Removal and Biodegradation of 17β-Estradiol and Diethylstilbestrol by the Freshwater Microalgae Raphidocelis subcapitata

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Microalgae Species and Culture Conditions
2.2. Removal of Bacteria from Algal Cultures
2.3. Growth Inhibition Test
2.4. Residual Estrogens
2.5. Estrogens Adsorbed onto Cell Surface
2.6. Estrogens Absorbed into Cells
2.7. Determination of Estrogens
2.8. Statistical Analysis
3. Results
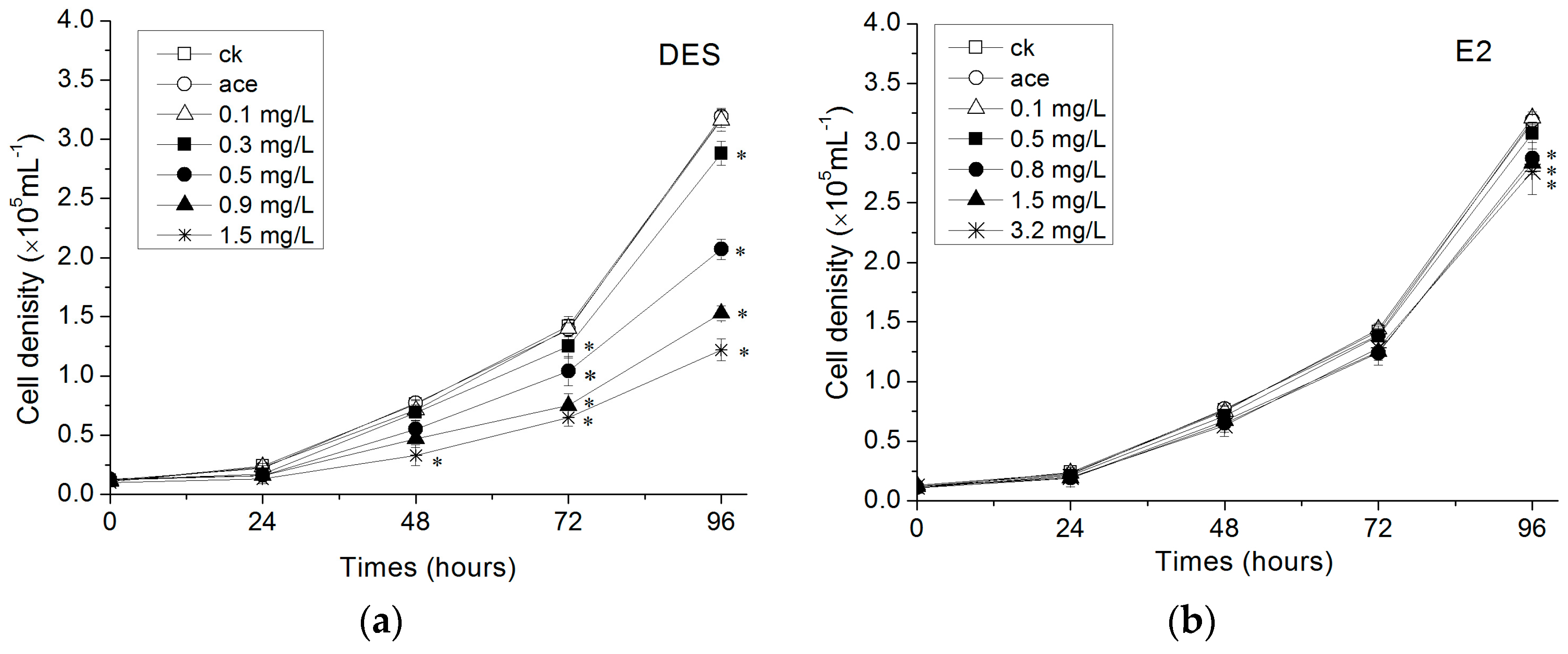
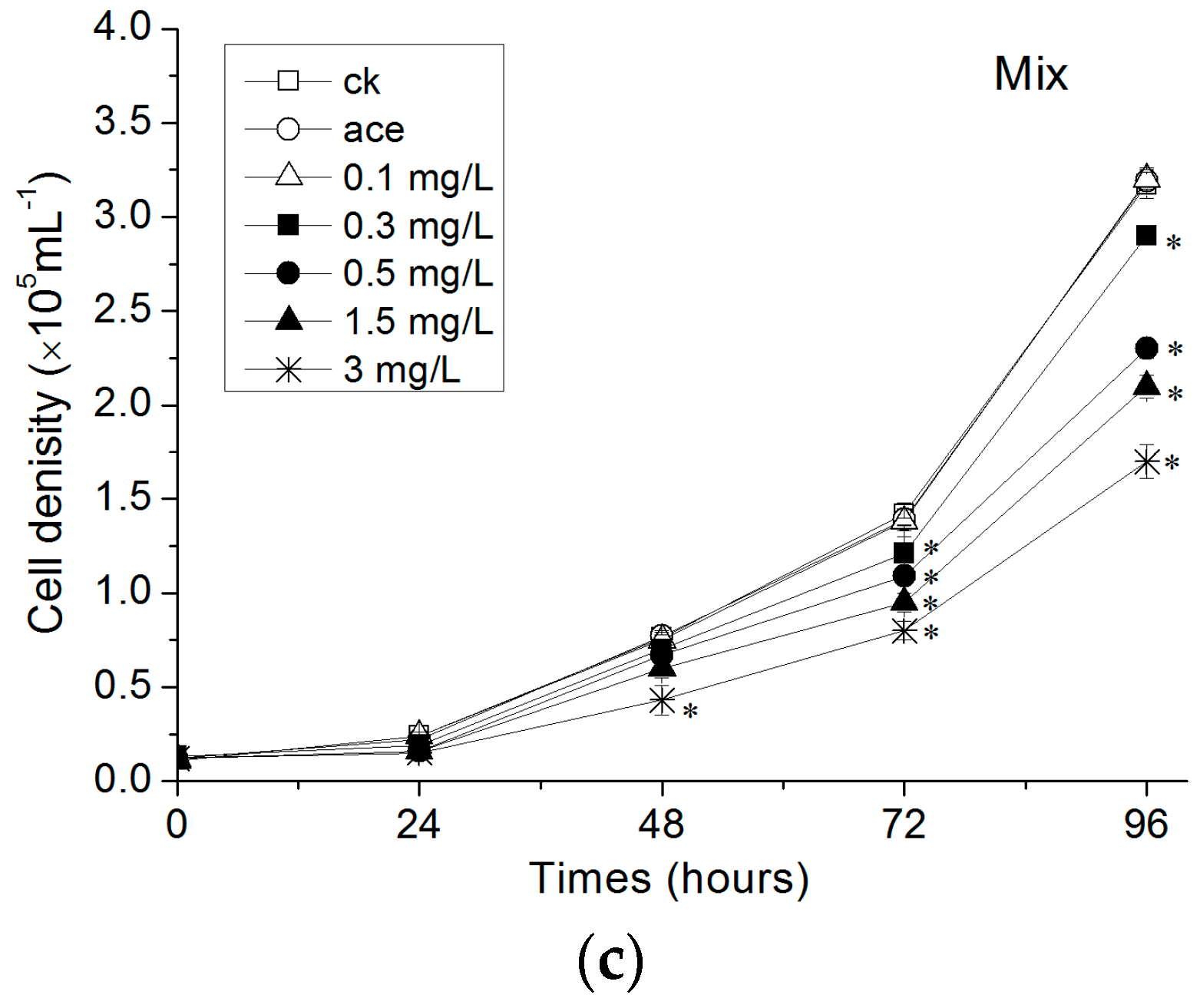
3.1. Growth of R. subcapitata Exposed to Estrogens
3.2. Removal of Estrogens by R. subcapitata
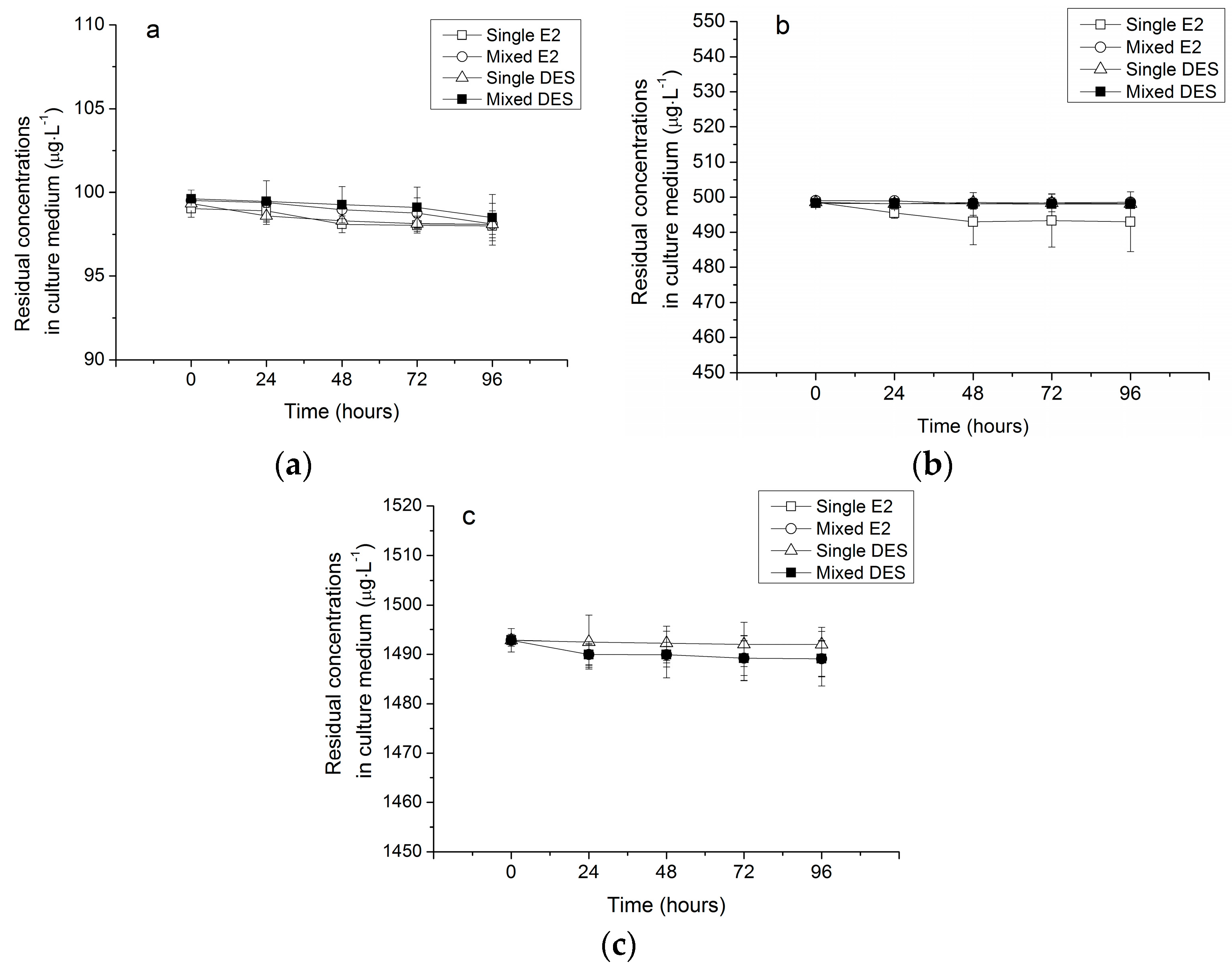
3.2.1. Abiotic Losses of Estrogens (AL)
3.2.2. Removal of Estrogens in Single Treatments by R. subcapitata
3.2.3. Removal of Estrogens in Mixed Treatments by R. subcapitata
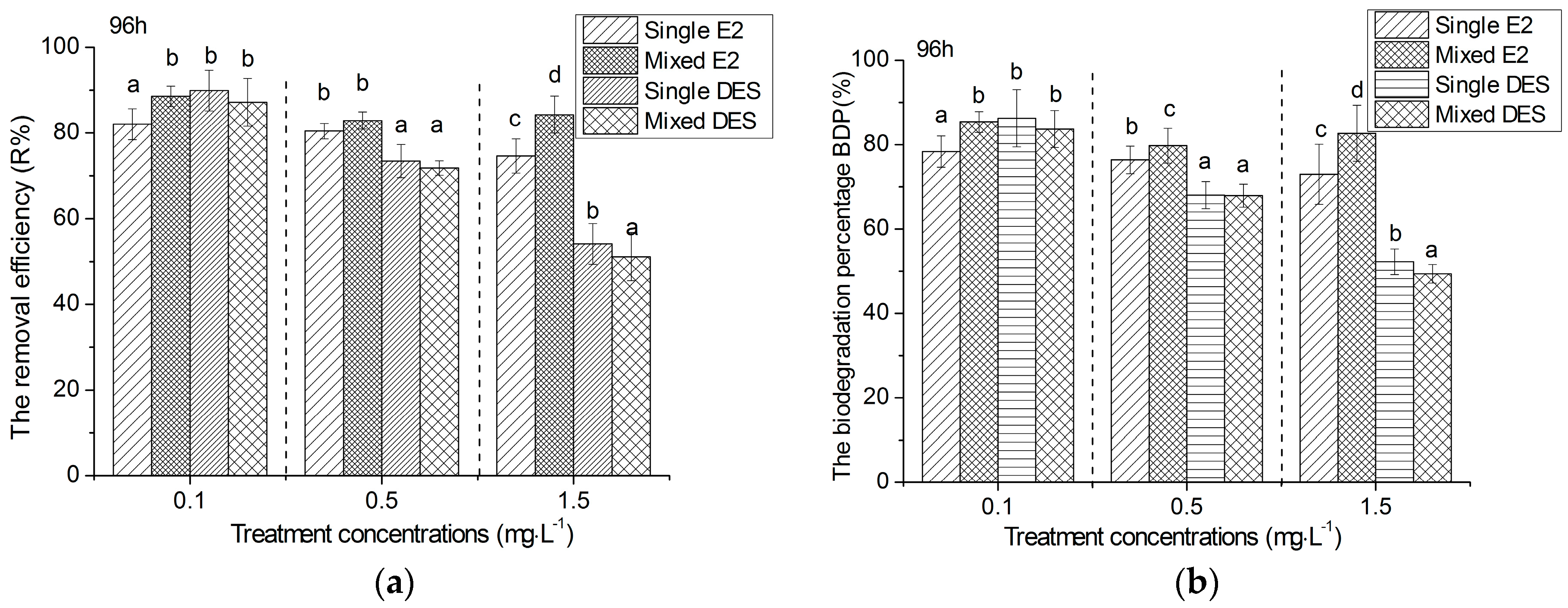
3.3. Removal Efficiency and Biodegradation of Estrogens by R. subcapitata
4. Discussion
4.1. Influence of Estrogens on R. subcapitata Growth
4.2. Capacity and Mechanism of R. subcapitata for the Removal of Estrogens
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Della Greca, M.; Pinto, G.; Pistillo, P.; Pollio, A.; Previtera, L.; Temussi, F. Biotransformation of ethinylestradiol by microalgae. Chemosphere 2008, 70, 2047–2053. [Google Scholar] [CrossRef] [PubMed]
- Khanal, S.K.; Xie, B.; Thompson, M.L.; Sung, S.; Ong, S.K.; Van Leeuwent, J. Fate, transport, and biodegradation of natural estrogens in the environment and engineered systems. Environ. Sci. Technol. 2006, 40, 6537–6546. [Google Scholar] [CrossRef] [PubMed]
- Kuster, M.; de Alda, M.J.; Hernando, M.D.; Petrovic, M.; Martin-Alonso, J.; Barcelo, D. Analysis and occurrence of pharmaceuticals, estrogens, progestogens and polar pesticides in sewage treatment plant effluents, river water and drinking water in the llobregat river basin (Barcelona, Spain). J. Hydrol. 2008, 358, 112–123. [Google Scholar] [CrossRef]
- Lei, B.L.; Peng, W.; Li, W.; Yu, Y.X.; Xu, J.; Wang, Y.P. Diethylstilbestrol at environmental levels affects the development of early life stage and target gene expression in japanese medaka (Oryzias latipes). Ecotoxicology 2016, 25, 563–573. [Google Scholar] [CrossRef] [PubMed]
- Adedeji, O.B.; Durhan, E.J.; Garcia-Reyero, N.; Kahl, M.D.; Jensen, K.M.; Lalone, C.A.; Makynen, E.A.; Perkins, E.J.; Thomas, L.; Villeneuve, D.L.; et al. Short-term study investigating the estrogenic potency of diethylstilbesterol in the fathead minnow (Pimephales promelas). Environ. Sci. Technol. 2012, 46, 7826–7835. [Google Scholar] [CrossRef] [PubMed]
