Exogenous GR24 Alleviates Cadmium Toxicity by Reducing Cadmium Uptake in Switchgrass (Panicum virgatum) Seedlings

Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Culture and Experimental Treatments
2.2. Determination of Plant Biomass and Relative Water Content
2.3. Estimation of Root Morphology
2.4. Determination of Cd, Subcellular Distribution and Microelement Contents
2.5. Determination of Photosynthetic Pigment and Photosynthetic Gas Exchange
2.6. Estimation of Lipid Peroxidation and the Antioxidant Enzyme Activities
2.7. Determination of Endogenous Strigolactones Content
2.8. Statistical Analysis
3. Results
3.1. GR24 Improved Plant Biomass and Relative Water Content of Switchgrass Seedlings under Cd Stress
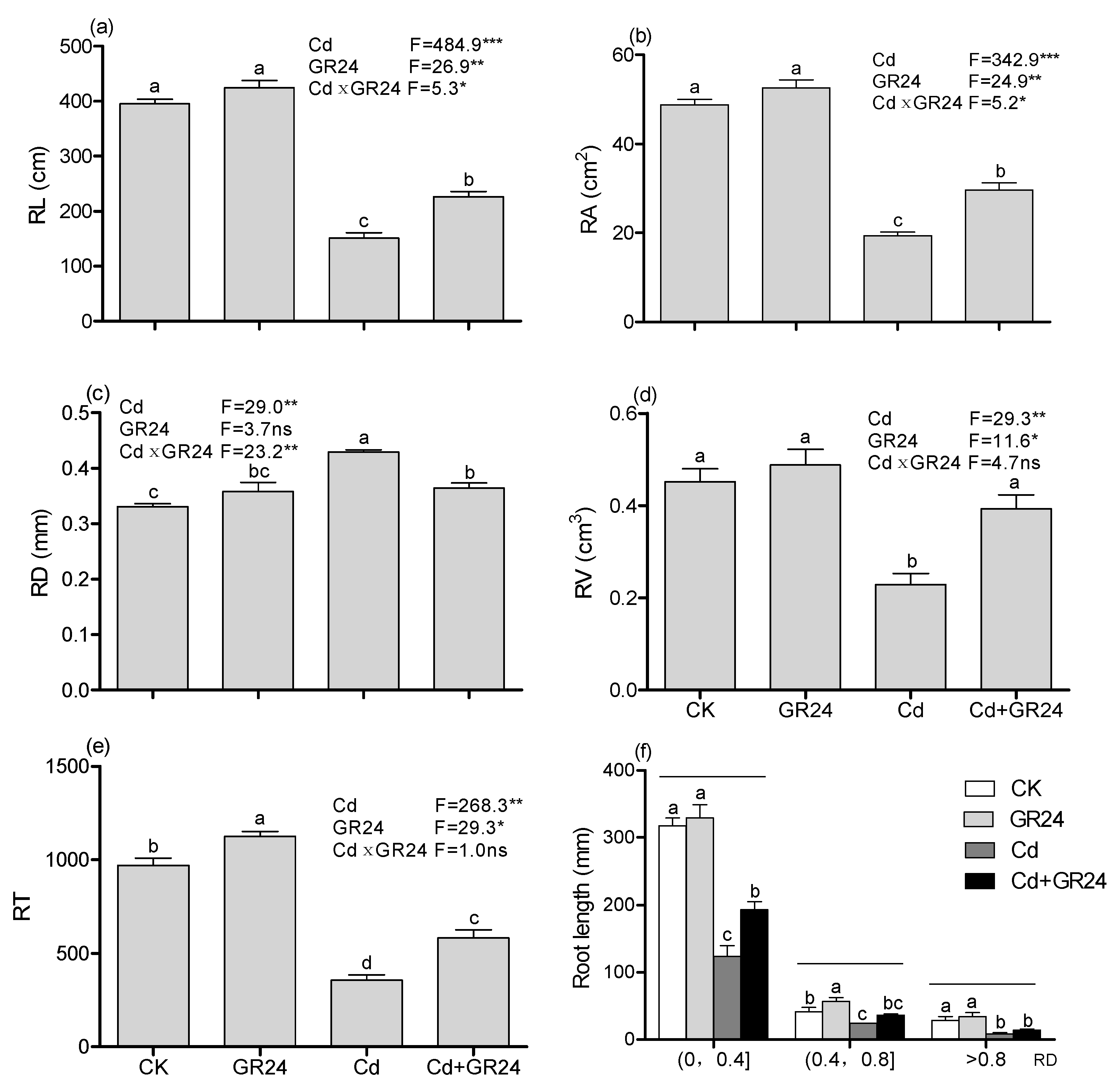
3.2. Effects of GR24 on Root Morphology of Switchgrass Seedlings under Cd Stress
3.3 GR24 Reduced Cd Concentrations in Plant Tissues and at a Subcellular Level
3.4. Effects of GR24 on Microelement Uptake in Plant Root and Shoot
3.5. Effects of GR24 on Photosynthetic Pigments and Photosynthetic Gas Exchange under Cd Stress
3.6. Effects of GR24 on Lipid Peroxidation and Antioxidant Enzyme Activity under Cd Stress
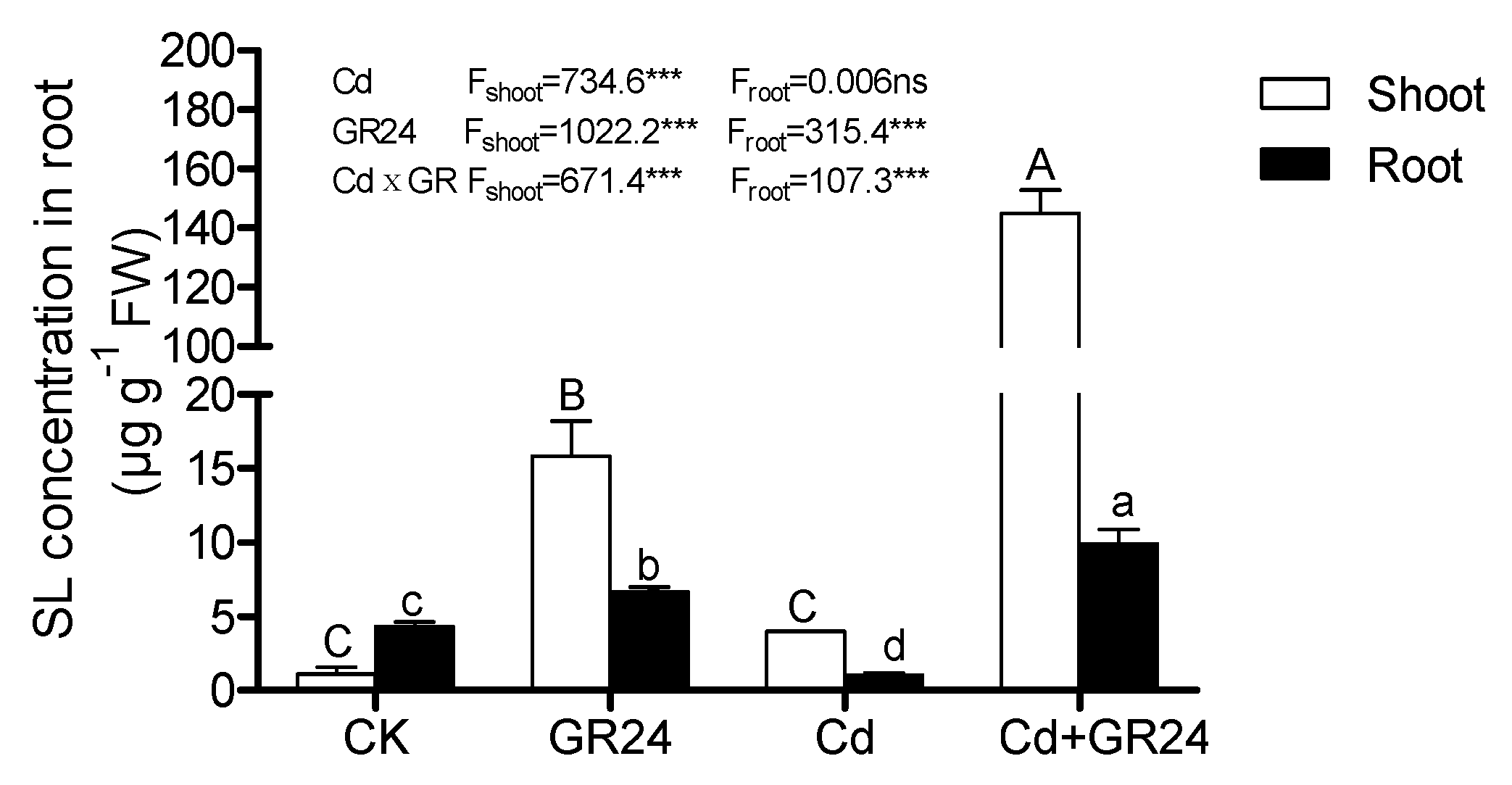
3.7. Endogenous Strigolactones in Shoots and Roots
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Tian, S.; Lu, L.; Zhang, J.; Wang, K.; Brown, P.; He, Z.; Liang, J.; Yang, X. Calcium protects roots of Sedum alfredii H. against cadmium-induced oxidative stress. Chemosphere 2011, 84, 63–69. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Yu, N.; Mu, G.; Shinwari, K.I.; Shen, Z.; Zheng, L. Screening for Cd-Safe Cultivars of Chinese Cabbage and a Preliminary Study on the Mechanisms of Cd Accumulation. Int. J. Environ. Res. Public Health 2017, 14, 395. [Google Scholar] [CrossRef] [PubMed]
- Azzi, V.S.; Kanso, A.; Kobeissi, A.; Kazpard, V.; Lartiges, B.; El Samrani, A. Effect of Cadmium on Lactuca sativa Grown in Hydroponic Culture Enriched with Phosphate Fertilizer. J. Environ. Prot. 2015, 6, 1337. [Google Scholar] [CrossRef]
- Benavides, M.P.; Gallego, S.M.; Tomaro, M.L. Cadmium toxicity in plants. Brazilian. J. Plant Physiol. 2005, 17, 21–34. [Google Scholar]
