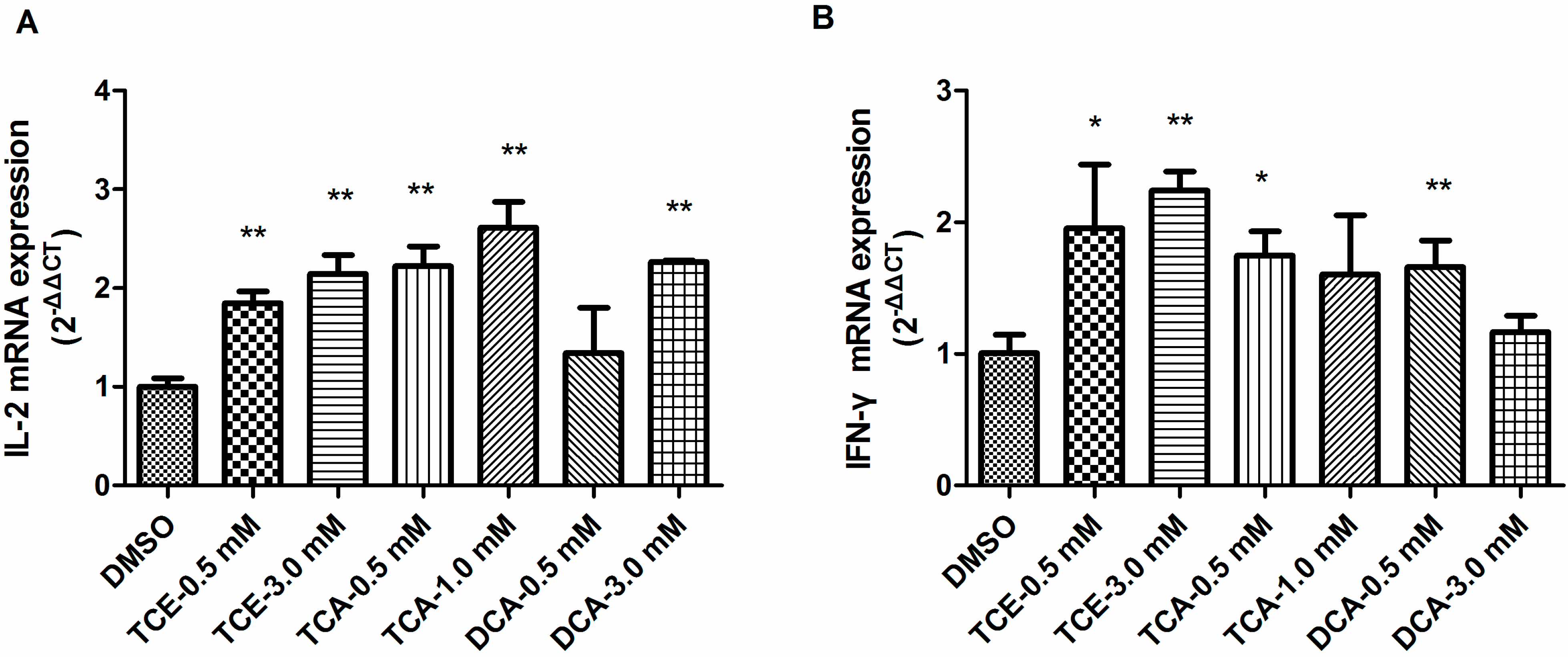
3.4. TCE, TCA and DCA Upregulated Th1 Cytokine Gene Expression in Jurkat Cells


4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Tang, X.J.; Li, L.Y. Guinea pig maximization test for trichloroethylene and its metabolites. Biomed. Environ. Sci. 2002, 15, 113–118. [Google Scholar] [PubMed]
- Tang, X.; Que, B. Characterization of liver injury associated with hypersensitive skin reactions induced by trichloroethylene in the guinea pig maximization test. J. Occup. Health 2008, 50, 114–121. [Google Scholar] [CrossRef] [PubMed]
- Lockey, J.E.; Kelly, C.R. Progressive systemic sclerosis associated with exposure to trichloroethylene. J. Occup. Med. 1987, 29, 493–496. [Google Scholar] [PubMed]
- Kilburn, K.H.; Warshaw, R.H. Prevalence of symptoms of systemic lupus erythematosus (SLE) and of fluorescent antinuclear antibodies associated with chronic exposure to trichloroethylene and other chemicals in well water. Environ. Res. 1992, 57, 1–9. [Google Scholar] [CrossRef]
- Goldman, J.A. Connective tissue disease in people exposed to organic chemical solvents: Systemic sclerosis (scleroderma) in dry cleaning plant and aircraft industry workers. J. Clin. Rheumatol. 1996, 2, 185–190. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, K.M.; Griffin, J.M. Trichloroethylene activates CD4+ T cells: Potential role in an autoimmune response. Drug Metab. Rev. 1999, 31, 901–916. [Google Scholar] [CrossRef] [PubMed]
- Kamijima, M.; Hisanaga, N. Occupational trichloroethylene exposure as a cause of idiosyncratic generalized skin disorders and accompanying hepatitis similar to drug hypersensitivities. Int. Arch. Occ. Env. Health 2007, 80, 357–370. [Google Scholar] [CrossRef] [PubMed]
- Kamijima, M.; Wang, H. Trichloroethylene causes generalized hypersensitivity skin disorders complicated by hepatitis. J. Occup. Health 2008, 50, 328–338. [Google Scholar] [CrossRef] [PubMed]
- Nakajima, T.; Yamanoshita, O. Generalized skin reactions in relation to trichloroethylene exposure: A review from the viewpoint of drug-metabolizing enzymes. J. Occup. Health 2003, 45, 8–14. [Google Scholar] [CrossRef] [PubMed]
- Guengerich, F.P.; Shimada, T. Oxidation of toxic and carcinogenic chemicals by human cytochrome P-450 enzymes. Chem. Res. Toxicol. 1991, 4, 391–407. [Google Scholar] [CrossRef] [PubMed]
- Lash, L.H.; Fisher, J.W. Metabolism of trichloroethylene. Environ. Health Perspect. 2000, 108, 177–200. [Google Scholar] [CrossRef] [PubMed]
- Blossom, S.J. Exposure to a metabolite of the environmental toxicant, trichloroethylene, attenuates CD4+ T cell activation-induced cell death by metalloproteinase-dependent fasl shedding. Toxicol. Sci. 2006, 92, 103–114. [Google Scholar] [CrossRef] [PubMed]
- Blossom, S.J.; Doss, J.C. Chronic exposure to a trichloroethylene metabolite in autoimmune-prone MRL+/+ mice promotes immune modulation and alopecia. Toxicol. Sci. 2006, 95, 401–411. [Google Scholar] [CrossRef] [PubMed]
- Jia, Q.; Zang, D. Cytokine expression in trichloroethylene-induced hypersensitivity dermatitis: An in vivo and in vitro study. Toxicol. Lett. 2012, 215, 31–39. [Google Scholar] [CrossRef] [PubMed]
