
Current and Future Repellent Technologies: The Potential of Spatial Repellents and Their Place in Mosquito-Borne Disease Control


Abstract
:1. Introduction
2. A Brief History of Repellents
3. The Role of Repellents in Disease Prevention
4. Spatial vs. Contact Repellents: Similarities and Differences
5. Resistance to Repellent Chemistries
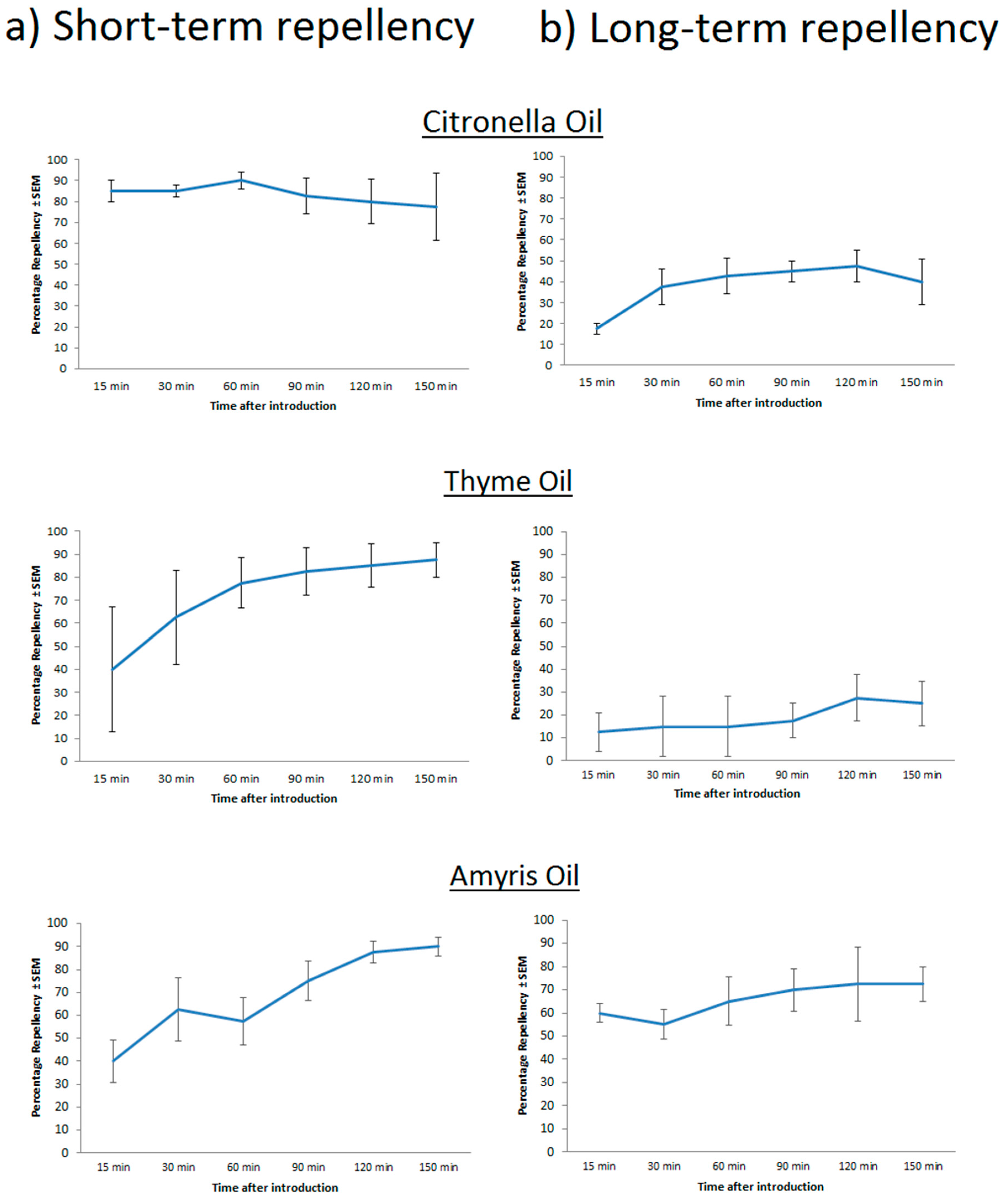
6. Current and Future Technologies
7. Conclusions
Author Contributions
Conflicts of Interest
References
- Service, M. Medical Entomology for Students; Service, M., Ed.; Cambridge University Press: Cambridge, UK, 2008. [Google Scholar]
- Norris, E.J.; Gross, A.D.; Dunphy, B.M.; Bessette, S.; Bartholomay, L.; Coats, J.R. Comparison of the insecticidal characteristics of commercially available plant essential oils against Aedes aegypti and Anopheles gambiae (Diptera: Culicidae). J. Med. Entomol. 2015, 52, 993–1002. [Google Scholar] [CrossRef] [PubMed]
- Tolle, M.A. Mosquito-borne diseases. Curr. Prob. Pediatr. Adolesc. Health Care 2009, 39, 97–140. [Google Scholar] [CrossRef] [PubMed]
- Mok, S.; Ashley, E.A.; Ferreira, P.E.; Zhu, L.; Lin, Z.; Yeo, T.; Chotivanich, K.; Imwong, M.; Pukrittayakamee, S.; Dhorda, M.; et al. Population transcriptomics of human malaria parasites reveals the mechanism of artemisinin resistance. Science 2014, 347, 431–435. [Google Scholar] [CrossRef] [PubMed]
- Mita, T.; Tanabe, K.; Kita, K. Spread and evolution of Plasmodium falciparum drug resistance. Parasitol. Int. 2009, 58, 201–209. [Google Scholar] [CrossRef] [PubMed]
- Laughlin, C.A.; Morens, D.M.; Cassetti, M.C.; Denis, A.C.; Martin, J.S.; Whitehead, S.S.; Fauci, A.S. Dengue research opportunities in the Americas. J. Inf. Dis. 2012, 206, 1121–1128. [Google Scholar] [CrossRef] [PubMed]
- Salazar-gonzález, J.A.; Angulo, C.; Rosales-mendoza, S. Chikungunya virus vaccines: Current strategies and prospects for developing plant-made vaccines. Vaccine 2015, 33, 3650–3658. [Google Scholar] [CrossRef] [PubMed]
- Peter, R.J.; Bossche, P.; Van Den Penzhorn, B.L.; Sharp, B. Tick, fly, and mosquito control—Lessons from the past, solutions for the future. Vet. Parasitol. 2005, 132, 205–215. [Google Scholar] [CrossRef] [PubMed]
- Revay, E.E.; Müller, G.C.; Qualls, W.A.; Kline, D.L.; Naranjo, D.P.; Arheart, K.L.; Kravchenko, V.D.; Yefremova, Z.; Hausmann, A.; Beier, J.C.; et al. Control of Aedes albopictus with attractive toxic sugar baits (ATSB) and potential impact on non-target organisms in St. Augustine, Florida. Parasitol. Rev. 2014, 113, 73–79. [Google Scholar] [CrossRef] [PubMed]