- Yin, P.; Li, Y.W.; Chen, Q.L.; Liu, Z.H. Diethylstilbestrol, flutamide and their combination impaired the spermatogenesis of male adult zebrafish through disrupting hpg axis, meiosis and apoptosis. Aquat. Toxicol. 2017, 185, 129–137. [Google Scholar] [CrossRef] [PubMed]
- Baronti, C.; Curini, R.; D‘Ascenzo, G.; Di Corcia, A.; Gentili, A.; Samperi, R. Monitoring natural and synthetic estrogens at activated sludge sewage treatment plants and in a receiving river water. Environ. Sci. Technol. 2000, 34, 5059–5066. [Google Scholar] [CrossRef]
- Silva, C.P.; Otero, M.; Esteves, V. Processes for the elimination of estrogenic steroid hormones from water: A review. Environ. Pollut. 2012, 165, 38–58. [Google Scholar] [CrossRef] [PubMed]
- Esteban, S.; Gorga, M.; Petrovic, M.; Gonzalez-Alonso, S.; Barcelo, D.; Valcarcel, Y. Analysis and occurrence of endocrine-disrupting compounds and estrogenic activity in the surface waters of central spain. Sci. Total Environ. 2014, 466–467, 939–951. [Google Scholar] [CrossRef] [PubMed]
- Nie, M.; Yang, Y.; Liu, M.; Yan, C.; Shi, H.; Dong, W.; Zhou, J.L. Environmental estrogens in a drinking water reservoir area in Shanghai: Occurrence, colloidal contribution and risk assessment. Sci. Total Environ. 2014, 487, 785–791. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Gao, Y.; Li, Q.; Li, G.; Guo, Q.; Yan, C. Estrogenic compounds and estrogenicity in surface water, sediments, and organisms from yundang lagoon in Xiamen, China. Arch. Environ. Contam. Toxicol. 2011, 61, 93–100. [Google Scholar] [CrossRef] [PubMed]
- Nie, M.; Yan, C.; Dong, W.; Liu, M.; Zhou, J.; Yang, Y. Occurrence, distribution and risk assessment of estrogens in surface water, suspended particulate matter, and sediments of the yangtze estuary. Chemosphere 2015, 127, 109–116. [Google Scholar] [CrossRef] [PubMed]
- Rocha, S.; Domingues, V.F.; Pinho, C.; Fernandes, V.C.; Delerue-Matos, C.; Gameiro, P.; Mansilha, C. Occurrence of bisphenol a, estrone, 17β-estradiol and 17α ethinylestradiol in portuguese rivers. Bull. Environ. Contam. Toxicol. 2013, 90, 73–78. [Google Scholar] [CrossRef] [PubMed]
- D‘Ascenzo, G.; di Corcia, A.; Gentili, A.; Mancini, R.; Mastropasqua, R.; Nazzari, M.; Samperi, R. Fate of natural estrogen conjugates in municipal sewage transport and treatment facilities. Sci. Total Environ. 2003, 302, 199–209. [Google Scholar] [CrossRef]
