
Requalification of a Brazilian Trichoderma Collection and Screening of Its Capability to Decolourise Real Textile Effluent


 ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Microorganisms and Culture Conditions
2.2. Morphological Fungal Identification
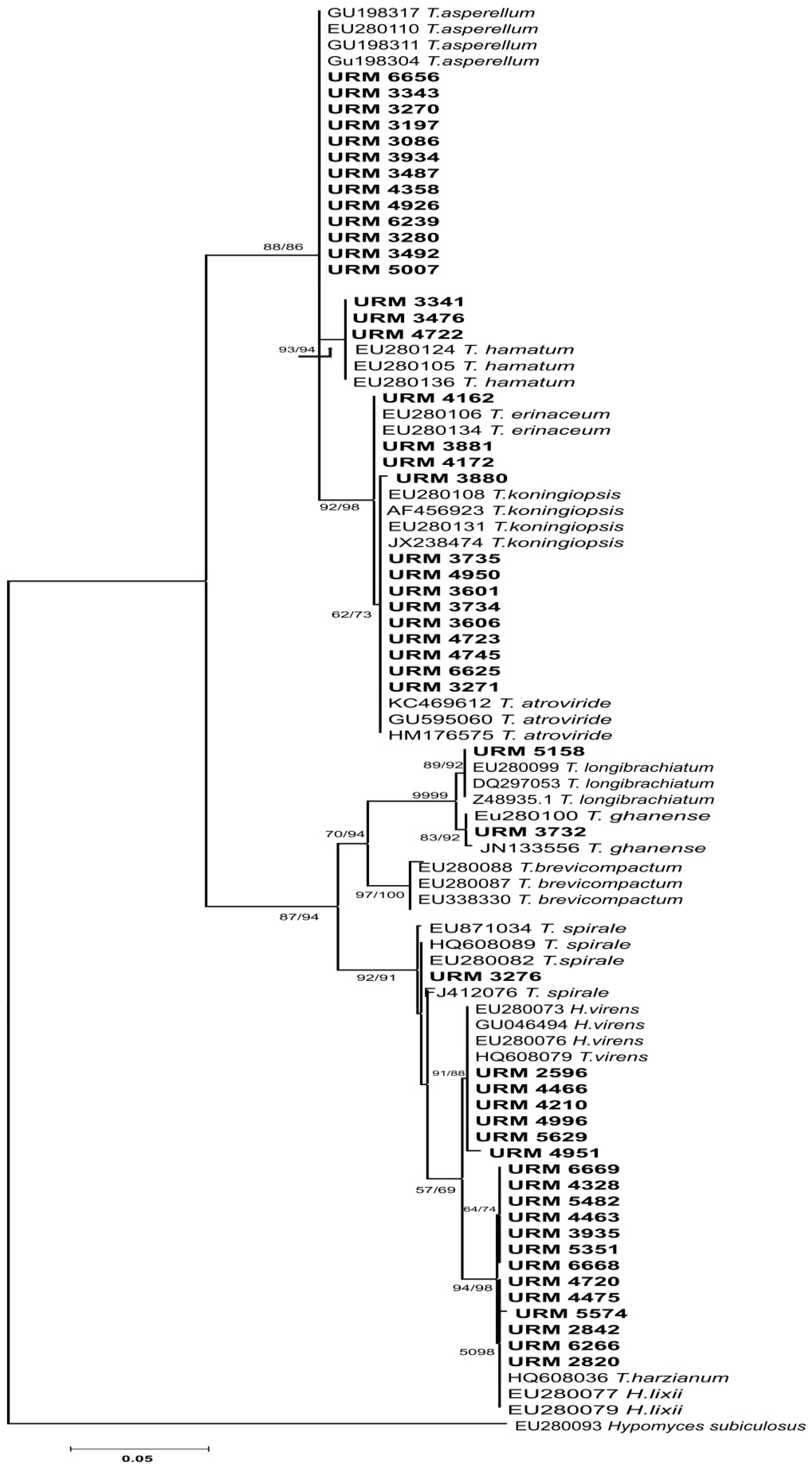
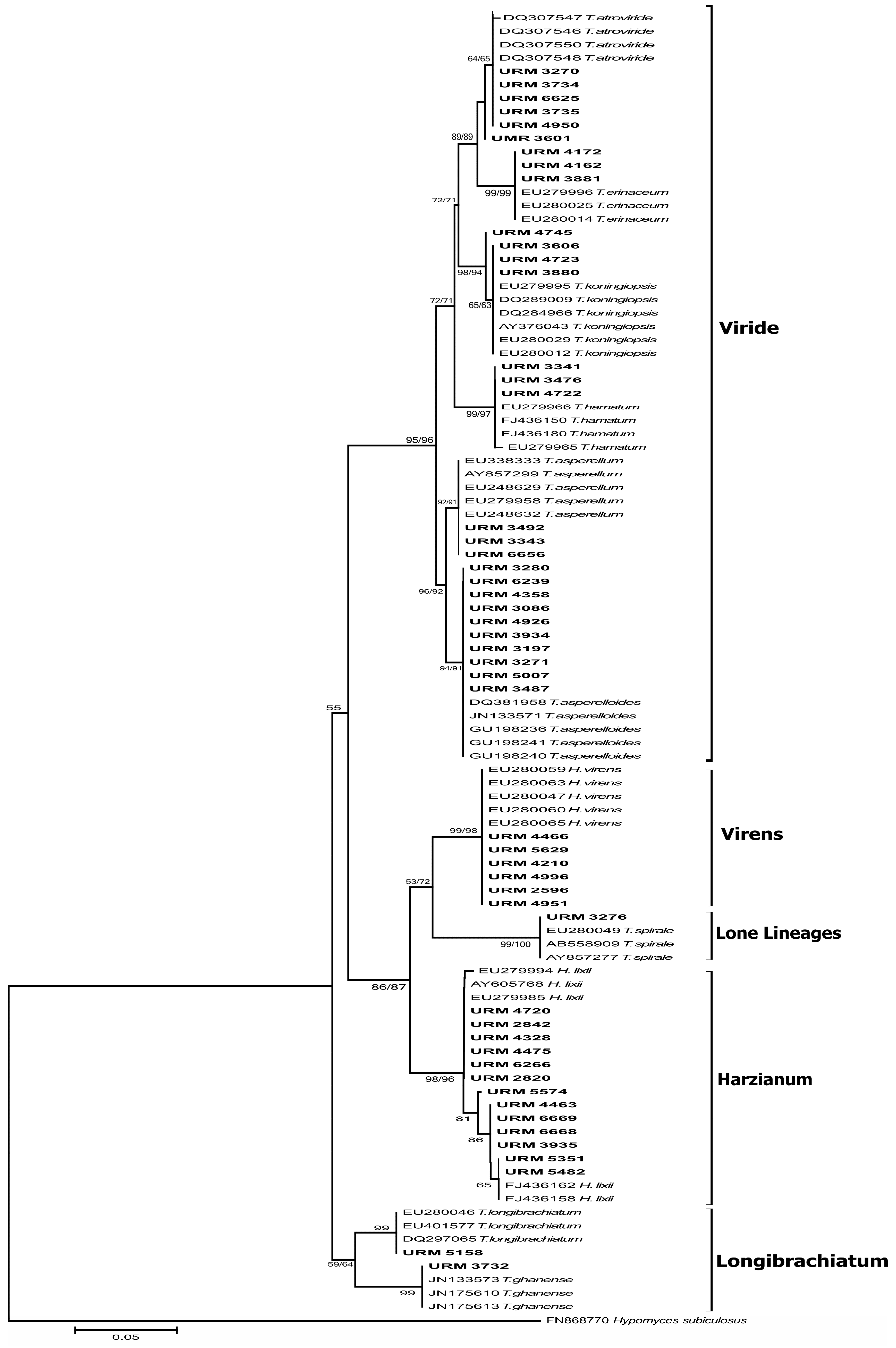
2.3. Molecular Fungal Identification
2.4. Decolourisation Assay
2.5. Determination of Enzymatic Activities
2.6. Phenotype Data Analysis
3. Results and Discussion
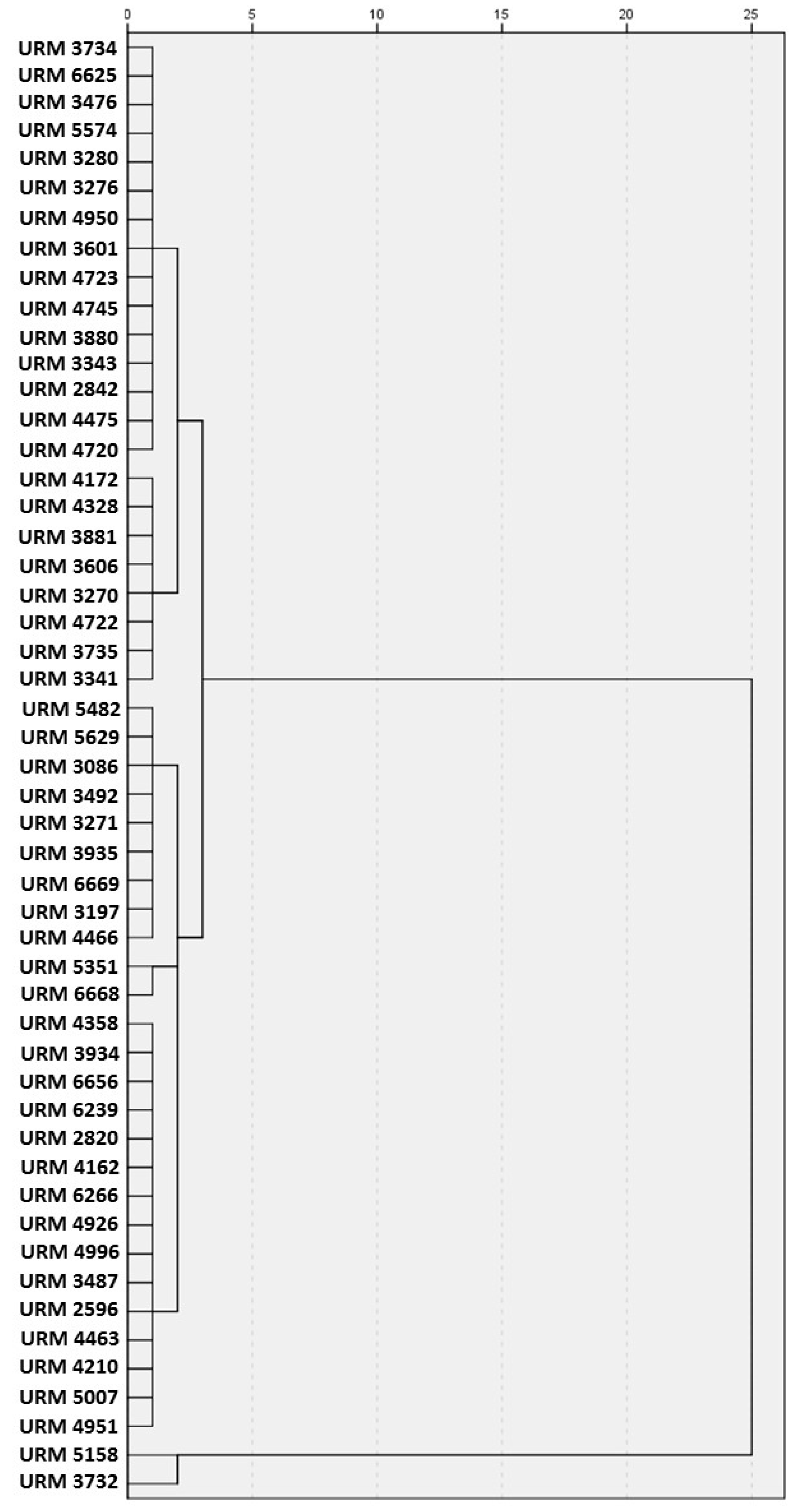
3.1. Morphological and Molecular Identification
3.2. Screening of Indigo Carmine Decolourisation in Malt Broth
3.3. Activity of Oxidase Enzymes
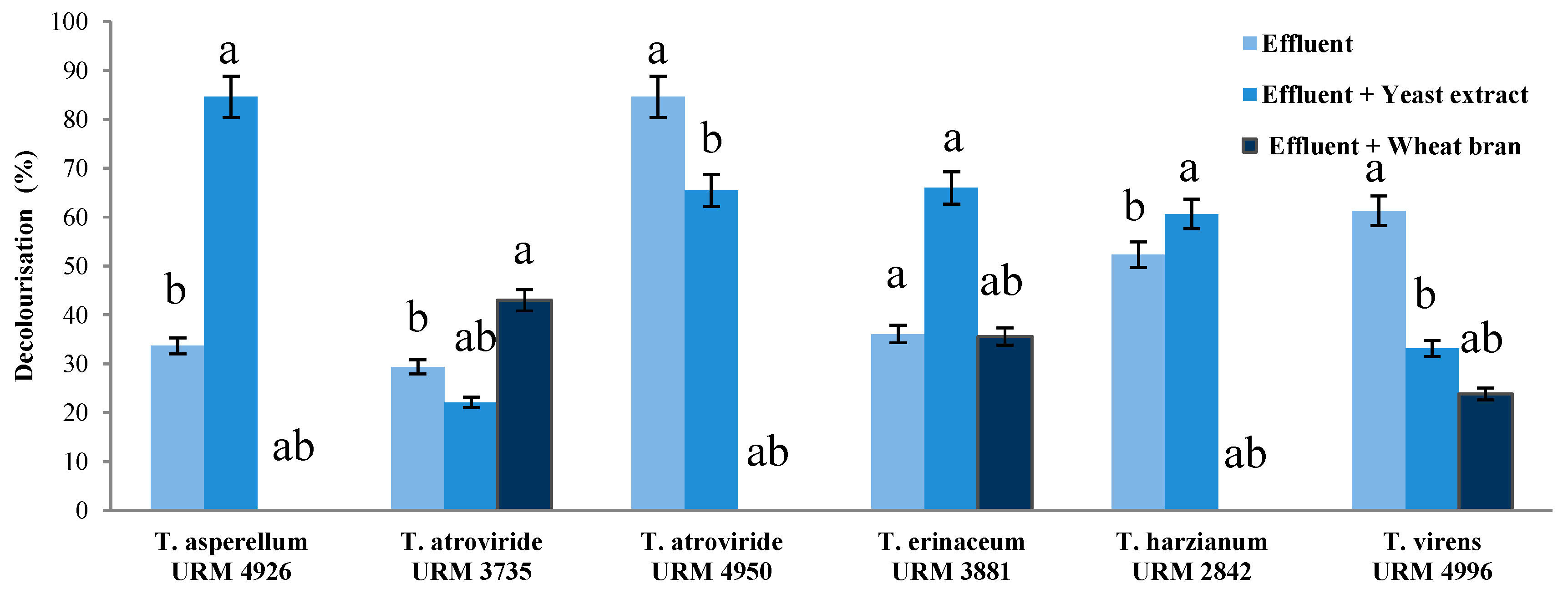
3.4. Treatment of Real Textile Effluent
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Durruty, I.; Fasce, I.; González, J.F.; Wolski, E.A. A kinetic study of textile dyeing wastewater degradation by Penicillium chrysogenum. Bioprocess Biosyst. Eng. 2015, 38, 1019–1031. [Google Scholar] [CrossRef] [PubMed]
- Ottoni, C.A.; Santos, C.; Kozakiewicz, Z.; Lima, N. White-rot fungi capable of decolourising textile dyes under alkaline conditions. Folia Microbiol. 2013, 58, 187–193. [Google Scholar] [CrossRef] [PubMed]
