Airborne PM2.5-Induced Hepatic Insulin Resistance by Nrf2/JNK-Mediated Signaling Pathway


Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents and Antibodies
2.2. Animals
2.3. Experimental Design
2.4. Blood and Tissue Collection and Homogenate Preparation
2.5. Measurement of Glucose and Hormone Levels
2.6. Biochemical Analysis
2.7. Quantitative Analysis of Gene Expression
2.8. Western Blotting
2.9. Statistical Analysis
3. Results
3.1. Effects of PM2.5 Exposure on Body Weight, Consumption of Food and Water, Liver Weight, and Biochemicals in Serum
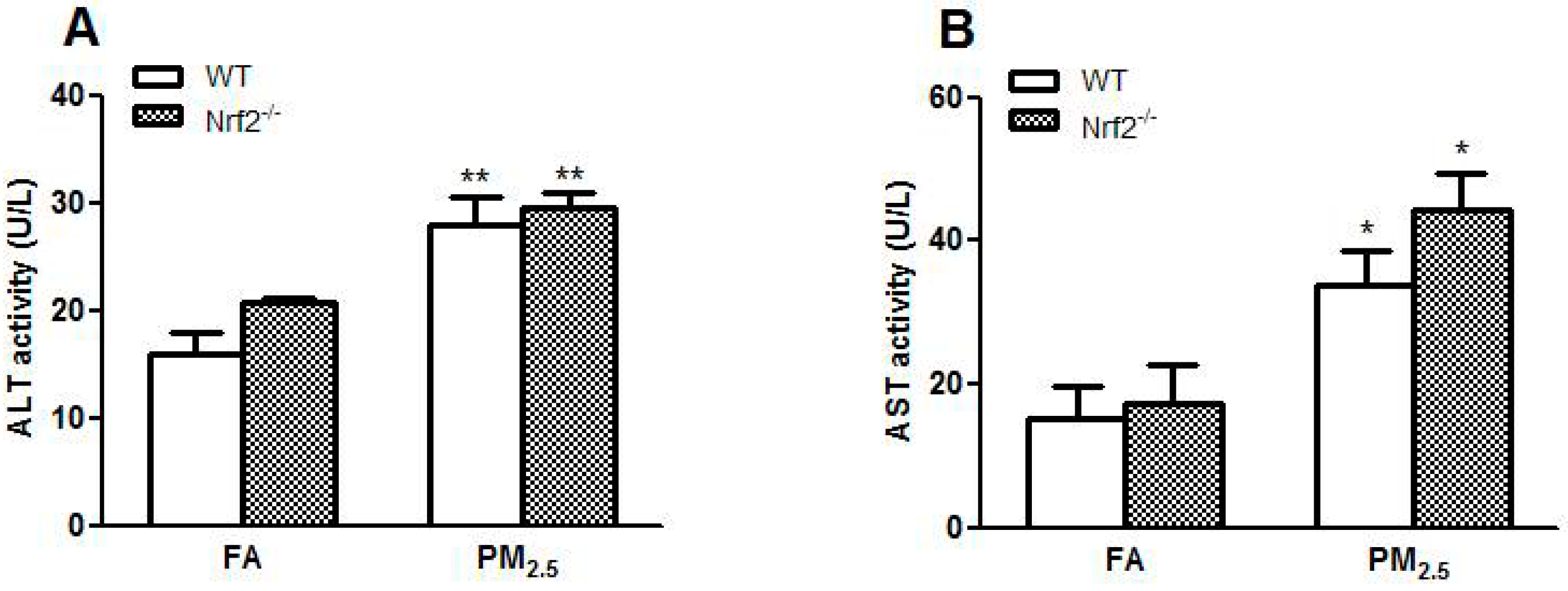
3.2. PM2.5 Exposure Induced Liver Injury
3.3. PM2.5 Exposure Induced Impaired Glucose Tolerance, and Insulin Resistance
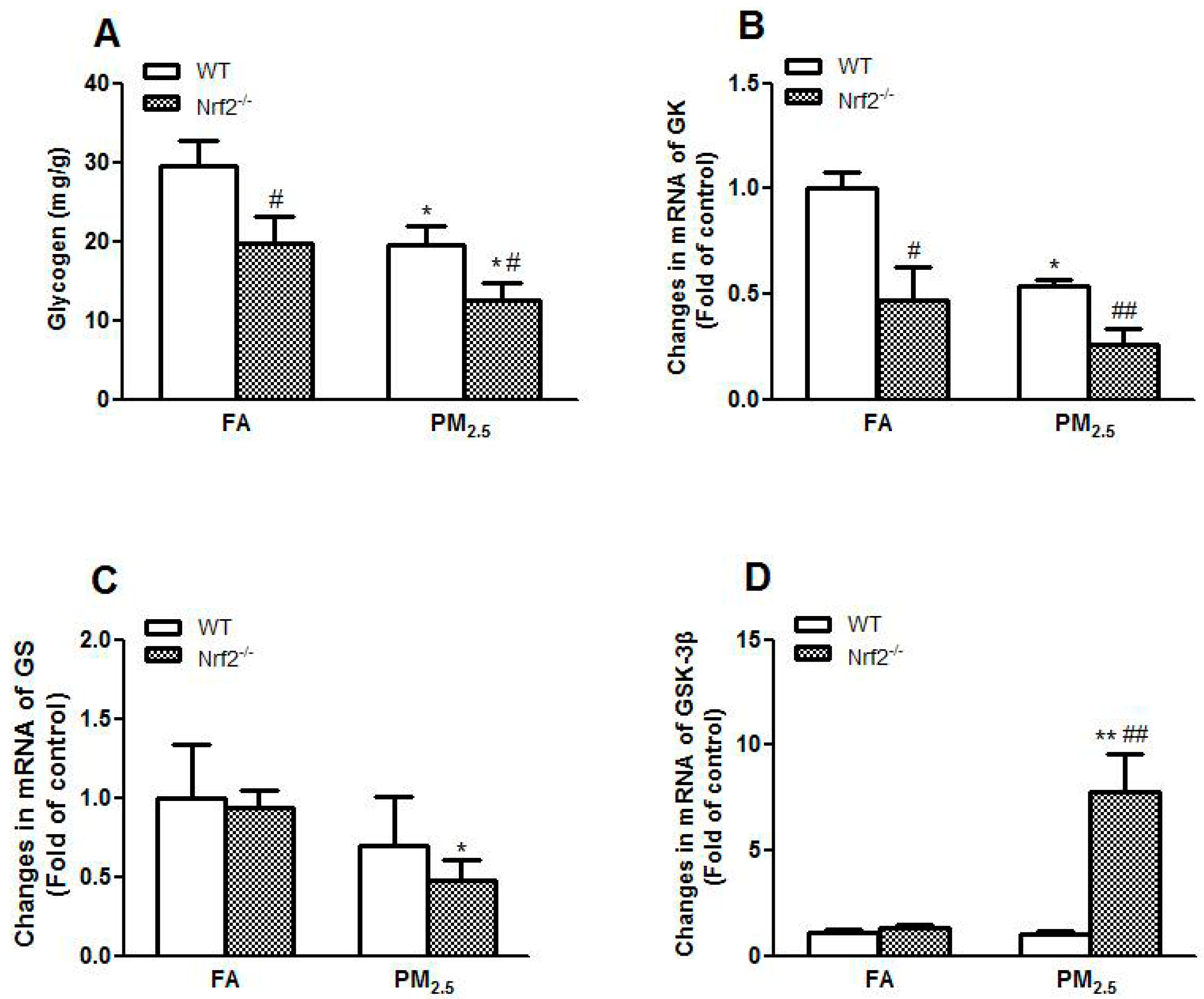
3.4. PM2.5 Exposure Induced Impaired Glycogen Storage through Decreasing Glycogen Synthesis in Liver of Mice
3.5. PM2.5 Exposure Induced Oxidative Responses in the Liver
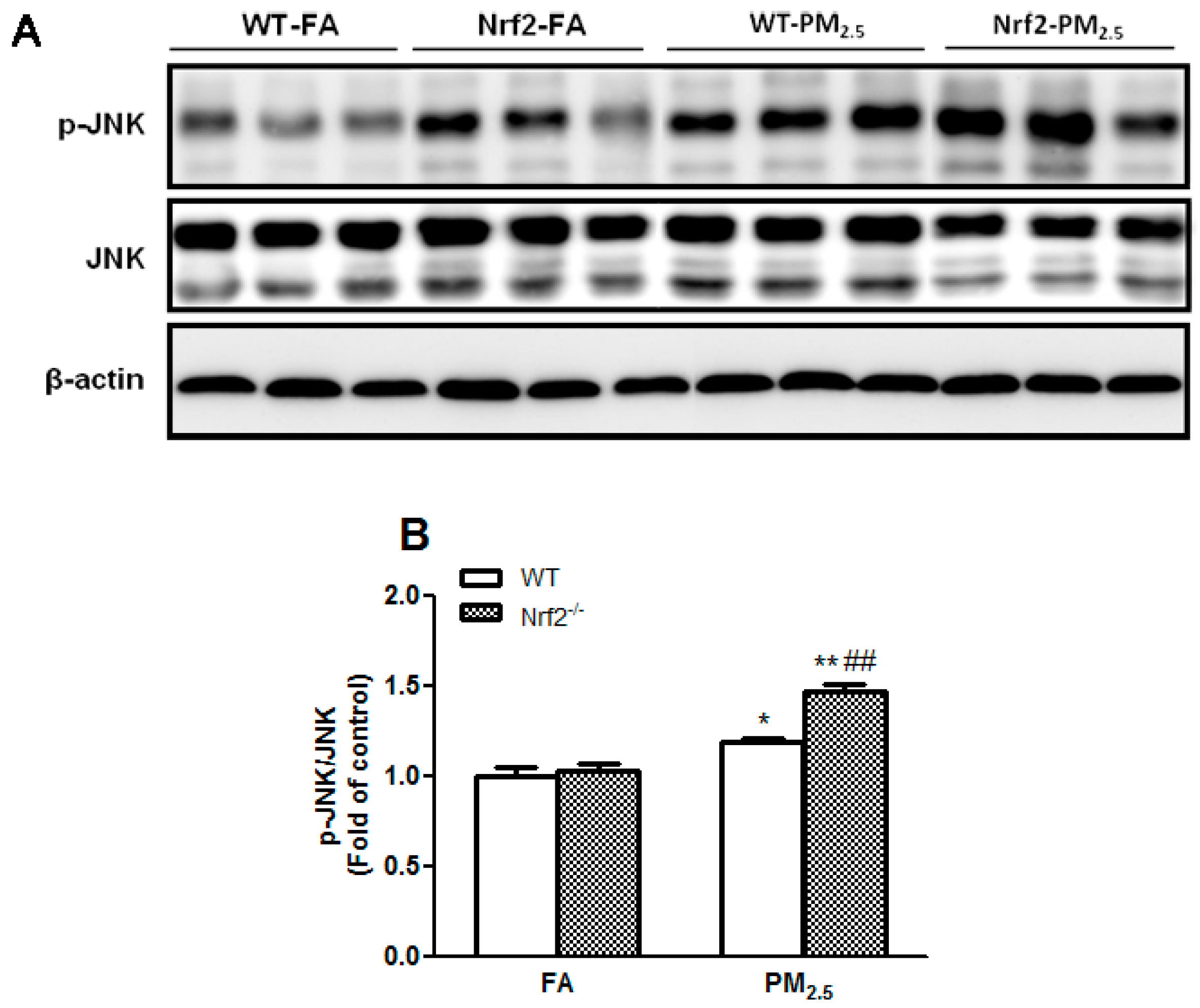
3.6. PM2.5 Exposure Activated JNK Signaling Pathway in the Liver Mice
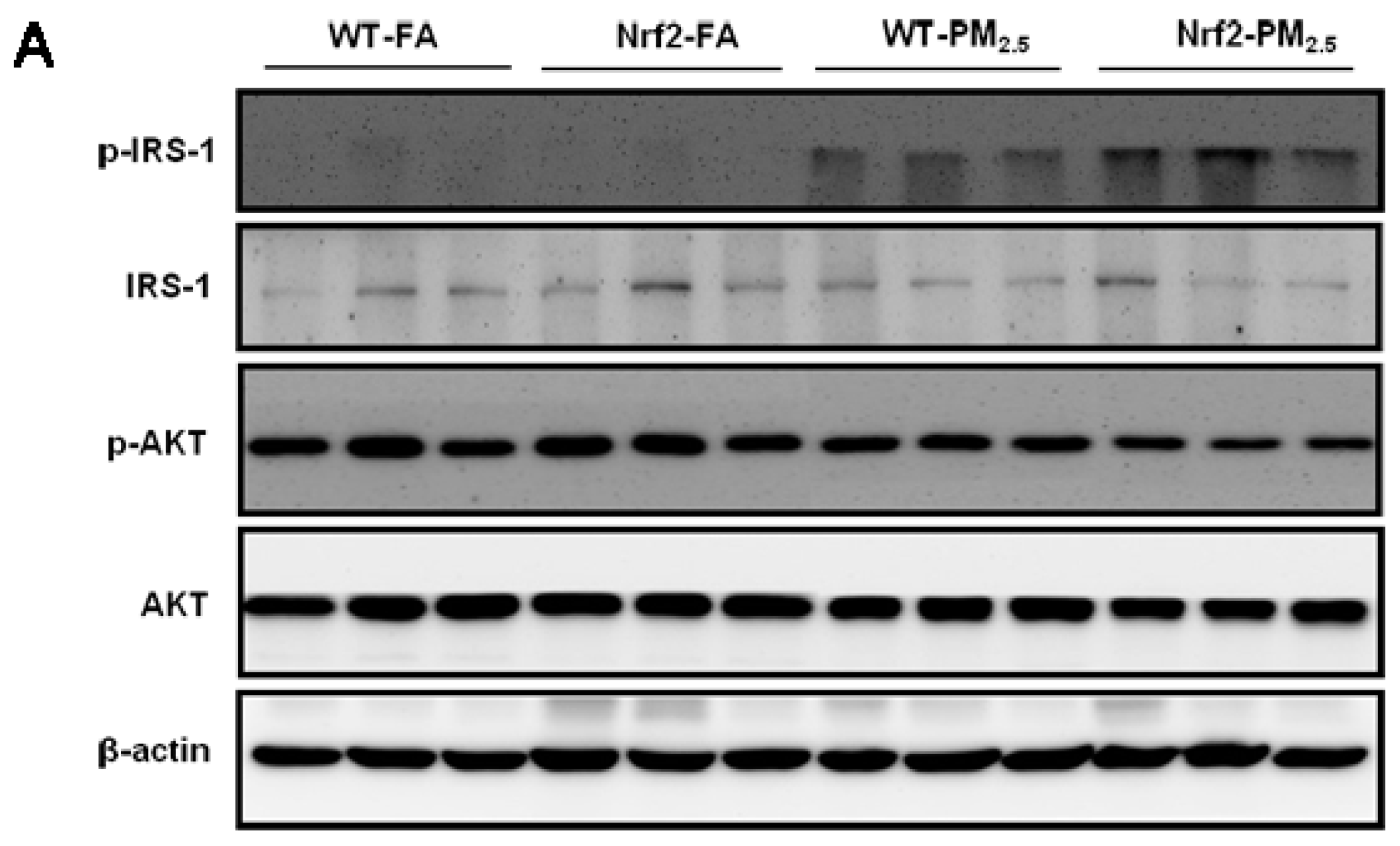
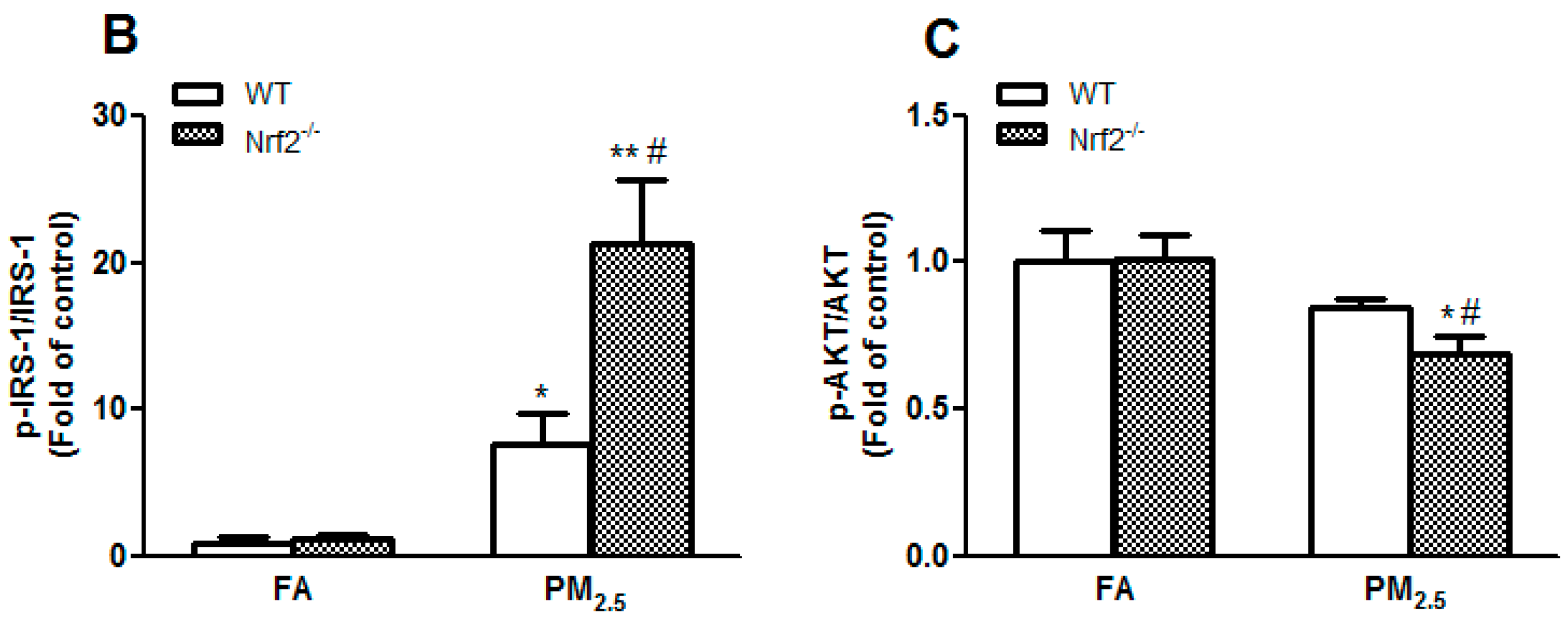
3.7. PM2.5 Exposure Induced Insulin Resistance via Suppressing the IRS-1/AKT Signaling Pathway
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Ostro, B.; Broadwin, R.; Green, S.; Feng, W.Y.; Lipsett, M. Fine particulate air pollution and mortality in nine California counties: Results from CALFINE. Environ. Health Perspect. 2006, 114, 29–33. [Google Scholar] [CrossRef] [PubMed]
- Zanobetti, A.; Schwartz, J. Ozone and survival in four cohorts with potentially predisposing diseases. Am. J. Respir. Crit. Care Med. 2011, 184, 836–841. [Google Scholar] [CrossRef] [PubMed]
- Oberdorster, G.; Sharp, Z.; Atudorei, V.; Elder, A.; Gelein, R.; Kreyling, W.; Cox, C. Translocation of inhaled ultrafine particles to the brain. Inhal. Toxicol. 2004, 16, 437–445. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.W.; Park, S.; Lim, C.W.; Lee, K.; Kim, B. The role of air pollutants in initiating liver disease. Toxicol. Res. 2014, 30, 65–70. [Google Scholar] [CrossRef] [PubMed]
- Brook, R.D. Cardiovascular effects of air pollution. Clin. Sci. 2008, 115, 175–187. [Google Scholar] [CrossRef] [PubMed]
- Balti, E.V.; Echouffo-Tcheugui, J.B.; Yako, Y.Y.; Kengne, A.P. Air pollution and risk of type 2 diabetes mellitus: A systematic review and meta-analysis. Diabetes Res. Clin. Pract. 2014, 106, 161–172. [Google Scholar] [CrossRef] [PubMed]
- Esposito, K.; Petrizzo, M.; Maiorino, M.I.; Bellastella, G.; Giugliano, D. Particulate matter pollutants and risk of type 2 diabetes: A time for concern? Endocrine 2016, 51, 32–37. [Google Scholar] [CrossRef] [PubMed]