- López-Millán, A.F.; Sagardoy, R.; Solanas, M.; Abadía, A.; Abadía, J. Cadmium toxicity in tomato (Lycopersicon esculentum) plants grown in hydroponics. J. Environ. Exp. Bot. 2009, 65, 376–385. [Google Scholar] [CrossRef]
- Mobin, M.; Khan, N.A. Photosynthetic activity, pigment composition and antioxidative response of two mustard (Brassica juncea) cultivars differing in photosynthetic capacity subjected to cadmium stress. J. Plant Physiol. 2007, 164, 601–610. [Google Scholar] [CrossRef] [PubMed]
- Hall, J. Cellular mechanisms for heavy metal detoxification and tolerance. J. Environ. Exp. Bot. 2002, 53, 1–11. [Google Scholar] [CrossRef]
- Zhang, C.; Zhang, P.; Mo, C.; Yang, W.; Li, Q.; Pan, L.; Lee, D. Cadmium uptake, chemical forms, subcellular distribution, and accumulation in Echinodorus osiris Rataj. Environ. Sci. Processes Impacts 2013, 15, 1459–1465. [Google Scholar] [CrossRef] [PubMed]
- Marzec, M. Strigolactones as part of the plant defence system. Trends Plant Sci. 2016, 21, 900–903. [Google Scholar] [CrossRef] [PubMed]
- Koltai, H. Strigolactones are regulators of root development. New Phytol. 2011, 190, 545–549. [Google Scholar] [CrossRef] [PubMed]
- Andreo-Jimenez, B.; Ruyter-Spira, C.; Bouwmeester, H.J.; Lopez-Raez, J.A. Ecological relevance of strigolactones in nutrient uptake and other abiotic stresses, and in plant-microbe interactions below-ground. Plant Soil 2015, 394, 1–19. [Google Scholar] [CrossRef]
- Sadeghpour, A.; Hashemi, M.; Herbert, S.J. A simple vigor test for adjusting switchgrass seeding rate in marginal and fertile soils. Grassl. Sci. 2014, 60, 252–255. [Google Scholar] [CrossRef]
- Sharma, N.; Piscioneri, I.; Pignatelli, V. An evaluation of biomass yield stability of switchgrass (Panicum virgatum L.) cultivars. Energy Convers. Manag. 2003, 44, 2953–2958. [Google Scholar] [CrossRef]
- Liu, C.; Lou, L.; Deng, J.; Li, D.; Yuan, S.; Cai, Q. Morph-physiological responses of two switchgrass (Panicum virgatum L.) cultivars to cadmium stress. Grassl. Sci. 2016, 62, 92–101. [Google Scholar] [CrossRef]