- Saroj, S.; Dubey, S.; Agarwal, P.; Prasad, R.; Singh, R.P. Evaluation of the efficacy of a fungal consortium for degradation of azo dyes and simulated textile dye effluents. Sustain. Water Resour. Manag. 2015, 1, 233–243. [Google Scholar] [CrossRef]
- Kanayama, N.; Suzuki, T.; Kawap, K. Purification and Characterization of an Alkaline Manganese Peroxidase from Aspergillus terreus LD-1. J. Biosci Bioeng. 2002, 93, 405–410. [Google Scholar] [CrossRef]
- Saparrat, M.C.M.; Martínez, M.J.; Cabello, M.N.; Arambarri, A.M. Screening for ligninolytic enzymes in autochthonous fungal strains from Argentina isolated from different substrata. Rev. Iberoam. Micol. 2002, 19, 181–185. [Google Scholar] [PubMed]
- Bonugli-Santos, R.C.; Durranta, L.R.; Silva, M.; Settec, L.D. Production of laccase, manganese peroxidase and lignin peroxidase by Brazilian marine-derived fungi. Enzyme Microb. Technol. 2010, 46, 32–37. [Google Scholar] [CrossRef]
- Maciel, M.; Ottoni, C.; Santos, C.; Lima, N.; Moreira, K.; Souza-Motta, C. Production of polygalacturonases by Aspergillus section Nigri strains in a fixed bed reactor. Molecules 2013, 18, 1660–1671. [Google Scholar] [CrossRef] [PubMed]
- Maciel, M.D.H.C.; Ottoni, C.A.; Herculano, P.N.; Porto, T.S.; Porto, A.L.F.; Santos, C.; Lima, N.; Moreira, K.A.; Souza-Motta, C.M. Purification of polygalacturonases produced by Aspergillus niger using an aqueous two-phase system. Fluid Phase Equilibr. 2014, 371, 125–130. [Google Scholar] [CrossRef]