- Praveena, S.M.; Lui, T.S.; Hamin, N.; Razak, S.Q.; Aris, A.Z. Occurrence of selected estrogenic compounds and estrogenic activity in surface water and sediment of langat river (Malaysia). Environ. Monit. Assess. 2016, 188, 442. [Google Scholar] [CrossRef] [PubMed]
- He, N.; Sun, X.; Zhong, Y.; Sun, K.; Liu, W.; Duan, S. Removal and biodegradation of nonylphenol by four freshwater microalgae. Int. J. Environ. Res. Public Health 2016, 13, 1239. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Wong, Y.S.; Tam, F.Y. Green microalgae in removal and biotransformation of estradiol and ethinylestradiol. J. Appl. Phycol. 2017, 29, 263–273. [Google Scholar] [CrossRef]
- Lai, K.M.; Scrimshaw, M.D.; Lester, J.N. Biotransformation and bioconcentration of steroid estrogens by Chlorella vulgaris. Appl. Environ. Microbiol. 2002, 68, 859–864. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.L.; Habteselassie, M.Y.; Resurreccion, E.P.; Mantripragada, V.; Peng, S.S.; Bauer, S.; Colosi, L.M. Evaluating removal of steroid estrogens by a model alga as a possible sustainability benefit of hypothetical integrated algae cultivation and wastewater treatment systems. ACS Sustain. Chem. Eng. 2014, 2, 2544–2553. [Google Scholar] [CrossRef]
- Hom-Diaz, A.; Llorca, M.; Rodríguez-Mozaz, S.; Vicent, T.; Barceló, D.; Blánquez, P. Microalgae cultivation on wastewater digestate: B-estradiol and 17α-ethynylestradiol degradation and transformation products identification. J. Environ. Manag. 2015, 155, 106–113. [Google Scholar] [CrossRef] [PubMed]
- Zhou, G.J.; Peng, F.Q.; Yang, B.; Ying, G.G. Cellular responses and bioremoval of nonylphenol and octylphenol in the freshwater green microalga scenedesmus obliquus. Ecotoxicol. Environ. Saf. 2012, 87, 10–16. [Google Scholar] [CrossRef] [PubMed]
- Peng, F.Q.; Ying, G.G.; Yang, B.; Liu, S.; Lai, H.J.; Liu, Y.S.; Chen, Z.F.; Zhou, G.J. Biotransformation of progesterone and norgestrel by two freshwater microalgae (scenedesmus obliquus and chlorella pyrenoidosa): Transformation kinetics and products identification. Chemosphere 2014, 95, 581–588. [Google Scholar] [CrossRef] [PubMed]
- Subashchandrabose, S.R.; Ramakrishnan, B.; Megharaj, M.; Venkateswarlu, K.; Naidu, R. Consortia of cyanobacteria/microalgae and bacteria: Biotechnological potential. Biotechnol. Adv. 2011, 29, 896–907. [Google Scholar] [CrossRef] [PubMed]
- Mishamandani, S.; Gutierrez, T.; Berry, D.; Aitken, M.D. Response of the bacterial community associated with a cosmopolitan marine diatom to crude oil shows a preference for the biodegradation of aromatic hydrocarbons. Environ. Microbiol. 2016, 18, 1817–1833. [Google Scholar] [CrossRef] [PubMed]