- Kline, D.L. Semiochemicals, traps/targets and mass trapping technology for mosquito management. J. Am. Mosq. Control. Assoc. 2007, 23, 241–251. [Google Scholar] [CrossRef]
- Caputo, B.; Ienco, A.; Cianci, D.; Pombi, M.; Petrarca, V.; Baseggio, A.; Devine, G.J.; della Torre, A. The “Auto-Dissemination” approach: A novel concept to fight Aedes albopictus in urban areas. PLoS Negl. Trop. Dis. 2012, 6, 4–11. [Google Scholar] [CrossRef] [PubMed]
- Menger, D.J.; Omusula, P.; Holdinga, M.; Homan, T. Field evaluation of a push-pull system to reduce malaria transmission. PLoS ONE 2015. [Google Scholar] [CrossRef] [PubMed]
- Atyame, C.M.; Labbé, P.; Lebon, C.; Tortosa, P. Comparison of irradiation and Wolbachia-based approaches for sterile-male strategies targeting Aedes albopictus. PLoS ONE 2016. [Google Scholar] [CrossRef] [PubMed]
- Winskill, P.; Carvalho, D.O.; Capurro, M.L.; Alphey, L. Dispersal of engineered male Aedes aegypti mosquitoes. PLoS Negl. Trop. Dis. 2015. [Google Scholar] [CrossRef] [PubMed]
- Pennisi, B.E.; Jasinskiene, N. Gene drive turns mosquitoes into malaria fighters. Science 2015, 350, 1014. [Google Scholar] [CrossRef] [PubMed]
- Achee, N.L.; Bangs, M.J.; Farlow, R.; Killeen, G.F.; Lindsay, S.; Logan, J.G.; Moore, S.J.; Rowland, M.; Sweeney, K.; Torr, S.J.; et al. Spatial repellents: From discovery and development to evidence-based validation. PLoS ONE 2012, 11, 164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paluch, G.; Bartholomay, L.; Coats, J. Mosquito repellents: A review of chemical structure diversity and olfaction. Pest Manag. Sci. 2010, 66, 925–935. [Google Scholar] [CrossRef] [PubMed]
- Bacot, A.; Talbot, G. The comparitive effectiveness of certain culicifuges under laboratory conditions. Parasitology 1919, 11, 221–236. [Google Scholar] [CrossRef]
- Bunker, C.; Hirschfelder, A. Mosquito repellents. Am. J. Trop. Med. Hygiene 1925, 5, 359–383. [Google Scholar]
- Dethier, V. Repellents. Annu. Rev. Entomol. 1956, 1, 181–202. [Google Scholar] [CrossRef]
- Morton, F.; Travis, B.; Linduska, J. Results of Screening Tests with Materials Evaluated as Insecticides, Miticides, and Repellents at the Orlando Laboratory. Available online: http://ufdc.ufl.edu/AA00025211/00001 (accessed on 26 January 2017).
- Travis, B. Mosquito repellents. Am. Perf. 1947, 50, 141–142. [Google Scholar]