- Eze, I.C.; Hemkens, L.G.; Bucher, H.C.; Hoffmann, B.; Schindler, C.; Kunzli, N.; Schikowski, T.; Probst-Hensch, N.M. Association between ambient air pollution and diabetes mellitus in Europe and North America: Systematic review and meta-analysis. Environ. Health Perspect. 2015, 123, 381–389. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.Q.; Ying, Z.K.; Harkema, J.; Sun, Q.H.; Rajagopalan, S. Epidemiological and experimental links between air pollution and type 2 diabetes. Toxicol. Pathol. 2013, 41, 361–373. [Google Scholar] [CrossRef] [PubMed]
- Park, S.K.; Wang, W. Ambient air pollution and type 2 diabetes: A systematic review of epidemiologic research. Curr. Environ. Health Rep. 2014, 1, 275–286. [Google Scholar] [CrossRef] [PubMed]
- Rao, X.Q.; Patel, P.; Puett, R.; Rajagopalan, S. Air pollution as a risk factor for type 2 diabetes. Toxicol. Sci. 2015, 143, 231–241. [Google Scholar] [CrossRef] [PubMed]
- Kolterman, O.G.; Insel, J.; Saekow, M.; Olefsky, J.M. Mechanisms of insulin resistance in human obesity: Evidence for receptor and postreceptor defects. J. Clin. Investig. 1980, 65, 1272–1284. [Google Scholar] [CrossRef] [PubMed]
- Cleeman, J.I.; Grundy, S.M.; Becker, D.; Clark, L.T.; Cooper, R.S.; Denke, M.A.; Howard, W.J.; Hunninghake, D.B.; Illingworth, D.R.; Luepker, R.V.; et al. Executive summary of the third report of the national cholesterol education program (NCEP) expert panel on detection, evaluation, and treatment of high blood cholesterol in adults (adult treatment panel III). JAMA J. Am. Med. Assoc. 2001, 285, 2486–2497. [Google Scholar]
- Bjornholm, M.; Kawano, Y.; Lehtihet, M.; Zierath, J.R. Insulin receptor substrate-1 phosphorylation and phosphatidylinositol 3-kinase activity in skeletal muscle from NIDDM subjects after in vivo insulin stimulation. Diabetes 1997, 46, 524–527. [Google Scholar] [CrossRef] [PubMed]
- Sorensen, M.; Daneshvar, B.; Hansen, M.; Dragsted, L.O.; Hertel, O.; Knudsen, L.; Loft, S. Personal PM2.5 exposure and markers of oxidative stress in blood. Environ. Health Perspect. 2003, 111, 161–165. [Google Scholar] [CrossRef] [PubMed]
- Deng, X.B.; Rui, W.; Zhang, F.; Ding, W.J. PM2.5 induces Nrf2-mediated defense mechanisms against oxidative stress by activating PI3K/AKT signaling pathway in human lung alveolar epithelial A549 cells. Cell Biol. Toxicol. 2013, 29, 143–157. [Google Scholar] [CrossRef] [PubMed]
- Browning, J.D.; Horton, J.D. Molecular mediators of hepatic steatosis and liver injury. J. Clin. Investig. 2004, 114, 147–152. [Google Scholar] [CrossRef] [PubMed]
