
Protein Kinase CK2 Expression Predicts Relapse Survival in ERα Dependent Breast Cancer, and Modulates ERα Expression in Vitro


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Kaplan Meier Survival Analysis
2.2. Cell Culture
2.3. Luciferase Reporter Assay
2.4. Flow Cytometric Detection of ERα
3. Results and Discussion
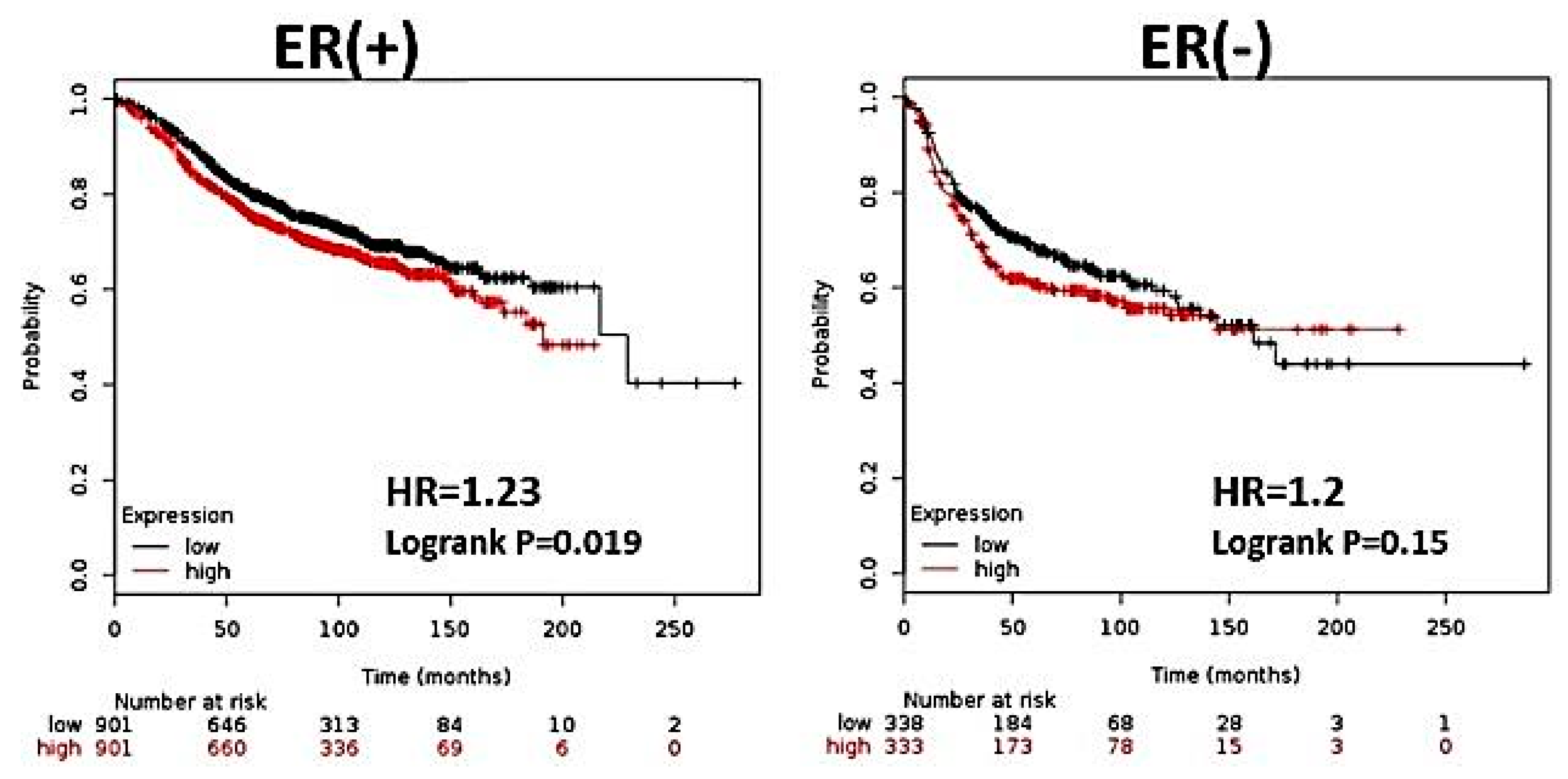
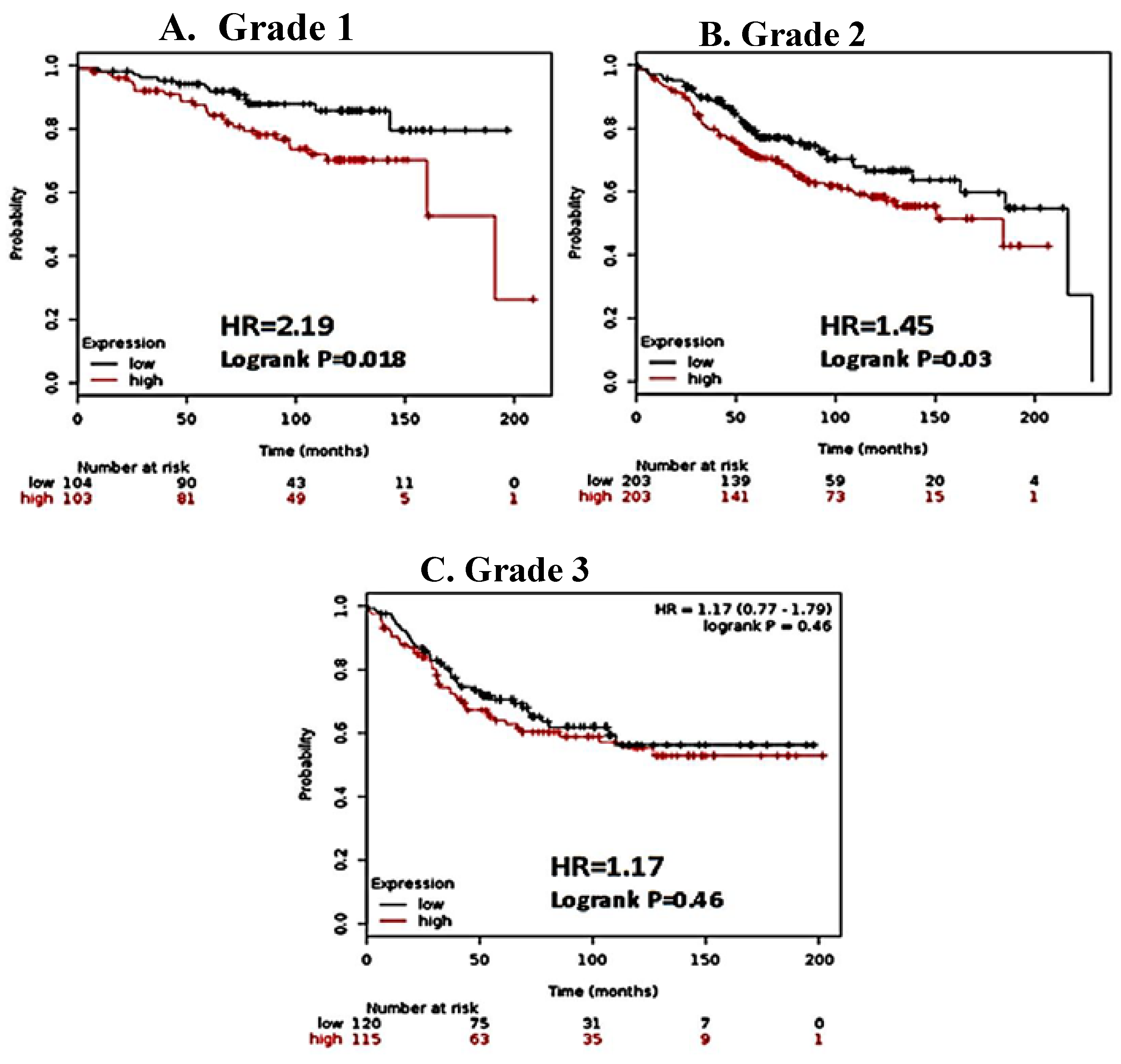
3.1. CK2α Overexpression is Associated with Decreased RFS in Patients with ERα (+) Breast Cancer

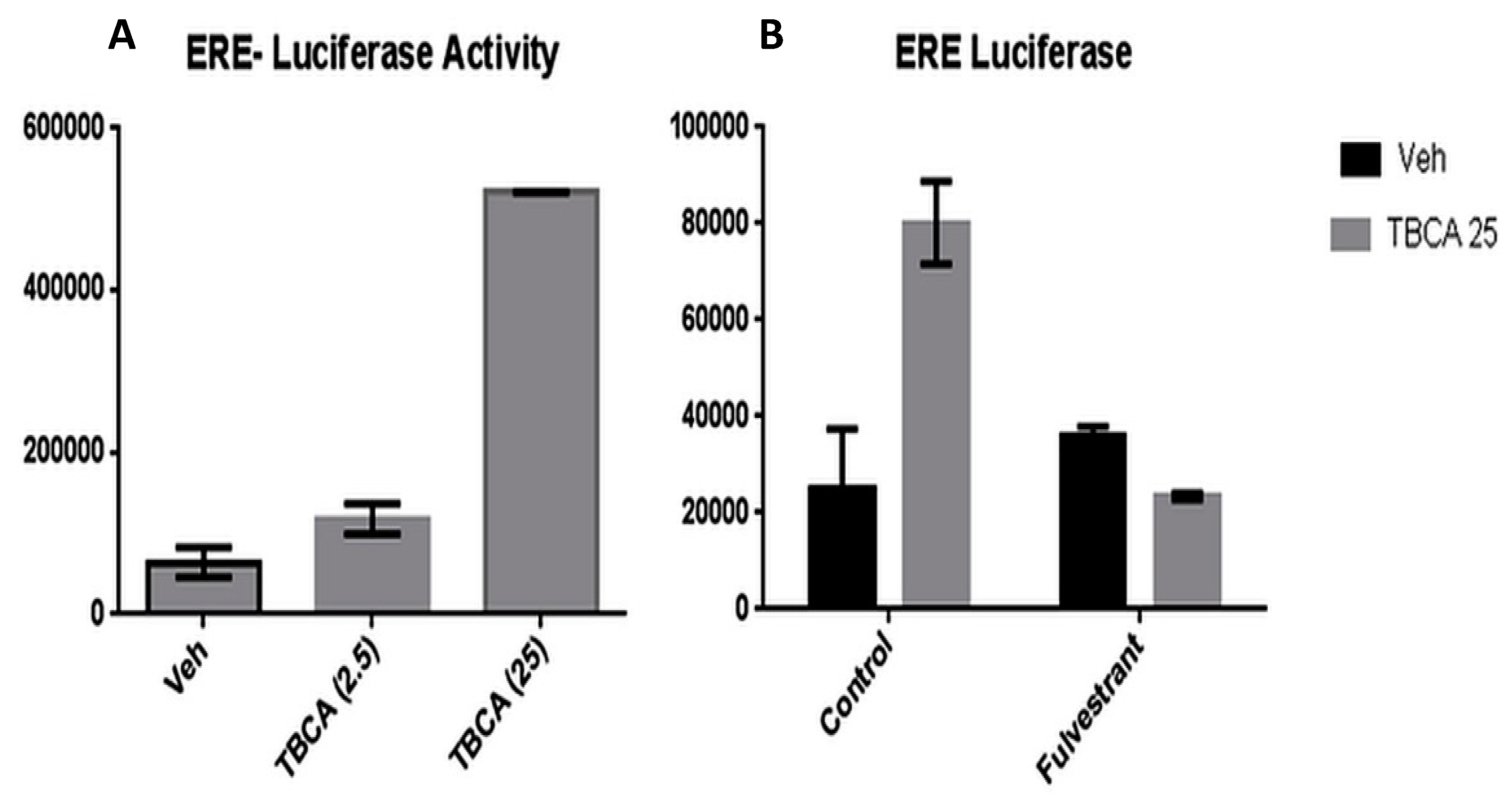
3.2. Inhibition of CK2α Results in Increased ER Transactivation in Stably Transfected T47D-luciferase Cells



3.3. TBCA Causes Increased Expression of ERα in ERα (+) Breast Cancer Cells, As Well As a Sub-Population of ERα (−) Breast Cancer Cells

4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Kurebayashi, J. Endocrine-resistant breast cancer: Underlying mechanisms and strategies for overcoming resistance. Breast Cancer 2003, 10, 112–119. [Google Scholar] [CrossRef] [PubMed]
- Issinger, O.G. Casein kinases: Pleiotropic mediators of cellular regulation. Pharmacol. Ther. 1993, 59, 1–30. [Google Scholar] [CrossRef]
- Allende, J.E.; Allende, C.C. Protein kinases. 4. Protein kinase CK2: An enzyme with multiple substrates and a puzzling regulation. FASEB J. 1995, 9, 313–323. [Google Scholar] [PubMed]
- Munstermann, U.; Fritz, G.; Seitz, G.; Lu, Y.P.; Schneider, H.R.; Issinger, O.G. Casein kinase II is elevated in solid human tumours and rapidly proliferating non-neoplastic tissue. Eur. J. Biochem. 1990, 189, 251–257. [Google Scholar] [CrossRef] [PubMed]