- Chen, B.C.; Lai, H.Y.; Lee, D.Y.; Juang, K.W. Using chemical fractionation to evaluate the phytoextraction of cadmium by switchgrass from Cd-contaminated soils. Ecotoxicology 2011, 20, 409–418. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.F.; Ghosal, A.; Alam, M.F.; Kabir, A.H. Remediation of cadmium toxicity in field peas (Pisum sativum L.) through exogenous silicon. Ecotoxicol. Environ. Saf. 2017, 135, 165–172. [Google Scholar] [CrossRef] [PubMed]
- Hawrylak-Nowak, B.; Dresler, S.; Matraszek, R. Exogenous malic and acetic acids reduce cadmium phytotoxicity and enhance cadmium accumulation in roots of sunflower plants. Plant Physiol. Biochem. 2015, 94, 225–234. [Google Scholar] [CrossRef] [PubMed]
- De Graaff, M.A.; Six, J.; Jastrow, J.D.; Schadt, C.W.; Wullschleger, S.D. Variation in root architecture among switchgrass cultivars impacts root decomposition rates. Soil Biol. Biochem. 2013, 58, 198–206. [Google Scholar] [CrossRef]
- Lai, H.Y. Subcellular distribution and chemical forms of cadmium in Impatiens walleriana in relation to its phytoextraction potential. Chemosphere 2015, 138, 370–376. [Google Scholar] [CrossRef] [PubMed]
- Ma, W.; Xu, W.; Xu, H.; Chen, Y.; He, Z.; Ma, M. Nitric oxide modulates cadmium influx during cadmium-induced programmed cell death in tobacco BY-2 cells. Planta 2010, 232, 325–335. [Google Scholar] [CrossRef] [PubMed]
- Lichtenthaler, H.K. Chlorophylls and carotenoids: pigments of photosynthetic biomembranes. Methods Enzymol. 1987, 148, 350–382. [Google Scholar]
- Zhang, X.; Zhao, X.; Wang, Z.; Shen, W.; Xu, X. Protective effects of hydrogen-rich water on the photosynthetic apparatus of maize seedlings (Zea mays L.) as a result of an increase in antioxidant enzyme activities under high light stress. Plant Growth Regul. 2015, 77, 43–56. [Google Scholar] [CrossRef]
- Li, H.S.; Zhao, S. Principles and Techniques of Plant Physiological Biochemical Experiment; Beijing Higher Education Press: Beijing, China, 2000; pp. 195–197. [Google Scholar]
- Li, J.T.; Qiu, Z.B.; Zhang, X.W.; Wang, L.S. Exogenous hydrogen peroxide can enhance tolerance of wheat seedlings to salt stress. Acta Physiol. Plant. 2011, 33, 835–842. [Google Scholar] [CrossRef]