- Sun, Q.H.; Yue, P.; Deiuliis, J.A.; Lumeng, C.N.; Kampfrath, T.; Mikolaj, M.B.; Cai, Y.; Ostrowski, M.C.; Lu, B.; Parthasarathy, S.; et al. Ambient air pollution exaggerates adipose inflammation and insulin resistance in a mouse model of diet-induced obesity. Circulation 2009, 119, 538–591. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Z.; Xu, X.H.; Zhang, X.B.; Wang, A.X.; Zhang, C.B.; Huttemann, M.; Grossman, L.I.; Chen, L.C.; Rajagopalan, S.; Sun, Q.H.; et al. Exposure to ambient particulate matter induces a NASH-like phenotype and impairs hepatic glucose metabolism in an animal model. J. Hepatol. 2013, 58, 148–154. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.H.; Yavar, Z.B.; Verdin, M.; Ying, Z.K.; Mihai, G.; Kampfrath, T.; Wang, A.X.; Zhong, M.H.; Lippmann, M.; Chen, L.C.; et al. Effect of early particulate air pollution exposure on obesity in mice role of p47(phox). Arterioscl. Throm. Vas. Biol. 2010, 30, 2518–2527. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.D. Mechanistic studies of the Nrf2-keap1 signaling pathway. Drug Metab. Rev. 2006, 38, 769–789. [Google Scholar] [CrossRef] [PubMed]
- Satoh, H.; Moriguchi, T.; Taguchi, K.; Takai, J.; Maher, J.M.; Suzuki, T.; Winnard, P.T.; Raman, V.; Ebina, M.; Nukiwa, T.; et al. Nrf2-deficiency creates a responsive microenvironment for metastasis to the lung. Carcinogenesis 2010, 31, 1833–1843. [Google Scholar] [CrossRef] [PubMed]
- Cheng, X.H.; Siow, R.C.M.; Mann, G.E. Impaired redox signaling and antioxidant gene expression in endothelial cells in diabetes: A role for mitochondria and the nuclear factor-E2-related factor 2-kelch-like ECH-associated protein 1 defense pathway. Antioxid. Redox Signal. 2011, 14, 469–487. [Google Scholar] [CrossRef] [PubMed]
- Sykiotis, G.P.; Habeos, I.G.; Samuelson, A.V.; Bohmann, D. The role of the antioxidant and longevity-promoting Nrf2 pathway in metabolic regulation. Curr. Opin. Clin. Nutr. Metab. Care 2011, 14, 41–48. [Google Scholar] [CrossRef] [PubMed]
- Yoh, K.; Hirayama, A.; Ishizaki, K.; Yamada, A.; Takeuchi, M.; Yamagishi, S.; Morito, N.; Nakano, T.; Ojima, M.; Shimohata, H.; et al. Hyperglycemia induces oxidative and nitrosative stress and increases renal functional impairment in Nrf2-deficient mice. Genes Cells 2008, 13, 1159–1170. [Google Scholar] [CrossRef] [PubMed]