- Ottoni, C.A.; Lima, L.; Santos, C.; Lima, N. Effect of Different Carbon Sources on Decolourisation of an Industrial Textile Dye Under Alkaline-Saline Conditions. Curr. Microbiol. 2014, 68, 53–58. [Google Scholar] [CrossRef] [PubMed]
- Passarini, M.R.Z.; Santos, C.; Lima, N.; Berlinck, R.G.S.; Sette, L.D. Filamentous fungi from the Atlantic marine sponge Dragmacidon reticulatum. Arch. Microbiol. 2013, 195, 99–111. [Google Scholar] [CrossRef] [PubMed]
- Sarayu, K.; Sandhya, S. Current technologies for biological treatment of textile wastewater: A review. Appl. Biochem. Biotechnol. 2012, 167, 645–661. [Google Scholar] [CrossRef] [PubMed]
- Ottoni, C.; Simões, M.F.; Fernandes, S.; Santos, C.R.; Lima, N. High laccase expression by Trametes versicolor in a simulated textile effluent with different carbon sources and pHs. Int. J. Environ. Res. Public Health 2016, 13, 778. [Google Scholar] [CrossRef] [PubMed]
- Zucca, P.; Neves, C.M.B.; Simões, M.M.Q.; Neves, M.G.P.M.S.; Cocco, G.; Sanjust, E. Immobilized lignin peroxidase-like metalloporphyrins as reusable catalysts in oxidative bleaching of industrial dyes. Molecules 2016, 21, 964. [Google Scholar] [CrossRef] [PubMed]