- Travis, B.; Morton, F.; Jones, H.; Robinson, J. The more effective mosquito repellents tested at the Orlando, Fla., laboratory. J. Econ. Entomol. 1949, 42, 686–694. [Google Scholar] [CrossRef] [PubMed]
- Leal, W.S. The enigmatic reception of DEET—The gold standard of insect repellents. Curr. Opin. Insect Sci. 2014, 6, 93–98. [Google Scholar] [CrossRef] [PubMed]
- Carroll, S.P.; Loye, J. PMD, a registered botanical mosquito repellent with deet- like efficacy. J. Am. Mosq. Control Assoc. 2006, 22, 507–514. [Google Scholar] [CrossRef]
- Traboulsi, A.F.; El-Haj, S.; Tueni, M.; Taoubi, K.; Nader, N.A.; Mrad, A. Repellency and toxicity of aromatic plant extracts against the mosquito Culex pipiens molestus (Diptera: Culicidae). J. Med. Entomol. 2005, 604, 597–604. [Google Scholar]
- Zhu, J.; Xeng, Z.; Ma, Y.; Liu, T.; Han, Y.; Xue, S.; Tucker, B.; Schultz, G.; Coats, J.; Rowley, W.; Zhang, A. Adult repellency and larvicidal activity of five plant essential oils against mosquitoes. J. Am. Mosq. Control Assoc. 2006, 22, 515–522. [Google Scholar] [CrossRef]
- Vongsombath, C.; Pålsson, K.; Björk, L.; Karlson, B.; Jaenson, T.G.T. Mosquito (Diptera: Culicidae) repellency field tests of essential oils from plants traditionally used in Laos. J. Med. Entomol. 2012, 49, 1398–1404. [Google Scholar] [CrossRef] [PubMed]
- Müller, G.C.; Junnila, A.; Kravchenko, V.D.; Revay, E.E.; Butler, J.; Schlein, Y. Indoor protection against mosquito and sand fly bites: A comparison between citronella, linalool, and geraniol candles. J. Am. Mosq. Control Assoc. 2008, 24, 150–153. [Google Scholar]
- McDonald, G. The Epidemiology and Control of Malaria; Oxford Univeristy Press: Oxford, UK, 1957. [Google Scholar]
- Naowarat, S.; Thongjaem, P.; Tang, M. Effect of mosquito repellent on the transmission model of Chikungunya fever. Am. J. Appl. Sci. 2012, 9, 563–569. [Google Scholar]
- Philip, C.; Paul, J.; Sabin, A. Dimethyl phthalate as a repellent in control of phlebotomous (pappataci or sandfly) fever. War Med. 1944, 6, 27–33. [Google Scholar]
- Kroeger, A.; Gerhardus, A.; Kruger, G.; Mancheno, M.; Pesse, A. The contribution of repellent soap to malaria control. Am. J. Trop. Med. Hyg. 1997, 56, 580–584. [Google Scholar] [PubMed]
- Lutambi, A.; Penny, M.; Smith, T.; Chitnis, N. Mathematical modelling of mosquito dispersal in a heterogeneous environment. Math. Biosci. 2013, 241, 198–216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uzzan, B.; Konate, L.; Diop, A.; Nicolas, P.; Dia, I.; Dieng, Y. Efficacy of four insect repellents against mosquito bites: A double-blind randomized placebo-controlled field study in Senegal. Fundam. Clin. Pharmacol. 2009, 23, 589–594. [Google Scholar] [CrossRef] [PubMed]