- Sarno, S.; Papinutto, E.; Franchin, C.; Bain, J.; Elliott, M.; Meggio, F.; Kazimierczuk, Z.; Orzeszko, A.; Zanotti, G.; Battistutta, R.; et al. ATP site-directed inhibitors of protein kinase CK2: An update. Curr. Top Med. Chem. 2011, 11, 1340–1351. [Google Scholar] [CrossRef] [PubMed]
- Sarduy, M.R.; García, I.; Coca, M.A.; Perera, A.; Torres, L.A.; Valenzuela, C.M.; Baladrón, I.; Solares, M.; Reyes, V.; Hernández, I.; et al. CERVIFARM-300-II Study Group Optimizing CIGB-300 intraregional delivery in locally advanced cervical cancer. Br. J. Cancer 2015, 112, 1636–1643. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Kim, S.H. Drug ability of the CK2 inhibitor CX-4945 as an anticancer drug and beyond. Arch. Pharm. Res. 2012, 35, 1293–1296. [Google Scholar] [CrossRef] [PubMed]
- Guerra, B.; Hochscherf, J.; Jensen, N.B.; Issinger, O.-G. Identification of a novel potent, selective and cell permeable inhibitor of protein kinase CK2 from the NIH/NCI Diversity Set Library. Mol. Cell. Biochem. 2015, 406, 151–161. [Google Scholar] [CrossRef] [PubMed]
- Alchab, F.; Ettouati, L.; Bouaziz, Z.; Bollacke, A.; Delcros, J.-G.; Gertzen, C.G.W.; Gohlke, H.; Pinaud, N.; Marchivie, M.; Guillon, J.; et al. Synthesis, Biological Evaluation and Molecular Modeling of Substituted Indeno [1,2-b]indoles as Inhibitors of Human Protein Kinase CK2. Pharmaceuticals 2015, 8, 279–302. [Google Scholar] [CrossRef] [PubMed]