- Qiu, Z.; Guo, J.; Zhu, A.; Zhang, L.; Zhang, M. Exogenous jasmonic acid can enhance tolerance of wheat seedlings to salt stress. Ecotoxicol. Environ. Saf. 2014, 104, 202–208. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.; Chen, Q.; Liu, Q.; Zhang, W.; Ding, R. Exogenous silicon (Si) increases antioxidant enzyme activity and reduces lipid peroxidation in roots of salt-stressed barley (Hordeum vulgare L.). J. Plant Physiol. 2003, 160, 1157–1164. [Google Scholar] [CrossRef] [PubMed]
- López-Ráez, J.A.; Kohlen, W.; Charnikhova, T.; Mulder, P.; Undas, A.K.; Sergeant, M.J.; Verstappen, F.; Bugg, T.D.; Thompson, A.J.; Ruyter-Spira, C. Does abscisic acid affect strigolactone biosynthesis? New Phytol. 2010, 187, 343–354. [Google Scholar] [CrossRef] [PubMed]
- Agami, R.A.; Mohamed, G.F. Exogenous treatment with indole-3-acetic acid and salicylic acid alleviates cadmium toxicity in wheat seedlings. Ecotoxicol. Environ. Saf. 2013, 94, 164–171. [Google Scholar]
- Hsu, Y.T.; Kao, C.H. Toxicity in leaves of rice exposed to cadmium is due to hydrogen peroxide accumulation. Plant Soil 2007, 298, 231–241. [Google Scholar] [CrossRef]
- Lux, A.; Martinka, M.; Vaculík, M.; White, P.J. Root responses to cadmium in the rhizosphere: A review. J. Exp. Bot. 2011, 62, 21–37. [Google Scholar] [CrossRef] [PubMed]
- Wei, S.; Wang, S.; Li, Y.; Zhu, J. Root system responses of hyperaccumulator Solanum nigrum L. to Cd. J. Soils Sediments 2013, 13, 1069–1074. [Google Scholar] [CrossRef]
- Kapulnik, Y.; Resnick, N.; Mayzlish-Gati, E.; Kaplan, Y.; Wininger, S.; Hershenhorn, J.; Koltai, H. Strigolactones interact with ethylene and auxin in regulating root-hair elongation in Arabidopsis. J. Exp. Bot. 2011, 62, 2915–2924. [Google Scholar] [CrossRef] [PubMed]
- Rasmussen, A.; Mason, M.G.; De Cuyper, C.; Brewer, P.B.; Herold, S.; Agusti, J.; Geelen, D.; Greb, T.; Goormachtig, S. Beeckman, T. Strigolactones suppress adventitious rooting in Arabidopsis and pea. Plant Physiol. 2012, 158, 1976–1987. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, A.; Yamaji, N.; Yokosho, K.; Ma, J.F. Nramp5 is a major transporter responsible for manganese and cadmium uptake in rice. Plant Cell 2012, 24, 2155–2167. [Google Scholar] [CrossRef] [PubMed]