- Yamagishi, T.; Yamaguchi, H.; Suzuki, S.; Horie, Y.; Tatarazako, N. Cell reproductive patterns in the green alga pseudokirchneriella subcapitata (=selenastrum capricornutum) and their variations under exposure to the typical toxicants potassium dichromate and 3,5-dcp. PLoS ONE 2017, 12, e0171259. [Google Scholar] [CrossRef] [PubMed]
- Mp, G.D.L.; Olmos-Espejel, J.J.; Díaz-Flores, G.; Montaño-Montiel, A. Biodegradation of benzo(a)pyrene by two freshwater microalgae selenastrum capricornutum and scenedesmus acutus: A comparative study useful for bioremediation. Environ. Sci. Pollut. Res. Int. 2016, 23, 1–11. [Google Scholar]
- Chan, S.M.; Luan, T.; Wong, M.H.; Tam, N.F. Removal and biodegradation of polycyclic aromatic hydrocarbons by selenastrum capricornutum. Environ. Toxicol. Chem. 2010, 25, 1772–1779. [Google Scholar] [CrossRef]
- Lin, K.; Luo, L.J.; Ping, W.; Luan, T.G.; Tam, F. Effects of metals on biosorption and biodegradation of mixed polycyclic aromatic hydrocarbons by a freshwater green alga selenastrum capricornutum. Bioresour. Technol. 2010, 101, 6961–6972. [Google Scholar]
- Gao, Q.T.; Tam, N.F. Growth, photosynthesis and antioxidant responses of two microalgal species, Chlorella vulgaris and Selenastrum capricornutum, to nonylphenol stress. Chemosphere 2011, 82, 346–354. [Google Scholar] [CrossRef] [PubMed]
- Stanier, R.Y.; Kunisawa, R.; Mandel, M.; Cohen-Bazire, G. Purification and properties of unicellular blue-green algae (order chroococcales). Bacteriol. Rev. 1971, 35, 171–205. [Google Scholar] [PubMed]
- Su, J.Q.; Yang, X.; Zheng, T.L.; Hong, H.S. An efficient method to obtain axenic cultures of alexandrium tamarense—A psp-producing dinoflagellate. J. Microbiol. Methods 2007, 69, 425. [Google Scholar] [CrossRef] [PubMed]
- Kaczala, F.; Salomon, P.S.; Marques, M.; Graneli, E.; Hogland, W. Effects from log-yard stormwater runoff on the microalgae scenedesmus subspicatus: Intra-storm magnitude and variability. J. Hazard. Mater. 2011, 185, 732–739. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Wong, M.H.; Tam, N.F.Y. Antioxidant responses of two microalgae, selenastrum capricornutum and chlorella sp., to estradiol and ethinylestradiol. J. Appl. Phycol. 2013, 25, 891–903. [Google Scholar] [CrossRef]
- Organization for Economic Cooperation and Development (OECD). Test no. 201: Freshwater Alga and Cyanobacteria, Growth Inhibitiontest. OECD Guidelines for the Testing of Chemicals, Section 2; OECD Publishing: Paris, France, 2011. [Google Scholar]