- Adnan, L.A.; Sathishkumar, P.; Yusoff, A.R.M.; Hadibarata, T. Metabolites characterization of laccase mediated Reactive Black 5 biodegradation by fast growing ascomycete fungus Trichoderma atroviride F03. Int. Biodeterior. Biodegradation 2015, 104, 274–282. [Google Scholar] [CrossRef]
- Hoyos-Carvajal, L.; Orduz, S.; Bissett, J. Genetic and metabolic diversity of Trichoderma from Colombia and adjacent neotropic regions. Fungal. Genet. Biol. 2009, 46, 615–631. [Google Scholar] [CrossRef] [PubMed]
- Nicolás, C.; Hermosa, R.; Rubio, B.; Mukherjee, P.K.; Monteb, E. Trichoderma genes in plants for stress tolerance-status and prospects. Plant. Sci. 2014, 228, 71–78. [Google Scholar] [CrossRef] [PubMed]
- Saravanakumar, K.; Kathiresan, K. Bioremoval of the synthetic dye malachite green by marine Trichoderma sp. SpringerPlus 2014, 3, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, P.; Singh, P.C.; Mishra, A.; Chauhan, P.S.; Dwivedi, S.; Bais, R.T.; Tripathi, R.D. Trichoderma: A potential bioremediator for environmental clean up. Clean Techn. Environ. Policy 2013, 15, 541–550. [Google Scholar] [CrossRef]
- Samuels, G.J.; Chaverri, P.; Farr, D.F.; McCray, E.B. Trichoderma Online, Systematic Mycology and Microbiology Laboratory, ARS, USDA. Available online: http://nt.ars-grin.gov/taxadescriptions/keys/TrichodermaIndex.cfm (accessed on 18 February 2016).