- Aleksunes, L.M.; Reisman, S.A.; Yeager, R.L.; Goedken, M.J.; Klaassen, C.D. Nuclear factor erythroid 2-related factor 2 deletion impairs glucose tolerance and exacerbates hyperglycemia in type 1 diabetic mice. J. Pharmacol. Exp. Ther. 2010, 333, 140–151. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.Y.; Barnes, G.T.; Yang, Q.; Tan, Q.; Yang, D.S.; Chou, C.J.; Sole, J.; Nichols, A.; Ross, J.S.; Tartaglia, L.A.; et al. Chronic inflammation in fat plays a crucial role in the development of obesity-related insulin resistance. J. Clin. Investig. 2003, 112, 1821–1830. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.H.; Liu, C.Q.; Xu, Z.B.; Tzan, K.; Zhong, M.H.; Wang, A.X.; Lippmann, M.; Chen, L.C.; Rajagopalan, S.; Sun, Q.H. Long-term exposure to ambient fine particulate pollution induces insulin resistance and mitochondrial alteration in adipose tissue. Toxicol. Sci. 2011, 124, 88–98. [Google Scholar] [CrossRef] [PubMed]
- Hu, S.W.; Chang, Y.G.; Wang, J.F.; Xue, C.H.; Li, Z.J.; Wang, Y.M. Fucosylated chondroitin sulfate from sea cucumber in combination with rosiglitazone improved glucose metabolism in the liver of the insulin-resistant mice. Biosci. Biotech. Biochem. 2013, 77, 2263–2268. [Google Scholar] [CrossRef] [PubMed]
- Ebersbach-Silva, P.; Alves, T.; Fonseca, A.T.S.; Oliveira, M.A.D.; Machado, U.F.; Seraphim, P.M. Cigarette smoke exposure severely reduces peripheral insulin sensitivity without changing GLUT4 expression in oxidative muscle of wistar rats. Arq. Bras. Endocrinol. 2013, 57, 19–26. [Google Scholar] [CrossRef]
- Liu, C.Q.; Xu, X.H.; Bai, Y.T.; Wang, T.Y.; Rao, X.Q.; Wang, A.X.; Sun, L.X.; Ying, Z.K.; Gushchina, L.; Maiseyeu, A.; et al. Air pollution-mediated susceptibility to inflammation and insulin resistance: Influence of CCR2 pathways in mice. Environ. Health Perspect. 2014, 122, 17–26. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Li, W.; Li, Y.; Wang, Q.; Gao, L.; Zhao, J.J. Dietary lycium barbarum polysaccharide induces Nrf2/ARE pathway and ameliorates insulin resistance induced by high-fat via activation of PI3K/AKT signaling. Oxidat. Med. Cell. Longev. 2014, 2014, 145641. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.N.; Li, T.G.; Han, P. The effect of Tianmai Xiaoke Pian on insulin resistance through PI3-K/AKT signal pathway. J. Diabetes Res. 2016, 2016, 9261259. [Google Scholar] [CrossRef] [PubMed]
- Bloch-Damti, A.; Bashan, N. Proposed mechanisms for the induction of insulin resistance by oxidative stress. Antioxid. Redox Signal. 2005, 7, 1553–1567. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Zhao, L.P.; Mao, J.N.; Huang, J.; Chen, J.C. Antioxidant effects of hydrogen sulfide on left ventricular remodeling in smoking rats are mediated via PI3K/AKT-dependent activation of Nrf2. Toxicol. Sci. 2015, 144, 197–203. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.S.; Choi, I.W.; Han, M.H.; Kim, G.Y.; Hong, S.H.; Park, C.; Hwang, H.J.; Kim, C.M.; Kim, B.W.; Choi, Y.H. The cytoprotective effects of 7,8-dihydroxyflavone against oxidative stress are mediated by the upregulation of Nrf2-dependent HO-1 expression through the activation of the PI3K/AKT and ERK pathways in C2C12 myoblasts. Int. J. Mol. Med. 2015, 36, 501–510. [Google Scholar] [CrossRef] [PubMed]