- Rowland, M.; Downey, G.; Rab, A.; Freeman, T.; Mohammad, N.; Rehman, H.; Durrani, N.; Reybum, H.; Curtis, C.; et al. DEET mosquito repellent provides personal protection against malaria: A household randomized trial in an Afghan refugee camp in Pakistan. Trop. Med. Intern. Health 2004, 9, 335–342. [Google Scholar] [CrossRef]
- Sangoro, O.; Kelly, A.H.; Mtali, S.; Moore, S.J. Feasibility of repellent use in a context of increasing outdoor transmission: A qualitative study in rural Tanzania. Malar. J. 2014, 13, 347. [Google Scholar] [CrossRef] [PubMed]
- Sangoro, O.; Turner, E.; Simfukwe, E.; Miller, J.E.; Moore, S.J. A cluster-randomized controlled trial to assess the effectiveness of using 15% DEET topical repellent with long-lasting insecticidal nets (LLINs) compared to a placebo lotion on malaria transmission. Malar. J. 2014, 13, 324–339. [Google Scholar] [CrossRef] [PubMed]
- Lalani, T.; Yun, H.; Tribble, D.; Ganesan, A.; Kunz, A.; Fairchok, M.; Schnaubelt, E.; Fraser, J.; Mitra, I.; Kronmann, K.C.; et al. A comparison of compliance rates with anti-vectorial protective measures during travel to regions with dengue or chikungunya activity, and regions endemic for Plasmodium falciparum malaria. J. Travel Med. 2016, 23. [Google Scholar] [CrossRef] [PubMed]
- Barnard, D.R. Biological assay methods for mosquito repellents. J. Am. Mosq. Control Assoc. 2005, 21, 12–16. [Google Scholar] [CrossRef]
- Kawada, H.; Temu, E.A.; Minjas, J.N.; Matsumoto, O.; Iwasaki, T.; Takagi, M. Field evaluation of field repellency of metofluthrin impregnated latticework plastic strips against Aedes aegypti (L.) and analysis of environmental factors affecting its efficacy in My Tho City, Tien Giang, Vietnam. Am. J. Trop. Med. Hyg. 2006, 75, 1153–1157. [Google Scholar] [PubMed]
- SC Johnson Expands WOW Business Concept in Ghana. PR Newswire, 2014. Available online: http://www.ic.galegroup.com/ic/bic1/NewsDetailsPage/NewsDetailsWindow?disableHighlighting=&displayGroupName=News&currPage=&dviSelectedPage=&scanId=&query=&prodId=&search_within_results=&p=BIC1&mode=view&catId=&limiter=&display-query=&displayGroups=&contentModules=&action=e&sortBy=&documentId=GALE%7CA388599505&windowstate=normal&activityType=&failOverType=&commentary=&source=Bookmark&u=iastu_main&jsid=83e6733e1b20f489aed9d391225e32fc (accessed on 5 November 2016).