- Kısa, D.; Öztürk, L.; Tekin, Ş. Gene expression analysis of metallothionein and mineral elements uptake in tomato (Solanum lycopersicum) exposed to cadmium. J. Plant Res. 2016, 129, 989–995. [Google Scholar] [CrossRef] [PubMed]
- Anjum, S.A.; Tanveer, M.; Hussain, S.; Wang, L.; Khan, I.; Samad, R.A.; Tung, S.A.; Anam, M.; Shahzad, B. Morpho-Physiological growth and yield responses of two contrasting maize cultivars to cadmium exposure. CLEAN–Soil Air Water 2016, 44, 29–36. [Google Scholar] [CrossRef]
- Woo, H.R.; Kim, J.H.; Nam, H.G.; Lim, P.O. The delayed leaf senescence mutants of Arabidopsis, ore1, ore3, and ore9 are tolerant to oxidative stress. Plant Cell Physiol. 2004, 45, 923–932. [Google Scholar] [CrossRef] [PubMed]
- Snowden, K.C.; Simkin, A.J.; Janssen, B.J.; Templeton, K.R.; Loucas, H.M.; Simons, J.L.; Karunairetnam, S.; Gleave, A.P.; Clark, D.G.; Klee, H.J. The Decreased apical dominance1/Petunia hybrida CAROTENOID CLEAVAGE DIOXYGENASE8 gene affects branch production and plays a role in leaf senescence, root growth, and flower development. Plant Cell 2005, 17, 746–759. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Xing, F.; Xing, D. Characterization of target site of aluminum phytotoxicity in photosynthetic electron transport by fluorescence techniques in tobacco leaves. Plant Cell Physiol. 2012, 53, 1295–1309. [Google Scholar] [CrossRef] [PubMed]
- Mihailovic, N.; Drazic, G.; Vucinic, Z. Effects of aluminium on photosynthetic performance in Al-sensitive and Al-tolerant maize inbred lines. Photosynthetica 2008, 46, 476–480. [Google Scholar] [CrossRef]
- Šimonová, E.; Henselová, M.; Masarovičová, E.; Kohanová, J. Comparison of tolerance of Brassica juncea and Vigna radiata to cadmium. Biol. Plan. 2007, 51, 488–492. [Google Scholar] [CrossRef]
- De Azevedo Neto, A.D.; Prisco, J.T.; Enéas-Filho, J.; de Abreu, C.E.B.; Gomes-Filho, E. Effect of salt stress on antioxidative enzymes and lipid peroxidation in leaves and roots of salt-tolerant and salt-sensitive maize genotypes. Environ. Exp. Bot. 2006, 56, 87–94. [Google Scholar] [CrossRef]