- Ortega, C.E.; Seidner, Y.; Dominguez, I. Mining CK2 in cancer. PLoS ONE 2014, 9, e115609. [Google Scholar] [CrossRef] [PubMed]
- Kren, B.T.; Unger, G.M.; Abedin, M.J.; Vogel, R.I.; Henzler, C.M.; Ahmed, K.; Trembley, J.H. Preclinical evaluation of cyclin dependent kinase 11 and casein kinase 2 survival kinases as RNA interference targets for triple negative breast cancer therapy. Breast Cancer Res. 2015, 17, 19. [Google Scholar] [CrossRef] [PubMed]
- Landesman-Bollag, E.; Romieu-Mourez, R.; Song, D.H.; Sonenshein, G.E.; Cardiff, R.D.; Seldin, D.C. Protein kinase CK2 in mammary gland tumorigenesis. Oncogene 2001, 20, 3247–3257. [Google Scholar] [CrossRef] [PubMed]
- Yde, C.W.; Frogne, T.; Lykkesfeldt, A.E.; Fichtner, I.; Issinger, O.G.; Stenvang, J. Induction of cell death in antiestrogen resistant human breast cancer cells by the protein kinase CK2 inhibitor DMAT. Cancer Lett. 2007, 256, 229–237. [Google Scholar] [CrossRef] [PubMed]
- Williams, C.C.; Basu, A.; El-Gharbawy, A.; Carrier, L.M.; Smith, C.L.; Rowan, B.G. Identification of four novel phosphorylation sites in estrogen receptor alpha: Impact on receptor-dependent gene expression and phosphorylation by protein kinase CK2. BMC Biochem. 2009, 10, 36. [Google Scholar] [CrossRef] [PubMed]
- Györffy, B.; Lanczky, A.; Eklund, A.C.; Denkert, C.; Budczies, J.; Li, Q.; Szallasi, Z. An online survival analysis tool to rapidly assess the effect of 22,277 genes on breast cancer prognosis using microarray data of 1809 patients. Breast Cancer Res. Treat. 2010, 123, 725–731. [Google Scholar] [CrossRef] [PubMed]