- Itoh, K.; Ishii, T.; Wakabayashi, N.; Yamamoto, M. Regulatory mechanisms of cellular response to oxidative stress. Free Radical Res. 1999, 31, 319–324. [Google Scholar] [CrossRef]
- Wang, G.H.; Rajagopalan, S.; Sun, Q.H.; Zhang, K. Real-world exposure of airborne particulate matter triggers oxidative stress in an animal model. Int. J. Physiol. Pathophysiol. Pharmacol. 2010, 2, 64–68. [Google Scholar]
- Liu, F.; Xie, M.; Chen, D.; Li, J.; Ding, W. Effect of V(IV)O(dipic-Cl)(H2O)2 on lipid metabolism disorders in the liver of stz-induced diabetic rats. J. Diabetes Res. 2013, 2013, 956737. [Google Scholar] [CrossRef] [PubMed]
- Xia, F.Z.; Xu, X.; Zhai, H.L.; Meng, Y.; Zhang, H.X.; Du, S.C.; Xu, H.; Wu, H.; Lu, Y.L. Castration-induced testosterone deficiency increases fasting glucose associated with hepatic and extra-hepatic insulin resistance in adult male rats. Reprod. Biol. Endocrin. 2013, 11, 106. [Google Scholar] [CrossRef] [PubMed]
- Matthews, D.R.; Hosker, J.P.; Rudenski, A.S.; Naylor, B.A.; Treacher, D.F.; Turner, R.C. Homeostasis model assessment-insulin resistance and beta-cell function from fasting plasma-glucose and insulin concentrations in man. Diabetologia 1985, 28, 412–419. [Google Scholar] [CrossRef] [PubMed]
- Sorbi, D.; Boynton, J.; Lindor, K.D. The ratio of aspartate aminotransferase to alanine aminotransferase: Potential value in differentiating nonalcoholic steatohepatitis from alcoholic liver disease. Am. J. Gastroenterol. 1999, 94, 1018–1022. [Google Scholar] [CrossRef] [PubMed]
- Kretzschmar, M. Regulation of hepatic glutathione metabolism and its role in hepatotoxicity. Exp. Toxicol. Pathol. 1996, 48, 439–446. [Google Scholar] [CrossRef]
- Kamata, H.; Honda, S.; Maeda, S.; Chang, L.; Hirata, H.; Karin, M. Reactive oxygen species promote TNFα-induced death and sustained JNK activation by inhibiting MAP kinase phosphatases. Cell 2005, 120, 649–661. [Google Scholar] [CrossRef] [PubMed]
- Ventura, J.J. JNK potentiates TNF-stimulated necrosis by increasing the production of cytotoxic reactive oxygen species. Genes Dev. 2004, 18, 2905–2915. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.H.; Giraud, J.; Davis, R.J.; White, M.F. C-jun n-terminal kinase (JNK) mediates feedback inhibition of the insulin signaling cascade. J. Biol. Chem. 2002, 278, 2896–2902. [Google Scholar] [CrossRef] [PubMed]