- Shi, G.; Liu, C.; Cai, Q.; Liu, Q.; Hou, C. Cadmium accumulation and tolerance of two safflower cultivars in relation to photosynthesis and antioxidantive enzymes. Bull. Environ. Contam. Toxicol. 2010, 85, 256–263. [Google Scholar] [CrossRef] [PubMed]
- Meyer, C.-L.; Peisker, D.; Courbot, M.; Craciun, A.R.; Cazalé, A.-C.; Desgain, D.; Schat, H.; Clemens, S.; Verbruggen, N. Isolation and characterization of Arabidopsis halleri and Thlaspi caerulescens phytochelatin synthases. Planta 2011, 234, 83–95. [Google Scholar] [CrossRef] [PubMed]
- Bano, S.A.; Ashfaq, D. Role of mycorrhiza to reduce heavy metal stress. Nat. Sci. 2013, 5, 16. [Google Scholar] [CrossRef]
- Grace, E.; Cotsaftis, O.; Tester, M.; Smith, F.; Smith, S. Arbuscular mycorrhizal inhibition of growth in barley cannot be attributed to extent of colonization, fungal phosphorus uptake or effects on expression of plant phosphate transporter genes. New Phytol. 2009, 181, 938–949. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Fresh Weight (mg plant−1) | Dry Weight (mg plant−1) | Relative Water Content (%) | |||
|---|---|---|---|---|---|---|
| Root | Shoot | Root | Shoot | Root | ||
| CK | 507.0 ± 16.7 a | 745.5 ± 51.2a | 57.6 ± 2.9 a | 158.6 ± 20 a | 91.5 ± 2.78 a | |
| GR24 | 516.7 ± 98.5 a | 736.2.1 ± 104.9 a | 56 ± 5 a | 156.1 ± 15.7 a | 89.1 ± 2.2 a | |
| Cd | 183.0 ± 10.9 b | 251.5 ± 10.2 c | 32.9 ± 1.4 b | 71.4 ± 2.4 c | 82.8 ± 4.8 b | |
| Cd + GR24 | 291.6 ± 75.3 b | 423.7 ± 44.1b | 34.9 ± 5.3 b | 101.8 ± 13.1 b | 91.3 ± 0.65 a | |
| ANOVA (F value) | ||||||
| Cd | 45.5 *** | 123.9 *** | 20.6 ** | 34.1 *** | 3.5 ns | |
| GR24 | 5.4 * | 5.1 * | 13.4 ** | 8.5 * | 3.1 ns | |
| Cd × GR24 | 16.0 ** | 6.3 * | 6.1 * | 15.6 ** | 10.0 * | |
| Variable | Chlorophyll Content | Photosynthetic Parameters | |||
|---|---|---|---|---|---|
| Pn (µmol CO2 m−2 s−1) | Gs (mol (H2O) m−2 s−1) | Ci (µmol (CO2) mol−1) | Tr (mmol (H2O) m−2 s−1) | ||
| CK | 1.61 ± 0.07 a | 28.8 ± 1.9 a | 0.18 ± 0.004 a | 139.7 ± 4.7 a | 3.21 ± 0.2 a |
| GR24 | 1.38 ± 0.05 b | 27.7 ± 1.8 a | 0.17 ± 0.002 b | 136.0 ± 3.6 a | 3.17 ± 0.3 a |
| Cd | 1.07 ± 0.07 d | 15.8 ± 0.7 c | 0.12 ± 0.004 d | 98.7 ± 2.1 c | 2.09 ± 0.02 b |