- Santos, C.; Ventura, J.A.; Costa, H.; Fernandes, P.M.B.; Lima, N. MALDI-TOF MS to identify the pineapple pathogen Fusarium guttiforme and its antagonist Trichoderma asperellum on decayed pineapple. Trop. Plant Pathol. 2015, 40, 227–232. [Google Scholar] [CrossRef]
- Griffiths, R.I.; Whiteley, A.S.; O’donnell, A.G.; Bailey, M.J. Rapid method for coextraction of DNA and RNA from natural environments for analysis of ribosomal DNA and RRNA-based microbial community composition. Appl. Environ. Microbiol. 2000, 66, 5488–5491. [Google Scholar] [CrossRef] [PubMed]
- Samuels, G.J.; Dodd, S.L.; Gams, W.; Castlebury, L.A.; Petrini, O. Trichoderma species associated with the green mold epidemic of commercially grown Agaricus bisporus. Mycologia 2002, 94, 146–170. [Google Scholar] [CrossRef] [PubMed]
- Carbone, I.; Kohn, L.M. A method for designing primer sets for speciation studies in filamentous ascomycetes. Mycologia 1999, 91, 553–556. [Google Scholar] [CrossRef]
- White, T.J. PCR Protocols, A Guide to Methods and Applications; Academic Press: New York, NY, USA, 1990; pp. 315–322. [Google Scholar]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular evolutionary genetics analysisusing maximum likelihood, evolutionary distance, and maximum parsimony method. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef] [PubMed]
- Rice, E.W.; Baird, R.B.; Eaton, A.D.; Clesceri, L.S. Standard Methods for the Examination of Water and Wastewater, 22nd ed.; American Public Health Association: Washington, DC, USA, 2012. [Google Scholar]
- Miranda, R.C.M.; Gomes, E.B.; Pereira, J.N.; Marin-Morales, M.A.; Machado, K.M.G.; Gusmão, N.B. Treatment of textile effluent under static bioreactor by Curvularia lunata URM 6179 and Phanerochaete chrysosporium URM 6181. Bioresour. Technol. 2013, 142, 361–367. [Google Scholar] [CrossRef] [PubMed]
- Silva, F.A.Z.; Azevedo, C.A.V. Principal Components Analysis in the Software Assistat-Statistical Attendance. In World Congress on Computers in Agriculture, 7; American Society of Agricultural and Biological Engineers: Reno, NV, USA, 2009. [Google Scholar]
- Arora, D.S.; Gill, P.K. Comparison of two assay procedures for lignin peroxidase. Enzyme Microb. Technol. 2001, 28, 602–605. [Google Scholar] [CrossRef]
- Kuwahara, M.; Glenn, J.K.; Morgan, M.A.; Gold, M.H. Separation and characterization of two extracellular H2O2-dependent oxidases from ligninolytic cultures of Phanerochaete chrysosporium. FEBS Lett. 1984, 169, 247–250. [Google Scholar] [CrossRef]
- Silva, F.C.; Chalfoun, S.M.; Batista, L.R.; Santos, C.; Lima, N. Use of a polyphasic approach including MALDI-TOF MS for identification of Aspergillus section Flavi strains isolated from food commodities in Brazil. Ann. Microbiol. 2015, 65, 2119–2129. [Google Scholar] [CrossRef]
- Rodrigues, P.; Santos, C.; Venâncio, A.; Lima, N. Species identification of Aspergillus section Flavi isolates from Portuguese almonds using phenotypic, including MALDI-TOF ICMS, and molecular approaches. J. Appl. Microbiol. 2011, 111, 877–892. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, M.M.E.; Santos, C.; Sampaio, P.; Romeo, O.; Almeida-Paes, R.; Pais, C.; Lima, N.; Zancope-Oliveira, R.M. Development and optimization of a new MALDI-TOF protocol for identification of the Sporothrix species complex. Res. Microbiol. 2015, 166, 102–110. [Google Scholar] [CrossRef] [PubMed]
- Gajera, H.P.; Bambharolia, R.P.; Hirpara, D.G.; Patel, S.V.; Golakiya, B.A. Molecular identification and characterization of novel Hypocrea koningii associated with azo dyes decolourisation and biodegradation of textile dye effluents. Process Saf. Environ. 2015, 98, 406–416. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| URM | Conidia | Phialide | Sterile Hyphae | Chlamyd. | PDA (mm) | SNA (mm) | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Shape 1 | Ornament 2 | Colour 3 | Length (μm) | Width (μm) | Length (μm) | Base Width (μm) | 30 °C | 35 °C | 40 °C | 35 °C | |||