- Gillis, S.; Watson, J. Biochemical and biological characterization of lymphocyte regulatory molecules. V. Identification of an interleukin 2-producing human leukemia T cell line. J. Exp. Med. 1980, 152, 1709–1719. [Google Scholar] [CrossRef] [PubMed]
- Manger, B.; Weiss, A. T cell activation: Differences in the signals required for IL 2 production by nonactivated and activated T cells. J. Immunol. 1985, 135, 3669–3673. [Google Scholar] [PubMed]
- Tanaka, S.; Akaishi, E. Zinc ions suppress mitogen-activated interleukin-2 production in Jurkat cells. Biochem. Bioph. Res. Commun. 2005, 335, 162–167. [Google Scholar] [CrossRef] [PubMed]
- Higai, K.; Tsukada, M. Prolonged high glucose suppresses phorbol 12-myristate 13-acetate and ionomycin-induced interleukin-2 mRNA expression in Jurkat cells. Biochim. Biophys. Acta 2009, 1790, 8–15. [Google Scholar] [CrossRef] [PubMed]
- Vazquez, B.N.; Laguna, T. CD69 gene is differentially regulated in T and B Cells by evolutionarily conserved promoter-distal elements. J. Immunol. 2009, 183, 6513–6521. [Google Scholar] [CrossRef] [PubMed]
- Létourneau, S.; Krieg, C. IL-2– and CD25-dependent immunoregulatory mechanisms in the homeostasis of T-cell subsets. J. Allergy Clin. Immun. 2009, 123, 758–762. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Huang, H. Research progress on immune injury resulted from occupational medicamentose-like dermatitis induced by trichloroethylene. China Occup. Med. 2010, 2, 157–162. [Google Scholar]
- Conde-Salazar, L.; Guimaraens, D. Subcorneal pustular eruption and erythema from occupational exposure to trichloroethylene. Contact Dermatitis 1983, 9, 235–237. [Google Scholar] [CrossRef] [PubMed]
- Nakayama, H.; Kobayashi, M. Generalized eruption with severe liver dysfunction associated with occupational exposure to trichloroethylene. Contact Dermatitis 1988, 19, 48–51. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, H.; Tohyama, M. Occupational trichloroethylene hypersensitivity syndrome with human herpesvirus-6 and cytomegalovirus reactivation. Dermatology 2010, 221, 17–22. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Xia, L. Trichloroethylene hypersensitivity syndrome is potentially mediated through its metabolite chloral hydrate. PLoS ONE 2015, 10, e127101. [Google Scholar] [CrossRef] [PubMed]
- Abraham, R.T.; Weiss, A. Jurkat T cells and development of the T-cell receptor signalling paradigm. Nat. Rev. Immunol. 2004, 4, 301–308. [Google Scholar] [CrossRef] [PubMed]
- Zagorski, J.W.; Turley, A.E.; Dover, H.E.; VanDenBerg, K.R.; Compton, J.R.; Rockwell, C.E. The Nrf2 activator, tBHQ, differentially affects early events following stimulation of Jurkat Cells. Toxicol. Sci. 2013, 136, 63–71. [Google Scholar] [CrossRef] [PubMed]
- Griffin, J.M.; Blossom, S.J. Trichloroethylene accelerates an autoimmune response by Th1 T cell activation in MRL+/+ mice. Immunopharmacology 2000, 46, 123–137. [Google Scholar] [CrossRef]
- Griffin, J.M.; Gilbert, K.M. Inhibition of CYP2E1 reverses CD4+ T-cell alterations in trichloroethylene-treated MRL+/+ mice. Toxicol. Sci. 2000, 54, 384–389. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, K.M.; Pumford, N.R. Environmental contaminant trichloroethylene promotes autoimmune disease and inhibits T-cell apoptosis in MRL. J. Immunotoxicol. 2006, 3, 263–267. [Google Scholar] [CrossRef] [PubMed]