- Salomao, A.L.D.; Soroldoni, S.; Marques, M.; Hogland, W.; Bila, D.M. Effects of single and mixed estrogens on single and combined cultures of d-subspicatus and p-subcapitata. Bull. Environ. Contam. Toxicol. 2014, 93, 215–221. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Hu, H.Y.; Gan, K.; Sun, Y.X. Effects of different nitrogen and phosphorus concentrations on the growth, nutrient uptake, and lipid accumulation of a freshwater microalga Scenedesmus sp. Bioresour. Technol. 2010, 101, 5494. [Google Scholar]
- Fischer, B.B.; Roffler, S.; Eggen, R.I.L. Multiple stressor effects of predation by rotifers and herbicide pollution on different chlamydomonas strains and potential impacts on population dynamics. Environ. Toxicol. Chem. 2012, 31, 2832–2840. [Google Scholar] [CrossRef] [PubMed]
- Kojima, H.; Goto, K.; Okamoto, H. Characterization of atpase activity associated with plasma-membrane from vigna hypocotyls. Plant Cell Physiol. 1984, 25, 1265–1276. [Google Scholar]
- Mesenko, M.M.; Ivanov, V.B. The effects of H+-atpase activator and inhibitors on cell growth in the maize root. Russ. J. Plant Physiol. 2005, 52, 497–503. [Google Scholar] [CrossRef]
- Anila, N.; Simon, D.P.; Chandrashekar, A.; Sarada, R. Glucose-induced activation of H+-atpase in Dunaliella salina and its role in hygromycin resistance. J. Appl. Phycol. 2013, 25, 121–128. [Google Scholar] [CrossRef]
- Chaoui, A.; El Ferjani, E. Heavy metal-induced oxidative damage is reduced by beta-estradiol application in lentil seedlings. Plant Growth Regul. 2014, 74, 1–9. [Google Scholar] [CrossRef]
- Upadhyay, P.; Maier, C. Effects of 17β-estradiol on growth, primary metabolism, phenylpropanoid-flavonoid pathways and pathogen resistance in Arabidopsis thaliana. Am. J. Plant Sci. 2016, 7, 1693–1710. [Google Scholar] [CrossRef]
- Andrade, L.R.; Leal, R.N.; Noseda, M.; Duarte, M.E.R.; Pereira, M.S.; Mourao, P.A.S.; Farina, M.; Amado, G.M. Brown algae overproduce cell wall polysaccharides as a protection mechanism against the heavy metal toxicity. Mar. Pollut. Bull. 2010, 60, 1482–1488. [Google Scholar] [CrossRef] [PubMed]
- Perales-Vela, H.V.; Pena-Castro, J.M.; Canizares-Villanueva, R.O. Heavy metal detoxification in eukaryotic microalgae. Chemosphere 2006, 64, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Sabatini, S.E.; Juarez, A.B.; Eppis, M.R.; Bianchi, L.; Luquet, C.M.; Rios de Molina Mdel, C. Oxidative stress and antioxidant defenses in two green microalgae exposed to copper. Ecotoxicol. Environ. Saf. 2009, 72, 1200–1206. [Google Scholar] [CrossRef] [PubMed]
