1. Introduction
Commensal rodents are known to cause substantial pre- and postharvest losses. It is estimated that rodents contribute to 5% to 10% of the losses to rice production in Asia [
1]. Besides causing direct loss to stored food, rodents also cause indirect loss: Their gnawing makes stored produce more prone to insect or fungal attacks and they contaminate a large percentage of produce with their droppings, urine, and saliva, which could possibly harbour pathogens [
2,
3,
4,
5,
6]. A review by Meerburg et al. [
3] points out the links between food security and rodents as rodents are potential reservoir hosts for over 60 zoonotic pathogens [
7,
8,
9]. Asia is predisposed for (infectious) disease emergence [
10] and there are numerous infection pathways of rodents with viruses, bacteria, and protozoans in Asia [
11,
12,
13,
14,
15]. However, there is a limited number of studies available on the prevalence of rodent-borne diseases in many regions of Asia. This raises the need to determine pathogen prevalence, especially at locations where rodents come in close contact with humans or their stored food.
We studied a specific rodent-borne zoonotic pathogen,
Leptospira, which is known to cause high disease burdens in Asia. This emerging spirochaetal bacteria occurs around rice agro-ecosystems with serious impacts on human health [
1]. South-East Asia is mentioned as the most significant centre of the disease [
16]. Studies led by the World Health Organization (WHO) on the global burden of human leptospirosis estimated more than 1 million severe cases with over 60,000 deaths annually [
17,
18,
19,
20,
21]. Leptospirosis alone affects rural communities in most countries of Asia negatively, an endemic area for leptospirosis [
16,
22,
23,
24,
25,
26]. For example, in the rural areas of Bangladesh, there are innumerable ponds and shallow waters which facilitate the survival and transmission of the
Leptospira to both maintenance hosts as well as dead-end hosts, such as humans.
In Thailand, the cases of human leptospirosis markedly increased over the 1995 to 2000 period. In 2000, leptospirosis was associated with 320 deaths reported among rice farmers [
27]. This line is also seen in Malaysia, where the number of reported cases has multiplied over 14 times between 2004 (248 cases reported) and 2012 (3604 cases reported) [
28]. This is even likely to be an underestimation because of the lack of awareness of leptospirosis symptoms due to the wide variety of these [
29,
30,
31]. Moreover, it is expected that the global disease burden will increase due to climatic change in combination with population growth, the expansion of urban areas, and floods [
3,
32,
33,
34,
35,
36].
Leptospira is classified into 22 species, encompassing over 300 serovars [
37,
38], and in Bangladesh, at least 12 serovars have been observed [
39]. Leptospirosis is maintained through chronic infection in the renal tubules of reservoir hosts, which shed
Leptospira in their urine. The majority of mammalian species are natural hosts of pathogenic leptospires. Especially, small mammals can transmit infection directly or via contaminated water and food to domestic (farm) animals and humans [
40,
41,
42]. Almost every rodent species may carry and excrete leptospires [
43]. Rodents are thought to be the most important reservoir host for a variety of serovars, and serovar prevalence varies between rodent species [
40]. Rats serve mostly as reservoir of the serovars, Icterohaemorrhagiae and Copenhageni, whereas serovars of the Ballum serogroup can be found in house mice (
Mus musculus) [
40,
42,
44,
45].
Leptospira serovars usually do not cause disease in reservoir hosts, but do cause disease in the dead-end host, which in this case is the human [
46]. Humans can acquire infection by contact with infected animals, animal tissue, animal excretions, or by contact of abrasions, cuts in the skin, or conjunctiva with contaminated water [
47]. Leptospirosis causes feverish illness in humans, and when severe it can result in Weil’s disease [
43]. Weil’s disease is characterized by jaundice, acute renal failure, and bleeding [
48], and is often mistaken for several diagnoses [
40,
49,
50,
51].
An effective strategy to minimize infection is to limit contact between humans and commensal reservoir hosts. In Asian food production systems and storage, however, the risk of contact is almost unavoidable. Thus, it is essential to monitor infection prevalence of the reservoir, in order to target control of the reservoir host at times and places where the risk is highest [
52]. Therefore, we aimed to gain insight in the prevalence of leptospirosis occurring in and potentially transmitted by rodents in Bangladesh.