- Pearson, J.F.; Bachireddy, C.; Shyamprasad, S.; Goldfine, A.B.; Brownstein, J.S. Association between fine particulate matter and diabetes prevalence in the U.S. Diabetes Care 2010, 33, 2196–2201. [Google Scholar] [CrossRef] [PubMed]
- Tan, Y.; Ichikawa, T.; Li, J.Q.; Si, Q.S.; Yang, H.T.; Chen, X.B.; Goldblatt, C.S.; Meyer, C.J.; Li, X.K.; Cai, L.; et al. Diabetic downregulation of Nrf2 activity via ERK contributes to oxidative stress-induced insulin resistance in cardiac cells in vitro and in vivo. Diabetes 2011, 60, 625–633. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.H.; Salam, M.T.; Eckel, S.P.; Breton, C.V.; Gilliland, F.D. Chronic effects of air pollution on respiratory health in Southern California children: Findings from the Southern California children’s health study. J. Thorac. Dis. 2015, 7, 46–58. [Google Scholar] [PubMed]
- Moller, P.; Danielsen, P.H.; Karottki, D.G.; Jantzen, K.; Roursgaard, M.; Klingberg, H.; Jensen, D.M.; Christophersen, D.V.; Hemmingsen, J.G.; Cao, Y.; et al. Oxidative stress and inflammation generated DNA damage by exposure to air pollution particles. Mutat. Res.-Rev. Mutat. 2014, 762, 133–166. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.S.; Eum, K.D.; Fang, S.C.; Rodrigues, E.G.; Modest, G.A.; Christiani, D.C. Oxidative stress and systemic inflammation as modifiers of cardiac autonomic responses to particulate air pollution. Int. J. Cardiol. 2014, 176, 166–170. [Google Scholar] [CrossRef] [PubMed]
- Bagul, P.K.; Middela, H.; Matapally, S.; Padiya, R.; Bastia, T.; Madhusudana, K.; Reddy, B.R.; Chakravarty, S.; Banerjee, S.K. Attenuation of insulin resistance, metabolic syndrome and hepatic oxidative stress by resveratrol in fructose-fed rats. Pharmacol. Res. 2012, 66, 260–268. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.Q.; Liu, H.L.; Davies, K.J.A.; Sioutas, C.; Finch, C.E.; Morgan, T.E.; Forman, H.J. Nrf2-regulated phase II enzymes are induced by chronic ambient nanoparticle exposure in young mice with age-related impairments. Free Radic. Biol. Med. 2012, 52, 2038–2046. [Google Scholar] [CrossRef] [PubMed]
- Yi, S.; Zhang, F.; Qu, F.; Ding, W.J. Water-insoluble fraction of airborne particulate matter (PM10) induces oxidative stress in human lung epithelial A549 cells. Environ. Toxicol. 2014, 29, 226–233. [Google Scholar] [CrossRef] [PubMed]
- Embi, N.; Rylatt, D.B.; Cohen, P. Glycogen synthase kinase-3 from rabbit skeletal muscle. Separation from cyclic-AMP-dependent protein kinase and phosphorylase kinase. Eur. J. Biochem. 1980, 107, 519–527. [Google Scholar] [CrossRef] [PubMed]