- Tsang, C.K.; Lau, P.S.; Tam, N.F.; Wong, Y.S. Biodegradation capacity of tributyltin by two chlorella species. Environ. Pollut. 1999, 105, 289–297. [Google Scholar] [CrossRef]
- Nakajima, N.; Teramoto, T.; Kasai, F.; Sano, T.; Tamaoki, M.; Aono, M.; Kubo, A.; Kamada, H.; Azumi, Y.; Saji, H. Glycosylation of bisphenol a by freshwater microalgae. Chemosphere 2007, 69, 934–941. [Google Scholar] [CrossRef] [PubMed]
- Shi, W.; Wang, L.; Rousseau, D.P.; Lens, P.N. Removal of estrone, 17α-ethinylestradiol, and 17β-estradiol in algae and duckweed-based wastewater treatment systems. Environ. Sci. Pollut. Res. Int. 2010, 17, 824–833. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Chi, J. Biodegradation of phthalate acid esters by different marine microalgal species. Mar. Pollut. Bull. 2015, 99, 70–75. [Google Scholar] [CrossRef] [PubMed]
- Audrey, M.; Banu, O.; Edward, L.P.C. Use of sub-micron sized resin particles for removal of endocrine disrupting compounds and pharmaceuticals from water and wastewater. Acta Sci. Circumst. 2017, 51, 256–264. [Google Scholar]
- Xu, R.F.; Sun, M.X.; Liu, J.; Wang, H.; Li, X.; Zhu, X.Z.; Ling, W.T. Isolation, identification and characterization of a diethylstilbestrol-degrading bacterial strain Serratia sp. Acta Sci. Circumst. 2014, 35, 3169–3174. [Google Scholar]
- Kent, R.A.; Currie, D. Predicting algal sensitivity to a pesticide stress. Environ. Toxicol. Chem. 2010, 14, 983–991. [Google Scholar] [CrossRef]
- Tang, J.; Hoagland, K.D.; Siegfried, B.D. Uptake and bioconcentration of atrazine by selected freshwater algae. Environ. Toxicol. Chem. 1998, 17, 1085–1090. [Google Scholar] [CrossRef]
- Wu, X.L.; Wang, Y.Y.; Liang, R.X.; Dai, Q.Y.; Jin, D.C.; Chao, W.L. Biodegradation of an endocrine-disrupting chemical di-n-butyl phthalate by newly isolated Agrobacterium sp. and the biochemical pathway. Process Biochem. 2011, 46, 1090–1094. [Google Scholar] [CrossRef]
- Tsezos, M.; Bell, J.P. Comparison of the biosorption and desorption of hazardous organic pollutants by live and dead biomass. Water Res. 1989, 23, 561–568. [Google Scholar] [CrossRef]
- Xiong, J.Q.; Kurade, M.B.; Abou-Shanab, R.A.I.; Ji, M.K.; Choi, J.; Kim, J.O.; Jeon, B.H. Biodegradation of carbamazepine using freshwater microalgae chlamydomonas mexicana and scenedesmus obliquus and the determination of its metabolic fate. Bioresour. Technol. 2016, 205, 183–190. [Google Scholar] [CrossRef] [PubMed]
- Xu, N.; Xu, Y.F.; Xu, S.; Li, J.; Tao, H.C. Removal of estrogens in municipal wastewater treatment plants: A chinese perspective. Environ. Pollut. 2012, 165, 215–224. [Google Scholar] [CrossRef] [PubMed]