- Gyorffy, B.; Lánczky, A.; Szállási, Z. Implementing an online tool for genome-wide validation of survival-associated biomarkers in ovarian-cancer using microarray data from 1287 patients. Endocr. Relat. Cancer 2012, 19, 197–208. [Google Scholar] [CrossRef] [PubMed]
- Tilghman, S.L.; Townley, I.; Zhong, Q.; Carriere, P.P.; Zou, J.; Llopis, S.D.; Preyan, L.C.; Williams, C.C.; Skripnikova, E.; Bratton, M.R.; et al. Proteomic signatures of acquired letrozole resistance in breast cancer: Suppressed estrogen signaling and increased cell motility and invasiveness. Mol. Cell Proteomics 2013. [Google Scholar] [CrossRef] [PubMed]
- Al-Dhaheri, M.; Rowan, B. Application of phosphorylation site-specific antibodies to measure nuclear receptor signaling: Characterization of novel phosphoantibodies for estrogen receptor A. Nuclear Recept. Signal. 2006. [Google Scholar] [CrossRef] [PubMed]
- Williams, C.C.; Allison, J.G.; Vidal, G.A.; Burow, M.E.; Beckman, B.S.; Marrero, L.; Jones, F.E. The ERBB4/HER4 receptor tyrosine kinase regulates gene expression by functioning as a STAT5A nuclear chaperone. J. Cell Biol. 2004, 167, 469–478. [Google Scholar] [CrossRef] [PubMed]
- Pagano, M.A.; Poletto, G.; di Maira, G.; Cozza, G.; Ruzzene, M.; Sarno, S.; Bain, J.; Elliott, M.; Moro, S.; Zagotto, G.; et al. Tetrabromocinnamic acid (TBCA) and related compounds represent a new class of specific protein kinase CK2 inhibitors. Chembiochem 2007, 8, 129–139. [Google Scholar] [CrossRef] [PubMed]