- Davis, E.E.; Program, N.; Park, M.; Sokolove, P.G. Lactic acid-sensitive receptors on the antennae of the mosquito, Aedes aegypti. J. Comp. Physiol. 1976, 54, 43–54. [Google Scholar] [CrossRef]
- Davis, E. Insect repellents: concepts of their mode of action relative to potential sensory mechanisms in mosquitoes (Diptera: Culicidae). J. Med. Entomol. 1985, 22, 237–243. [Google Scholar] [CrossRef] [PubMed]
- Boeckh, J.; Breer, H.; Geier, M.; Hoever, F.; Kruger, B.; Nentwig, G.; Sass, H. Acylated 1,3-Aminopropanols as repellents against bloodsucking arthropods. Pestic. Sci. 1996, 48, 359–373. [Google Scholar] [CrossRef]
- Dogan, E.; Ayres, J.; Rossignol, P. Behavioural mode of action of deet: Inhibition of lactic acid attraction. Med. Vet. Entomol. 1999, 13, 97–101. [Google Scholar] [CrossRef] [PubMed]
- Syed, Z.; Pelletier, J.; Flounders, E.; Chitolina, R.F.; Leal, W.S. Generic insect repellent detector from the fruit fly Drosophila melanogaster. PLoS ONE 2011, 6, e17705. [Google Scholar] [CrossRef] [PubMed]
- Ditzen, M.; Pellegrino, M.; Vosshall, L.B. Insect odorant receptors are molecular targets of the insect repellent DEET. Science 2008, 319, 1838–1842. [Google Scholar] [CrossRef] [PubMed]
- Xu, P.; Choo, Y.; Rosa, A.D.; Leal, W.S. Mosquito odorant receptor for DEET and methyl jasmonate. Proc. Natl. Acad. Sci. USA 2014, 111, 16592–16597. [Google Scholar] [CrossRef] [PubMed]
- Sathantriphop, S.; White, S.A.; Achee, N.L.; Sanguanpong, U.; Chareonviriyaphap, T. Behavioral responses of Aedes aegypti, Aedes albopictus, Culex quinquefasciatus, and Anopheles minimus against various synthetic and natural repellent compounds. J. Vector Ecol. 2014, 39, 328–339. [Google Scholar] [CrossRef] [PubMed]
- Licciardi, S.; Herve, J.; Darriet, F.; Hougard, J.; Corbel, V. Lethal and behavioural effects of three synthetic repellents (DEET, IR3535 and KBR 3023) on Aedes aegypti mosquitoes in laboratory assays. Med. Vet. Entomol. 2006, 20, 288–293. [Google Scholar] [CrossRef] [PubMed]
- Kweka, E.J.; Mosha, F.W.; Lowassa, A.; Mahande, A.M.; Mahande, M.J.; Massenga, C.P.; Tenu, F.; Lyatuu, E.E.; Mboya, M.A.; et al. Longitudinal evaluation of Ocimum and other plants effects on the feeding behavioral response of mosquitoes (Diptera: Culicidae) in the field in Tanzania. Parasit. Vectors 2008, 8, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Faulde, M.K.; Albiez, G.; Nehring, O. Insecticidal, acaricidal and repellent effects of DEET- and IR3535-impregnated bed nets using a novel long-lasting polymer-coating technique. Parasitol. Res. 2010, 106, 957–965. [Google Scholar] [CrossRef] [PubMed]
- Choi, D.B.; Grieco, J.P.; Apperson, C.S.; Schal, C. Effect of spatial repellent exposure on Dengue vector attraction to oviposition sites. PLoS Negl. Trop. Dis. 2011, 10, e0004850. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Lydy, M.J.; You, J. Pyrethroids in indoor air during application of various mosquito repellents: Occurrence, dissipation and potential exposure risk. Chemosphere 2016, 144, 2427–2435. [Google Scholar] [CrossRef] [PubMed]
- Ogoma, S.B.; Lorenz, L.M.; Ngonyani, H.; Sangusangu, R.; Kitumbukile, M.; Kilalangongono, M.; Simfukwe, E.T.; Mseka, A.; Mbeyela, E.; Roman, D.; et al. An experimental hut study to quantify the effect of DDT and airborne pyrethroids on entomological parameters of malaria transmission. Malar. J. 2014, 13, 1–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, Ke. Pyrethroids activate olfactory receptors and elicit spatial repellency in Drosophila melanogaster. In Proceedings of the XXV International Congress of Entomology, Orlando, FL, USA, 27 September 2016.