| 3280 | GL/SGL | SW | G | 2.8–3.4 | 2.2–2.7 | 10.2–11.9 | 1.8–2.0 | − | + | 67.0 | 9.0 | Ø | 7.0 |
| 6239 | GL/SGL | SW | G | 2.8–3.4 | 3.2–3.6 | 8.5–10.2 | 1.8–2.0 | − | − | 77.0 | 24.0 | Ø | 16.0 |
| 3086 | GL/SGL | SW | G | 2.8–3.4 | 2.2–2.7 | 10.2–11.9 | 1.8–2.0 | − | + | 51.0 | 31.0 | Ø | 23.0 |
| 3487 | GL/SGL | SW | G | 2.8–3.4 | 2.2–2.7 | 10.2–11.9 | 1.8–2.0 | − | + | 74.0 | 28.0 | Ø | 24.0 |
| 3271 | GL/SGL | SW | G | 2.8–3.4 | 3.2–3.6 | 8.5–10.2 | 1.8–2.0 | − | − | 65.0 | 26.0 | Ø | 15.0 |
| 5007 | SGL/O | SW | G | 2.8–3.4 | 2.7–3.2 | 8.5–10.2 | 2.0–2.6 | − | − | 78.0 | 44.0 | Ø | 23.0 |
| 4358 | GL/SGL | SW | G | 3.5–4.1 | 2.2–2.7 | 10.2–11.9 | 1.8–2.0 | − | + | 76.0 | 25.0 | Ø | 20.0 |
| 3934 | GL/SGL | SW | G | 3.5–4.1 | 2.2–2.7 | 10.2–11.9 | 1.8–2.0 | − | + | 75.0 | 23.0 | Ø | 21.0 |
| 3343 | GL/SGL | SW | G | 2.8–3.4 | 2.2–2.7 | 8.5–10.2 | 2.0–2.6 | − | − | 73.0 | 14.0 | Ø | 6.0 |
| 6656 | SGL/O | SW | G | 2.8–3.4 | 2.2–2.7 | 8.5–10.2 | 2.0–2.6 | − | + | 77.0 | 22.0 | Ø | 18.0 |
| 4926 | GL/SGL | SW | G | 2.8–3.4 | 3.2–3.6 | 8.5–10.2 | 1.8–2.0 | + | + | 71.0 | 32.0 | Ø | 27.0 |
| 3197 | SGL/O | SW | G | 2.8–3.4 | 2.2–2.7 | 10.2–11.9 | 2.6–3.1 | − | + | 52.0 | 24.0 | Ø | 18.0 |
| 3735 | GL/SGL | S | G | 3.5–4.1 | 2.7–3.2 | 8.5–10.2 | 2.6–3.1 | − | − | 44.0 | Ø | Ø | Ø |
| 3734 | GL/SGL | S | G | 3.5–4.1 | 2.7–3.2 | 8.5–10.2 | 2.6–3.1 | − | − | 71.0 | Ø | Ø | Ø |
| 6625 | GL/SGL | S | G | 2.8–3.4 | 2.7–3.2 | 8.5–10.2 | 1.8–2.0 | − | − | 69.0 | Ø | Ø | Ø |
| 4950 | GL/SGL | S | G | 3.5–4.1 | 2.7–3.2 | 8.5–10.2 | 2.6–3.1 | − | − | 77.0 | Ø | Ø | Ø |
| 3601 | GL/SGL | S | G | 3.5–4.1 | 2.7–3.2 | 8.5–10.2 | 2.6–3.1 | − | − | 80.0 | Ø | Ø | Ø |
| 3270 | GL/SGL | S | G | 3.5–4.1 | 2.7–3.2 | 8.5–10.2 | 2.6–3.1 | − | − | 40.0 | Ø | Ø | Ø |
| 4162 | E | S | G | 4.2–4.8 | 2.7–3.2 | 6.8–8.5 | 1.8–2.0 | − | + | 73.0 | 20.0 | Ø | 17.0 |
| 4172 | E | S | G | 4.2–4.8 | 3.2–3.6 | 6.8–8.5 | 1.8–2.0 | − | + | 46.0 | 9.0 | Ø | 8.0 |
| 3881 | E | S | G | 4.2–4.8 | 3.2–3.6 | 6.8–8.5 | 1.8–2.0 | − | + | 50.0 | 14.0 | Ø | 11.0 |
| 3732 | E | S | G | 4.9–5.5 | 2.7–3.2 | 5.1–6.8 | 2.0–2.6 | − | − | 75.0 | 75.0 | 56.0 | 77.0 |
| 3341 | SGL/O | S | G | 3.5–4.1 | 2.7–3.2 | 10.2–11.9 | 2.0–2.6 | + | + | 54.0 | Ø | Ø | Ø |
| 3476 | SGL/O | S | G | 3.5–4.1 | 2.7–3.2 | 10.2–11.9 | 2.0–2.6 | + | + | 63.0 | Ø | Ø | Ø |
| 3492 | SGL/O | S | G | 2.8–3.4 | 2.7–3.2 | 10.2–11.9 | 1.8–2.0 | + | + | 64.0 | 25.0 | Ø | 9.0 |
| 4722 | SGL/O | S | G | 3.5–4.1 | 2.7–3.2 | 10.2–11.9 | 2.0–2.6 | + | + | 42.0 | Ø | Ø | Ø |
| 5351 | SGL/O | S | G | 2.4–2.7 | 1.8–2.2 | 8.5–10.2 | 1.8–2.0 | + | + | 75.0 | 35.0 | Ø | 36.0 |
| 6668 | SGL/O | S | G | 2.8–3.4 | 2.2–2.7 | 6.8–8.5 | 1.8–2.0 | − | + | 79.0 | 44.0 | Ø | 38.0 |
| 3935 | SGL/O | S | G | 2.4–2.7 | 1.8–2.2 | 10.2–11.9 | 2.0–2.6 | − | − | 58.0 | 20.0 | Ø | 14.0 |
| 5574 | SGL/O | S | G | 2.8–3.4 | 1.8–2.2 | 6.8–8.5 | 1.8–2.0 | + | − | 65.0 | 7.0 | Ø | 6.0 |
| 4463 | SGL/O | S | G | 2.8–3.4 | 1.8–2.2 | 8.5–10.2 | 1.8–2.0 | − | − | 78.0 | 29.0 | Ø | 29.0 |
| 2842 | SGL/O | S | G | 2.8–3.4 | 2.2–2.7 | 10.2–11.9 | 2.0–2.6 | − | − | 71.0 | 13.0 | Ø | 10.0 |
| 6669 | SGL/O | S | G | 2.4–2.7 | 1.8–2.2 | 10.2–11.9 | 2.0–2.6 | − | − | 58.0 | 17.0 | Ø | 12.0 |
| 4475 | SGL/O | S | G | 2.4–2.7 | 1.8–2.2 | 10.2–11.9 | 2.0–2.6 | − | − | 68.0 | 17.0 | Ø | 13.0 |
| 5482 | SGL/O | S | G | 2.4–2.7 | 1.8–2.2 | 10.2–11.9 | 2.0–2.6 | − | − | 57.0 | 31.0 | Ø | 28.0 |
| 4720 | GL/SGL | S | G | 2.4–2.7 | 2.2–2.7 | 8.5–10.2 | 2.0–2.6 | − | − | 64.0 | 15.0 | Ø | 11.0 |
| 6266 | SGL/O | S | G | 2.4–2.7 | 1.8–2.2 | 5.1–6.8 | 1.8–2.0 | − | + | 79.0 | 28.0 | Ø | 11.0 |
| 4328 | SGL/O | S | G | 2.4–2.7 | 1.8–2.2 | 10.2–11.9 | 2.0–2.6 | − | − | 50.0 | 8.0 | Ø | 8.0 |
| 2820 | SGL/O | S | G | 2.8–3.4 | 1.8–2.2 | 6.8–8.5 | 2.0–2.6 | − | − | 74.0 | 23.0 | Ø | 19.0 |
| 3606 | GL/SGL | S | G | 3.5–4.1 | 2.7–3.2 | 13.6–15.2 | 2.0–2.6 | − | − | 54.0 | 12.0 | Ø | 3.0 |