- Robertson, J.F. Faslodex (ICI 182, 780), a novel estrogen receptor downregulator—Future possibilities in breast cancer. J. Steroid. Biochem. Mol. Biol. 2001, 79, 209–212. [Google Scholar] [CrossRef]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Williams, M.D.; Nguyen, T.; Carriere, P.P.; Tilghman, S.L.; Williams, C. Protein Kinase CK2 Expression Predicts Relapse Survival in ERα Dependent Breast Cancer, and Modulates ERα Expression in Vitro. Int. J. Environ. Res. Public Health 2016, 13, 36. https://doi.org/10.3390/ijerph13010036
Williams MD, Nguyen T, Carriere PP, Tilghman SL, Williams C. Protein Kinase CK2 Expression Predicts Relapse Survival in ERα Dependent Breast Cancer, and Modulates ERα Expression in Vitro. International Journal of Environmental Research and Public Health. 2016; 13(1):36. https://doi.org/10.3390/ijerph13010036
Chicago/Turabian StyleWilliams, Marlon D., Thu Nguyen, Patrick P. Carriere, Syreeta L. Tilghman, and Christopher Williams. 2016. "Protein Kinase CK2 Expression Predicts Relapse Survival in ERα Dependent Breast Cancer, and Modulates ERα Expression in Vitro" International Journal of Environmental Research and Public Health 13, no. 1: 36. https://doi.org/10.3390/ijerph13010036
APA StyleWilliams, M. D., Nguyen, T., Carriere, P. P., Tilghman, S. L., & Williams, C. (2016). Protein Kinase CK2 Expression Predicts Relapse Survival in ERα Dependent Breast Cancer, and Modulates ERα Expression in Vitro. International Journal of Environmental Research and Public Health, 13(1), 36. https://doi.org/10.3390/ijerph13010036