- Torres, M.A.; Barros, M.P.; Campos, S.C.; Pinto, E.; Rajamani, S.; Sayre, R.T.; Colepicolo, P. Biochemical biomarkers in algae and marine pollution: A review. Ecotoxicol. Environ. Saf. 2008, 71, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Maeda, S.; Wada, H.; Kumeda, K.; Onoue, M.; Ohki, A.; Higashi, S.; Takeshita, T. Methylation of inorganic arsenic by arsenic-tolerant freshwater algae. Appl. Organ. Chem. 2010, 1, 465–472. [Google Scholar] [CrossRef]
- Wang, S.; Poon, K.; Cai, Z. Removal and metabolism of triclosan by three different microalgal species in aquatic environment. J. Hazard. Mater. 2018, 342, 643–650. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Wang, S.; Tai, Y.; Tao, R.; Dai, Y.; Guo, J.; Yang, Y.; Duan, S. Biogenic manganese oxides generated by green algae Desmodesmus sp. Wr1 to improve bisphenol a removal. J. Hazard. Mater. 2017, 339, 310–319. [Google Scholar] [CrossRef] [PubMed]
- Omar, T.F.T.; Ahmad, A.; Aris, A.Z.; Yusoff, F.M. Endocrine disrupting compounds (EDCS) in environmental and biota matrices: Review of analytical strategies for pharmaceuticals, estrogenic hormones and alkylphenol compounds. TrAC Trends Anal. Chem. 2016, 85, 241–259. [Google Scholar] [CrossRef]
- Norvill, Z.N.; Toledocervantes, A.; Blanco, S.; Shilton, A.; Guieysse, B.; Muñoz, R. Photodegradation and sorption govern tetracycline removal during wastewater treatment in algal ponds. Bioresour. Technol. 2017, 232, 35–43. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Estrogens | EC50 | 95% Confidence |
|---|---|---|
| E2 | >3.2 mg·L−1 | - |
| DES | 1.011 mg·L−1 | 0.79–1.22 mg·L−1 |
| Treatments (mg/L) | Times (h) | Amount of Estrogens | |||||
|---|---|---|---|---|---|---|---|
| Residual (μg·L−1) | Extracellular (10−8 µg·cell−1) | Intracellular (10−8 µg·cell−1) | |||||
| E2 | DES | E2 | DES | E2 | DES | ||
| 0.1 | 24 | 29.2 ± 0.7 * | 41.2 ± 1.1 | 0.31 ± 0.08 | 0.45 ± 0.15 | 0.21 ± 0.05 | 0.23 ± 0.08 |
| 48 | 24.3 ± 1.4 * | 34.3 ± 1.8 | 0.25 ± 0.02 * | 0.85 ± 0.05 | 0.20 ± 0.08 * | 0.43 ± 0.05 | |
| 96 | 17.6 ± 1.7 | 9.9 ± 0.8 * | 0.31 ± 0.05 | 0.24 ± 0.05 | ND * | 0.15 ± 0.07 | |
| 0.5 | 24 | 213.1 ± 5.5 * | 268.7 ± 23.1 | 0.98 ± 0.24 * | 6.0 ± 0.25 | 1.45 ± 0.33 * | 2.12 ± 0.42 |
| 48 | 121.7 ± 11.5 * | 158.7 ± 13.3 | 0.88 ± 0.1 * | 2.11 ± 0.67 | 2.26 ± 0.12 * | 4.34 ± 0.15 | |
| 96 | 95.8 ± 10.6 * | 129.5 ± 11.4 | 0.45 ± 0.11 * | 3.33 ± 0.59 | 1.6 ± 0.21 * | 4.10 ± 0.15 | |
| 1.5 | 24 | 750.2 ± 18.1 * | 861.9 ± 10.6 | 14.8 ± 6.6 * | 18.1 ± 3.9 | 8.41 ± 2.1 * | 14.2 ± 5.51 |