| 4723 | E | S | G | 3.5–4.1 | 1.8–2.2 | 6.8–8.5 | 1.8–2.0 | − | − | 76.0 | 8.0 | Ø | 15.0 |
| 4745 | E | S | G | 3.5–4.1 | 1.8–2.2 | 6.8–8.5 | 1.8–2.0 | − | − | 74.0 | 10.0 | Ø | 17.0 |
| 3880 | E | S | G | 3.5–4.1 | 1.8–2.2 | 6.8–8.5 | 1.8–2.0 | − | − | 72.0 | 11.0 | Ø | 13.0 |
| 5158 | E | S | G | 3.5–4.1 | 2.2–2.7 | 10.2–11.9 | 2.6–3.1 | − | + | 83.0 | 83.0 | 31.0 | 80.0 |
| 3276 | OB | S | G | 3.5–4.1 | 2.7–3.2 | 5.1–6.8 | 2.6–3.1 | + | + | 67.0 | 9.0 | Ø | Ø |
| 5629 | SGL/O | S | G | 3.5–4.1 | 2.2–2.7 | 13.6–15.2 | 1.8–2.0 | − | + | 56.0 | 28.0 | Ø | 27.0 |
| 4210 | SGL/O | S | G | 3.5–4.1 | 2.7–3.2 | 13.6–15.2 | 2.0–2.6 | − | − | 82.0 | 27.0 | Ø | 24.0 |
| 4466 | GL/SGL | S | G | 3.5–4.1 | 2.7–3.2 | 10.2–11.9 | 2.0–2.6 | − | + | 53.0 | 21.0 | Ø | 20.0 |
| 4951 | GL/SGL | S | G | 3.5–4.1 | 2.7–3.2 | 10.2–11.9 | 2.0–2.6 | + | + | 66.0 | 43.0 | Ø | 16.0 |
| 4996 | GL/SGL | S | G | 4.2–4.8 | 3.2–3.6 | 8.5–10.2 | 2.6–3.1 | − | + | 69.0 | 35.0 | Ø | 25.0 |
| 2596 | SGL/O | S | W/Y | 3.5–4.1 | 2.7–3.2 | 13.6–15.2 | 2.6–3.1 | − | + | 73.0 | 28.0 | Ø | 28.0 |
| URM | Revised Identification | Original Identification | Deposit Year | Substrate | Geographical Origin (Brazil) |
|---|---|---|---|---|---|
| 3280 | T. asperelloides | T. harzianum | 1992 | Soil | Paraná |
| 6239 | T. asperelloides | T. harzianum | 2010 | Soil | Pernambuco |
| 3086 | T. asperelloides | T. harzianum | 1989 | Leaf of R. gardenioides | São Paulo |
| 3487 | T. asperelloides | T. koningii | 1994 | Birds faeces | Pernambuco |
| 3271 | T. asperelloides | T. koningii | 1992 | Unknown | São Paulo |
| 5007 | T. asperelloides | T. virens | 2005 | Soil | Pernambuco |
| 4358 | T. asperelloides | T. virens | 2001 | Soil | Pernambuco |
| 3934 | T. asperelloides | T. virens | 1997 | Water | Pernambuco |
| 3343 | T. asperellum | T. aureoviride | 1993 | Unknown | Pernambuco |
| 6656 | T. asperellum | T. hamatum | 2012 | Soil with textile effluent | Pernambuco |
| 4926 | T. asperellum | T. harzianum | 2005 | Clay soil | Pernambuco |
| 3197 | T. asperellum | T. harzianum | 1990 | Amazonian nuts | Pará |
| 3735 | T. atroviride | T. aureoviride | 1997 | Lake water | Pernambuco |
| 3734 | T. atroviride | T. aureoviride | 1997 | Lake water | Pernambuco |
| 6625 | T. atroviride | T. aureoviride | 2012 | Soil of agroforestry | Pernambuco |
| 4950 | T. atroviride | T. harzianum | 2005 | Soil | Pernambuco |
| 3601 | T. atroviride | T. harzianum | 1995 | Unknown | Paraná |
| 3270 | T. atroviride | T. harzianum | 1992 | Unknown | São Paulo |
| 4162 | T. erinaceum | T. aureoviride | 1999 | Rhizosphere of sunflower | Pernambuco |
| 4172 | T. erinaceum | T. harzianum | 1999 | Rhizosphere of sunflower | Pernambuco |
| 3881 | T. erinaceum | T. koningii | 1997 | Soil of P. edulis culture | Pernambuco |
| 3732 | T. ghanense | T. koningii | 1997 | Lake water | Pernambuco |
| 3341 | T. hamatum | T. aureoviride | 1992 | Rhizosphere of V. herbacea | São Paulo |
| 3476 | T. hamatum | T. aureoviride | 1993 | Rhizosphere of V. herbacea | São Paulo |
| 3492 | T. hamatum | T. hamatum | 1994 | Birds faeces | Pernambuco |
| 4722 | T. hamatum | T. harzianum | 2003 | Garden of A. cephalotes | Alagoas |
| 5351 | T. harzianum | T. aureoviride | 2006 | Sugarcane | Unknown |
| 6668 | T. harzianum | T. aureoviride | 2012 | Soil with textile effluent | Pernambuco |
| 3935 | T. harzianum | T. aureoviride | 1997 | River water | Pernambuco |
| 5574 | T. harzianum | T. aureoviride | 2007 | Mangrove sediment | Pernambuco |
| 4463 | T. harzianum | T. aureoviride | 2002 | Sea water | Pernambuco |
| 2842 | T. harzianum | T. aureoviride | 1986 | Unknown | Pernambuco |
| 6669 | T. harzianum | T. aureoviride | 2012 | Soil with textile effluent | Pernambuco |
| 4475 | T. harzianum | T. harzianum | 2002 | Beach sand | Pernambuco |
| 5482 | T. harzianum | T. harzianum | 2007 | Unknown | São Paulo |