| Cd + GR24 | 1.20 ± 0.064 c | 23.4 ± 0.1 b | 0.15 ± 0.003 c | 119.7 ± 2.1 c | 2.99 ± 0.1 a |
| ANOVA (F value) | |||||
| Cd | 97.0 *** | 129.1 *** | 476.6 *** | 224.1 *** | 38.6 *** |
| GR24 | 1.8 ns | 17.8 * | 63.6 *** | 20.5 ** | 16.7 ** |
| Cd × GR24 | 24.9 ** | 33. 1*** | 134.1 *** | 41.5 *** | 20.5 ** |
| Variable | MDA Content (nmol g−1 FW) | Enzyme Activity | ||
|---|---|---|---|---|
| SOD (U·g−1 FW) | POD (U·min−1·g−1 FW) | CAT(U·min−1 ·g−1 FW) | ||
| CK | 11.7 ± 5.4 b | 96.1 ± 5.7 d | 517.1 ± 15.3 b | 178.5 ± 5.4 d |
| GR24 | 10.0 ± 0.5 b | 107.8 ± 3.7 c | 520.3 ± 16.3 b | 201.5 ± 7.1 c |
| Cd | 17.8 ± 1.7 a | 121.3 ± 1.4 b | 601.1 ± 39.5 a | 250.5 ± 3.0 b |
| Cd+GR24 | 12.6 ± 3.1 b | 134.1 ± 1.8 a | 626.1 ± 43.0 a | 284.5 ± 10.9 a |
| ANOVA (F value) | ||||
| Cd | 16.8 *** | 155.4 *** | 27.7 *** | 349.5 *** |
| GR24 | 10.7 * | 34.8 * | 0.61 ns | 47.3 ** |
| Cd × GR24 | 2.9 ns | 0.07 ns | 0.36 ns | 1.8 ns |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tai, Z.; Yin, X.; Fang, Z.; Shi, G.; Lou, L.; Cai, Q. Exogenous GR24 Alleviates Cadmium Toxicity by Reducing Cadmium Uptake in Switchgrass (Panicum virgatum) Seedlings. Int. J. Environ. Res. Public Health 2017, 14, 852. https://doi.org/10.3390/ijerph14080852
Tai Z, Yin X, Fang Z, Shi G, Lou L, Cai Q. Exogenous GR24 Alleviates Cadmium Toxicity by Reducing Cadmium Uptake in Switchgrass (Panicum virgatum) Seedlings. International Journal of Environmental Research and Public Health. 2017; 14(8):852. https://doi.org/10.3390/ijerph14080852
Chicago/Turabian StyleTai, Zhenglan, Xinqiang Yin, Zhigang Fang, Gaoling Shi, Laiqing Lou, and Qingsheng Cai. 2017. "Exogenous GR24 Alleviates Cadmium Toxicity by Reducing Cadmium Uptake in Switchgrass (Panicum virgatum) Seedlings" International Journal of Environmental Research and Public Health 14, no. 8: 852. https://doi.org/10.3390/ijerph14080852
APA StyleTai, Z., Yin, X., Fang, Z., Shi, G., Lou, L., & Cai, Q. (2017). Exogenous GR24 Alleviates Cadmium Toxicity by Reducing Cadmium Uptake in Switchgrass (Panicum virgatum) Seedlings. International Journal of Environmental Research and Public Health, 14(8), 852. https://doi.org/10.3390/ijerph14080852