- Liu, F.; Chen, L.; Appel, A.G.; Liu, N. Olfactory responses of the antennal trichoid sensilla to chemical repellents in the mosquito, Culex quinquefasciatus. J. Insect Physiol. 2013, 59, 1169–1177. [Google Scholar] [CrossRef] [PubMed]
- Carey, A.F.; Wang, G.; Su, C.; Zwiebel, L.J.; Carlson, J.R. Odorant reception in the malaria mosquito Anopheles gambiae. Nature 2010, 464, 66–72. [Google Scholar] [CrossRef] [PubMed]
- Sathantriphop, S.; Achee, N.L.; Sanguanpong, U.; Chareonviriyaphap, T. The effects of plant essential oils on escape response and mortality rate of Aedes aegypti and Anopheles minimus. J. Vector Ecol. 2015, 40, 318–326. [Google Scholar] [CrossRef] [PubMed]
- Vinauger, C.; Lutz, E.K.; Riffell, J.A. Olfactory learning and memory in the disease vector mosquito Aedes aegypti. J. Exp. Biol. 2014, 217, 2321–2330. [Google Scholar] [CrossRef] [PubMed]
- Pellegrino, M.; Steinbach, N.; Stensmyer, M.C.; Hanson, B.S.; Vosshall, L.B. A natural polymorphism alters odour and DEET sensitivity in an insect odorant receptor. Nature 2011, 478, 511–516. [Google Scholar] [CrossRef] [PubMed]
- DeGennaro, M.; McBride, C.S.; Seeholzer, L.; Nakagawa, T.; Dennis, E.J.; Goldman, C.; Jasinskiene, N.; James, A.A.; Vosshall, L.B. orco mutant mosquitoes lose strong preference for humans and are not repelled by volatile DEET. Nature 2011, 498, 487–491. [Google Scholar] [CrossRef] [PubMed]
- Stanczyk, N.M.; Brookfield, J.F.Y.; Ignell, R.; Logan, J.G.; Field, L.M. Behavioral insensitivity to DEET in Aedes aegypti is a genetically determined trait residing in changes in sensillum function. Proc. Natl. Acad. Sci. USA 2010, 107, 8575–8580. [Google Scholar] [CrossRef] [PubMed]
- Klun, J.A.; Strickman, D.; Rowton, E.; Williams, J.; Kramer, M.; Roberts, D.; Debboun, M. Comparative resistance of Anopheles albimanus and Aedes aegypti to N,N-diethyl-3-methylbenzamide (Deet) and 2-methylpiperidinyl-3-cyclohexen-1-carboxamide (AI3-37220) in laboratory human-volunteer repellent assays. J. Med. Entomol. 2004, 41, 418–422. [Google Scholar] [CrossRef] [PubMed]
- Wagman, J.M.; Achee, N.L.; Grieco, J.P. Insensitivity to the spatial repellent action of transfluthrin in Aedes aegypti: A heritable trait associated with decreased insecticide susceptibility. PLoS Negl. Trop. Dis. 2015, 9, e0003726. [Google Scholar] [CrossRef] [PubMed]
- Agramonte, N.; Bloomquist, J.; Bernier, U. Pyrethroid resistance reduces the biting protection of treated clothing against Puerto Rican Aedes aegypti. In Proceedings of the XXV International Congress of Entomology, Orlando, FL, USA, 26 September 2016.