- Blossom, S.; Doss, J. Developmental exposure to trichloroethylene promotes CD4+ T cell differentiation and hyperactivity in association with oxidative stress and neurobehavioral deficits in MRL+/+ mice. Toxicol. Appl. Pharm. 2008, 231, 344–353. [Google Scholar] [CrossRef] [PubMed]
- Blossom, S.J.; Pumford, N.R. Activation and attenuation of apoptosis of CD4+ T cells following in vivo exposure to two common environmental toxicants, trichloroacetaldehyde hydrate and trichloroacetic acid. J. Autoimmun. 2004, 23, 211–220. [Google Scholar] [CrossRef] [PubMed]
- Cai, P.; Boor, P.J. Immuno- and hepato-toxicity of dichloroacetic acid in MRL(+/+) and B6C3F1 mice. J. Immunotoxicol. 2007, 4, 107–115. [Google Scholar] [CrossRef] [PubMed]
- Iavicoli, I.; Marinaccio, A. Effects of occupational trichloroethylene exposure on cytokine levels in workers. J. Occup. Environ. Med. 2005, 47, 453–457. [Google Scholar] [CrossRef] [PubMed]
- Lehmann, I.; Thoelke, A. The influence of maternal exposure to volatile organic compounds on the cytokine secretion profile of neonatal T cells. Environ. Toxicol. 2002, 17, 203–210. [Google Scholar] [CrossRef] [PubMed]
- Lan, Q.; Zhang, L. Occupational exposure to trichloroethylene is associated with a decline in lymphocyte subsets and soluble CD27 and CD30 markers. Carcinogenesis 2010, 31, 1592–1596. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Bassig, B.A. Alterations in serum immunoglobulin levels in workers occupationally exposed to trichloroethylene. Carcinogenesis 2013, 34, 799–802. [Google Scholar] [CrossRef] [PubMed]
- Xia, L.H.; Huang, H.L. A clinical analysis of 50 cases of medicament-like dermatitis due to trichloroethylene. Zhonghua Lao Dong Wei Sheng Zhi Ye Bing Za Zhi 2004, 22, 207–210. (in Chiense). [Google Scholar] [PubMed]
- Muller, G.; Spassovski, M. Metabolism of trichloroethylene in man. II. Pharmacokinetics of metabolites. Arch. Toxicol. 1974, 32, 283–295. [Google Scholar] [CrossRef] [PubMed]
- Fisher, J.W.; Mahle, D. A human physiologically based pharmacokinetic model for trichloroethylene and its metabolites, trichloroacetic acid and free trichloroethanol. Toxicol. Appl. Pharmacol. 1998, 152, 339–359. [Google Scholar] [CrossRef] [PubMed]
- Merdink, J.L.; Robison, L.M. Kinetics of chloral hydrate and its metabolites in male human volunteers. Toxicology 2008, 245, 130–140. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pan, Y.; Wei, X.; Hao, W. Trichloroethylene and Its Oxidative Metabolites Enhance the Activated State and Th1 Cytokine Gene Expression in Jurkat Cells. Int. J. Environ. Res. Public Health 2015, 12, 10575-10586. https://doi.org/10.3390/ijerph120910575
Pan Y, Wei X, Hao W. Trichloroethylene and Its Oxidative Metabolites Enhance the Activated State and Th1 Cytokine Gene Expression in Jurkat Cells. International Journal of Environmental Research and Public Health. 2015; 12(9):10575-10586. https://doi.org/10.3390/ijerph120910575
Chicago/Turabian StylePan, Yao, Xuetao Wei, and Weidong Hao. 2015. "Trichloroethylene and Its Oxidative Metabolites Enhance the Activated State and Th1 Cytokine Gene Expression in Jurkat Cells" International Journal of Environmental Research and Public Health 12, no. 9: 10575-10586. https://doi.org/10.3390/ijerph120910575