- Eldar-Finkelman, H.; Schreyer, S.A.; Shinohara, M.M.; LeBoeuf, R.C.; Krebs, E.G. Increased glycogen synthase kinase-3 activity in diabetes- and obesity-prone C57BL/6J mice. Diabetes 1999, 48, 1662–1666. [Google Scholar] [CrossRef] [PubMed]
- Nikoulina, S.E.; Ciaraldi, T.P.; Mudaliar, S.; Mohideen, P.; Carter, L.; Henry, R.R. Potential role of glycogen synthase kinase-3 in skeletal muscle insulin resistance of type 2 diabetes. Diabetes 2000, 49, 263–271. [Google Scholar] [CrossRef] [PubMed]
- Salameh, W.; Helliwell, J.P.; Han, G.; McPhaul, L.; Khorram, O. Expression of endometrial glycogen synthase kinase-3 beta protein throughout the menstrual cycle and its regulation by progesterone. Mol. Hum. Reprod. 2006, 12, 543–549. [Google Scholar] [CrossRef] [PubMed]
- Kaspar, J.W.; Niture, S.K.; Jaiswal, A.K. Nrf2:Inrf2 (keap1) signaling in oxidative stress. Free Radic. Biol. Med. 2009, 47, 1304–1309. [Google Scholar] [CrossRef] [PubMed]
- Kaneto, H.; Matsuoka, T.; Nakatani, Y.; Kawamori, D.; Miyatsuka, T.; Matsuhisa, M.; Yamasaki, Y. Oxidative stress, ER stress, and the JNK pathway in type 2 diabetes. J. Mol. Med. 2005, 83, 429–439. [Google Scholar] [CrossRef] [PubMed]
- Kaneto, H. The JNK pathway as a therapeutic target for diabetes. Expert Opin. Ther. Targets 2005, 9, 581–592. [Google Scholar] [CrossRef] [PubMed]
- Nakatani, Y.; Kaneto, H.; Kawamori, D.; Hatazaki, M.; Miyatsuka, T.; Matsuoka, T.; Kajimoto, Y.; Matsuhisa, M.; Yamasaki, Y.; Hori, M. Modulation of the JNK pathway in liver affects insulin resistance status. J. Biol. Chem. 2004, 279, 45803–45809. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Xu, G.; Bai, Z.S.; Li, K.C.; Yan, J.Y.; Li, F.; Ma, S.; Xu, H.B.; Huang, K.X. Selenite exacerbates hepatic insulin resistance in mouse model of type 2 diabetes through oxidative stress-mediated JNK pathway. Toxicol. Appl. Pharmacol. 2015, 289, 409–418. [Google Scholar] [CrossRef] [PubMed]
- Bi, X.; Wang, P.; Ma, Q.; Han, L.; Wang, X.; Mu, Y.; Guan, P.; Qu, X.; Wang, Z.; Huang, X. Anti-inflammatory activities and liver protection of Alisol F and 25-anhydroalisol F through the inhibition of MAPK, STAT3, and NF-κB activation in vitro and in vivo. Molecules 2017, 22, 951. [Google Scholar]
- Yang, P.; Zhao, Y.F.; Zhao, L.; Yuan, J.; Chen, Y.; Varghese, Z.; Moorhead, J.F.; Chen, Y.X.; Ruan, X.Z. Paradoxical effect of rapamycin on inflammatory stress-induced insulin resistance in vitro and in vivo. Sci. Rep. 2015, 5, 13. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.T.; Song, L.Y.; Ju, W.H.; Wang, X.G.; Dong, L.; Zhang, Y.N.; Ya, P.; Yang, C.; Li, F.S. The acute airway inflammation induced by PM2.5 exposure and the treatment of essential oils in BALB/C mice. Sci. Rep. 2017, 7, 13. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.Q.; Bai, Y.T.; Xu, X.H.; Sun, L.X.; Wang, A.X.; Wang, T.Y.; Maurya, S.K.; Periasamy, M.; Morishita, M.; Harkema, J.; et al. Exaggerated effects of particulate matter air pollution in genetic type 2 diabetes mellitus. Part. Fibre Toxicol. 2014, 11, 14. [Google Scholar] [CrossRef] [PubMed]