- Isman, M.B.; Miresmailli, S.; Machial, C. Commercial opportunities for pesticides based on plant essential oils in agriculture, industry and consumer products. Phytochem. Rev. 2011, 10, 197–204. [Google Scholar] [CrossRef]
- Bruyne, M.; De Foster, K.; Carlson, J.R.; Haven, N. Odor coding in the Drosophila antenna. Cell Press 2001, 30, 537–552. [Google Scholar] [CrossRef]
- Maia, M.F.; Moore, S.J. Plant-based insect repellents: A review of their efficacy, development and testing. Malar. J. 2011, 10, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katz, T.M.; Miller, J.H.; Hebert, A.A. Insect repellents: Historical perspectives and new developments. J. Am. Acad. Dermatol. 2008, 58, 865–871. [Google Scholar] [CrossRef] [PubMed]
- Harrewijn, P.; Minks, A.K.; Mollema, C. Evolution of plant volatile production in insect-plant relationships. Chemoecology 1995, 73, 55–73. [Google Scholar] [CrossRef]
- Misni, N.; Nor, Z.M.; Ahmad, R. New candidates for plant-based repellents against Aedes aegypti. J. Am. Mosq. Control. Assoc. 2016, 32, 117–123. [Google Scholar] [CrossRef] [PubMed]
- Paluch, G.; Grodnitzky, J.; Bartholomay, L.; Coats, J. Quantitative structure-activity relationships of botanical sesquiterpenes: Spatial and contact repellency to the yellow fever mosquito, Aedes aegypti. J. Agric. Food Chem. 2009, 57, 7618–7625. [Google Scholar] [CrossRef] [PubMed]
- Peterson, C.J.; Nemetz, L.T.; Jones, L.M.; Coats, J.R. Behavioral activity of catnip (Lamiaceae) essential oil components to the German cockroach (Blattodea: Blattellidae). J. Econ. Entomol. 2002, 95, 377–380. [Google Scholar] [CrossRef] [PubMed]
- Schultz, G.; Peterson, C.; Coats, J.R. Natural insect repellents: Activity against mosquitoes and cockroaches. In Natural Products for Pest Management; Rimando, A.M., Duke, S.O., Eds.; American Chemical Society: Washington, DC, USA, 2006; Volume 927, pp. 168–181. [Google Scholar]
- Sangwan, N.S.; Farooqi, A.H.A.; Shabih, F.; Sangwan, R.S. Regulation of essential oil production in plants. Plant Growth Regul. 1999, 34, 3–21. [Google Scholar] [CrossRef]
- Coats, J.R.; Norris, E.J.; Bartholomay, L.C.; Klimavicz, J. Biorepellents: Natural and biorational terpenes. In Proceedings of the XXV International Congress of Entomology, Orlando, FL, USA, 30 September 2016.



{kind=link}
{kind=link}
| Natural and Biorational Compounds | Minutes after Treating Filter Paper | |||||
|---|---|---|---|---|---|---|
| 15 | 90 | 150 | 315 | 390 | 450 | |
| monoterpenoid | 32.5 | 80 | 87.5 | 30 | 21.5 | 30 |
| derivative 1 | 10 | 60 | 62.5 | 30 | 30 | 40 |
| derivative 2 | 22.5 | 92.5 | 90 | 70 | 80 | 65 |
| derivative 3 | KD | KD | KD | 27.5 | 40 | 55 |
| derivative 4 | 25 | 90 | 92.5 | 27.5 | 17.5 | 20 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Norris, E.J.; Coats, J.R. Current and Future Repellent Technologies: The Potential of Spatial Repellents and Their Place in Mosquito-Borne Disease Control. Int. J. Environ. Res. Public Health 2017, 14, 124. https://doi.org/10.3390/ijerph14020124
Norris EJ, Coats JR. Current and Future Repellent Technologies: The Potential of Spatial Repellents and Their Place in Mosquito-Borne Disease Control. International Journal of Environmental Research and Public Health. 2017; 14(2):124. https://doi.org/10.3390/ijerph14020124
Chicago/Turabian StyleNorris, Edmund J., and Joel R. Coats. 2017. "Current and Future Repellent Technologies: The Potential of Spatial Repellents and Their Place in Mosquito-Borne Disease Control" International Journal of Environmental Research and Public Health 14, no. 2: 124. https://doi.org/10.3390/ijerph14020124
APA StyleNorris, E. J., & Coats, J. R. (2017). Current and Future Repellent Technologies: The Potential of Spatial Repellents and Their Place in Mosquito-Borne Disease Control. International Journal of Environmental Research and Public Health, 14(2), 124. https://doi.org/10.3390/ijerph14020124