| 4720 | T. harzianum | T. harzianum | 2003 | Garden of A. cephalotes | Alagoas |
| 6266 | T. harzianum | T. harzianum | 2010 | Soil of agroforestry | Pernambuco |
| 4328 | T. harzianum | T. harzianum | 2001 | Soil of mining | Bahia |
| 2820 | T. harzianum | T. viride | 1985 | Sugarcane bagasse | Alagoas |
| 3606 | T. koningiopsis | T. hamatum | 1995 | Unknown | Paraná |
| 4723 | T. koningiopsis | T. koningii | 2003 | Garden of A. cephalotes | Alagoas |
| 4745 | T. koningiopsis | T. koningii | 2003 | Garden of A. cephalotes | Alagoas |
| 3880 | T. koningiopsis | T. koningii | 1997 | Soil of P. edulis culture | Pernambuco |
| 5158 | T. longibrachiatum | T. aureoviride | 2005 | Cement | Pernambuco |
| 3276 | T. spirale | T. hamatum | 1991 | sorghum | Pernambuco |
| 5629 | T. virens | T. hamatum | ---- | Unknown | Paraná |
| 4210 | T. virens | T. virens | 1999 | Rhizosphere of sunflower | Pernambuco |
| 4466 | T. virens | T. virens | 2002 | Sea water | Pernambuco |
| 4951 | T. virens | T. virens | 2005 | Soil | Pernambuco |
| 4996 | T. virens | T. virens | 2005 | Soil | Pernambuco |
| 2596 | T. virens | T. viride | 1980 | Unknown | Pernambuco |
| Species | URM | Lac | LiP | MnP |
|---|---|---|---|---|
| T. asperelloides | 3280 | 1.7 aC | 760.0 cdA | 379.3 defB |
| T. asperellum | 4926 | 8.3 aC | 517.3 fghA | 366.0 defgB |
| T. atroviride | 3270 | 2.0 aC | 758.0 cdA | 394.0 defB |
| T. atroviride | 3735 | 8.3 aC | 1307.0 aA | 617.3 bcB |
| T. atroviride | 4950 | 2.3 aB | 750.0 cdA | 680.0 bA |
| T. atroviride | 6625 | 1.3 aB | 692.0 deA | 90.7 jlB |
| T. erinaceum | 3881 | 10.0 aC | 378.7 iB | 875.3 aA |
| T. ghanense | 3732 | 1.7 aC | 766.3 cdA | 298.0 fghB |
| T. hamatum | 3476 | 1.0 aC | 593.3 efgA | 92.7 jlB |
| T. harzianum | 2842 | 2.3 aC | 924.0 bA | 176.0 hijB |
| T. harzianum | 3935 | 1.3 aC | 480.0 ghiA | 143.3 ijB |
| T. harzianum | 4328 | 2.7 aB | 378.0 iA | 442.7 deA |
| T. harzianum | 4463 | 1.7 aC | 758.7 cdA | 232.0 ghiB |
| T. harzianum | 5482 | 2.7 aC | 631.3 defA | 369.3 defB |
| T. harzianum | 6668 | 3.0 aB | 440.0 hiA | 0.0 lB |
| T. koningiopsis | 3880 | 7.7 aC | 645.3 defA | 146.0 ijB |
| T. virens | 4996 | 15.7 aC | 844.0 bcA | 329.3 efgB |
| T. virens | 5629 | 1.7 aC | 612.7 efgA | 500.0 cdB |
| COD (mg O2·L−1) | BOD (mg O2·L−1) | Colour (HAZEN) | Turbidity (NTU) | pH | SD (mL·L−1) |
|---|---|---|---|---|---|
| 3192.5 | 54.1 | 448.0 | 0.19 | 5.11 | 7.0 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Silva Lisboa, D.; Santos, C.; Barbosa, R.N.; Magalhães, O.; Paiva, L.M.; Moreira, K.A.; Lima, N.; Souza-Motta, C.M. Requalification of a Brazilian Trichoderma Collection and Screening of Its Capability to Decolourise Real Textile Effluent. Int. J. Environ. Res. Public Health 2017, 14, 373. https://doi.org/10.3390/ijerph14040373
Silva Lisboa D, Santos C, Barbosa RN, Magalhães O, Paiva LM, Moreira KA, Lima N, Souza-Motta CM. Requalification of a Brazilian Trichoderma Collection and Screening of Its Capability to Decolourise Real Textile Effluent. International Journal of Environmental Research and Public Health. 2017; 14(4):373. https://doi.org/10.3390/ijerph14040373
Chicago/Turabian StyleSilva Lisboa, Dianny, Cledir Santos, Renan N. Barbosa, Oliane Magalhães, Laura M. Paiva, Keila A. Moreira, Nelson Lima, and Cristina M. Souza-Motta. 2017. "Requalification of a Brazilian Trichoderma Collection and Screening of Its Capability to Decolourise Real Textile Effluent" International Journal of Environmental Research and Public Health 14, no. 4: 373. https://doi.org/10.3390/ijerph14040373
APA StyleSilva Lisboa, D., Santos, C., Barbosa, R. N., Magalhães, O., Paiva, L. M., Moreira, K. A., Lima, N., & Souza-Motta, C. M. (2017). Requalification of a Brazilian Trichoderma Collection and Screening of Its Capability to Decolourise Real Textile Effluent. International Journal of Environmental Research and Public Health, 14(4), 373. https://doi.org/10.3390/ijerph14040373