- Haberzettl, P.; O’Toole, T.; Bhatnagar, A.; Conklin, D.J. Exposure to fine particulate air pollution causes vascular insulin resistance by inducing pulmonar oxidative stress. Environ. Health Perspect. 2016, 124, 1830–1839. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | Forward Primer (5′-3′) | Reverse Primer (5′-3′) |
|---|---|---|
| Nrf2 | CTGAACTCCTGGACGGGACTA | CGGTGGGTCTCCGTAAATGG |
| HO-1 | GATAGAGCGCAACAAGCAGAA | CAGTGAGGCCCATACCAGAAG |
| GS | ACCAAGGCCAAAACGACAG | GGGCTCACATTGTTCTACTTGA |
| GSK-3β | ACCCTCATTACCTGACCTT | CTCAACTTAACAGACGGCT |
| GK | GGAACCAACTTCAGGGTGATG | CTGGTGTTTCGTCTTCACGCT |
| Actin | GTGACGTTGACATCCGTAAAGA | GCCGGACTCATCGTACTCC |
| Groups | Final Body Weight (g) | Food Consumption (g/mouse/day) | Water Consumption (g/mouse/day) | Liver Weight (g) | Glucose (mmol/L) | Insulin (μIU/mL) |
|---|---|---|---|---|---|---|
| WT-FA | 30.8 ± 0.76 | 4.27 | 4.33 | 1.16 ± 0.03 | 7.3 ± 0.31 | 12.7 ± 1.60 |
| WT-PM2.5 | 29.9 ± 1.18 | 4.19 | 3.59 | 1.26 ± 0.06 * | 7.5 ± 0.59 | 14.4 ± 1.62 |
| Nrf2-FA | 29.3 ± 0.58 | 5.44 | 3.82 | 1.11 ± 0.05 | 7.5 ± 0.23 | 13.5 ± 1.14 |
| Nrf2-PM2.5 | 29.9 ± 0.68 | 4.66 | 2.87 | 1.21 ± 0.10 | 8.0 ± 0.61 * | 20.0 ± 0.40 *,# |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, J.; Zhang, W.; Lu, Z.; Zhang, F.; Ding, W. Airborne PM2.5-Induced Hepatic Insulin Resistance by Nrf2/JNK-Mediated Signaling Pathway. Int. J. Environ. Res. Public Health 2017, 14, 787. https://doi.org/10.3390/ijerph14070787
Xu J, Zhang W, Lu Z, Zhang F, Ding W. Airborne PM2.5-Induced Hepatic Insulin Resistance by Nrf2/JNK-Mediated Signaling Pathway. International Journal of Environmental Research and Public Health. 2017; 14(7):787. https://doi.org/10.3390/ijerph14070787
Chicago/Turabian StyleXu, Jinxia, Wei Zhang, Zhongbing Lu, Fang Zhang, and Wenjun Ding. 2017. "Airborne PM2.5-Induced Hepatic Insulin Resistance by Nrf2/JNK-Mediated Signaling Pathway" International Journal of Environmental Research and Public Health 14, no. 7: 787. https://doi.org/10.3390/ijerph14070787
APA StyleXu, J., Zhang, W., Lu, Z., Zhang, F., & Ding, W. (2017). Airborne PM2.5-Induced Hepatic Insulin Resistance by Nrf2/JNK-Mediated Signaling Pathway. International Journal of Environmental Research and Public Health, 14(7), 787. https://doi.org/10.3390/ijerph14070787