| 48 | 501.5 ± 11.9 * | 778.1 ± 15.5 | 7.89 ± 0.93 * | 14.1 ± 0.75 | 6.32 ± 0.24 * | 18.2 ± 0.51 | |
| 96 | 377.5 ± 16.7 * | 680.3 ± 14.2 | 1.3 ± 0.37 * | 26.52 ± 1.25 | 2.8 ± 0.25 * | 24.2 ± 2.83 | |
| Treatments (mg/L) | Times (h) | Amount of Estrogens | |||||
|---|---|---|---|---|---|---|---|
| Residual (μg·L−1) | Extracellular (10−8 µg·cell−1) | Intracellular (10−8 µg·cell−1) | |||||
| E2 | DES | E2 | DES | E2 | DES | ||
| 0.1 | 24 | 48.4 ± 1.7 | 46.7 ± 3.3 | 0.28 ± 0.05 * | 0.48 ± 0.09 | 0.22 ± 0.09 | 0.30 ± 0.04 |
| 48 | 38.2 ± 2.1 | 36.7 ± 2.2 | 0.28 ± 0.02 * | 0.91 ± 0.03 | 0.22 ± 0.05 * | 0.55 ± 0.07 | |
| 96 | 11.2 ± 1.3 | 12.6 ± 0.7 | 0.28 ± 0.03 * | 0.31 ± 0.08 | ND * | 0.25 ± 0.05 | |
| 0.5 | 24 | 237.8 ± 6.4 * | 255.9 ± 18.5 | 1.08 ± 0.15 * | 5.3 ± 0.71 | 1.61 ± 0.37 | 1.81 ± 0.15 |
| 48 | 148.2 ± 21.3 * | 182.5 ± 11.3 | 1.04 ± 0.2 * | 3.33 ± 0.15 | 1.83 ± 0.37 * | 5.11 ± 0.41 | |
| 96 | 83.7 ± 6.5 * | 137.1 ± 15.5 | 0.21 ± 0.08 * | 5.65 ± 0.88 | 1.5 ± 0.09 * | 6.1 ± 0.29 | |
| 1.5 | 24 | 711.5 ± 15.4 * | 931.7 ± 13.3 | 3.33 ± 8.1 * | 11.5 ± 2.1 | 6.51 ± 1.5 * | 18.7 ± 3.8 |
| 48 | 477.2 ± 20.5 * | 822.3 ± 19.3 | 6.54 ± 0.66 * | 15.2 ± 0.33 | 3.83 ± 0.85 * | 11.1 ± 0.35 | |
| 96 | 233.9 ± 25.2 * | 724.5 ± 13.3 | 2.5 ± 0.57 * | 28.54 ± 2.55 | 3.8 ± 0.13 * | 21.7 ± 3.65 | |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, W.; Chen, Q.; He, N.; Sun, K.; Sun, D.; Wu, X.; Duan, S. Removal and Biodegradation of 17β-Estradiol and Diethylstilbestrol by the Freshwater Microalgae Raphidocelis subcapitata. Int. J. Environ. Res. Public Health 2018, 15, 452. https://doi.org/10.3390/ijerph15030452
Liu W, Chen Q, He N, Sun K, Sun D, Wu X, Duan S. Removal and Biodegradation of 17β-Estradiol and Diethylstilbestrol by the Freshwater Microalgae Raphidocelis subcapitata. International Journal of Environmental Research and Public Health. 2018; 15(3):452. https://doi.org/10.3390/ijerph15030452
Chicago/Turabian StyleLiu, Weijie, Qi Chen, Ning He, Kaifeng Sun, Dong Sun, Xiaoqing Wu, and Shunshan Duan. 2018. "Removal and Biodegradation of 17β-Estradiol and Diethylstilbestrol by the Freshwater Microalgae Raphidocelis subcapitata" International Journal of Environmental Research and Public Health 15, no. 3: 452. https://doi.org/10.3390/ijerph15030452
APA StyleLiu, W., Chen, Q., He, N., Sun, K., Sun, D., Wu, X., & Duan, S. (2018). Removal and Biodegradation of 17β-Estradiol and Diethylstilbestrol by the Freshwater Microalgae Raphidocelis subcapitata. International Journal of Environmental Research and Public Health, 15(3), 452. https://doi.org/10.3390/ijerph15030452