Granaries offer a rich feed source for rodents, as well as suitable circumstances for the survival of leptospires. As several
Leptospira reservoir hosts (e.g., rodents) live in the same locations as people work and produce food, granaries harbour a potential epidemiological niche for pathogen transmission to humans [
53]. Information on leptospirosis and its consequences is extremely limited in many regions in Asia, amongst which is Bangladesh [
27]. To our knowledge, this study is the first one conducted in Bangladesh on the infection of rodents with
Leptospira spp. This knowledge gap results in a lack of precautions taken when handling commensal rodents or when preparing and consuming potential contaminated food [
27], which requires actual prevalence rates of
Leptospira in rodents living in and around food storages. The objective of this study was to assess the presence and infection rate of pathogenic
Leptospira in commensal rodent species in rural Bangladesh.
4. Discussion
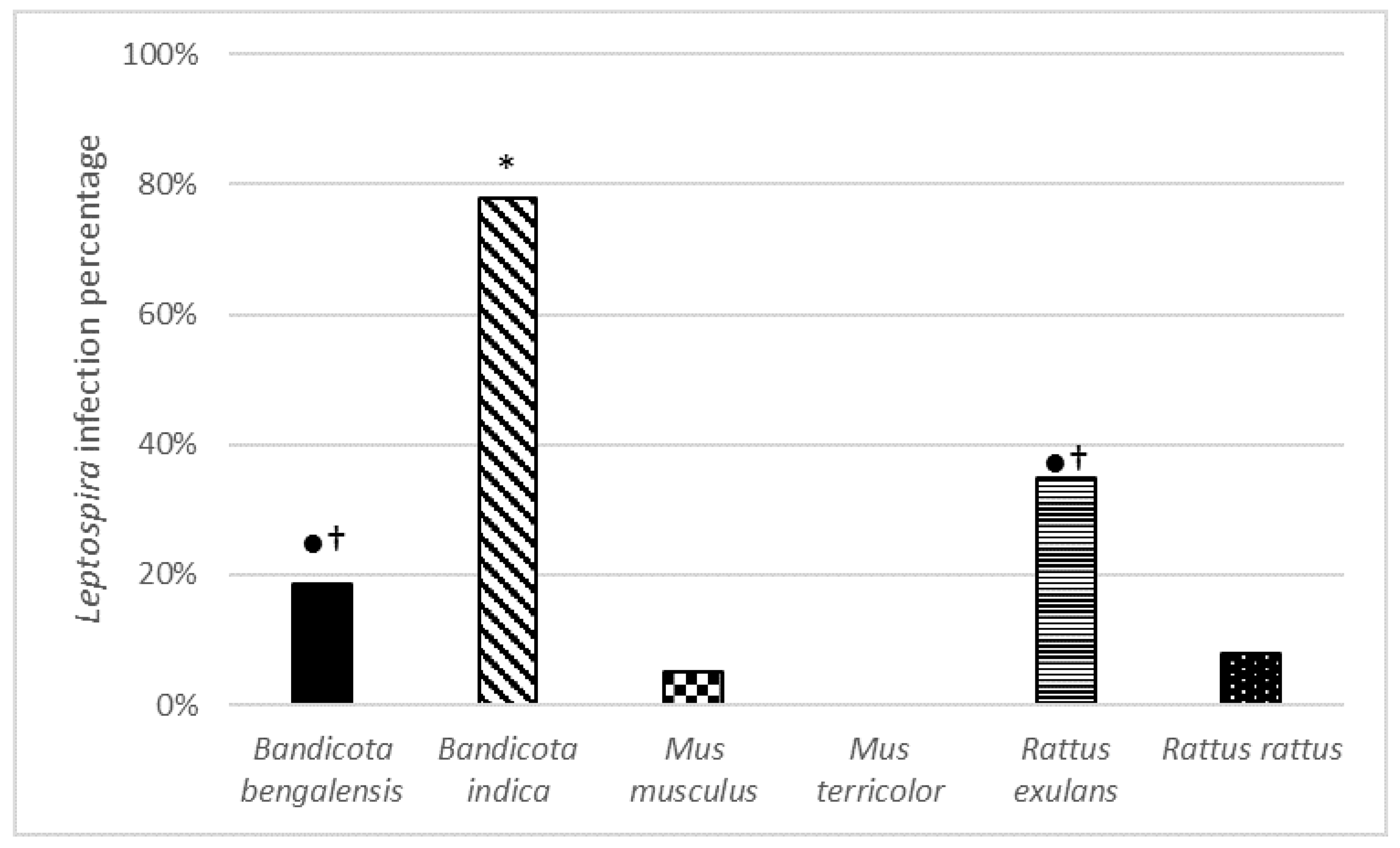
Pathogenic L. interrogans, L. borgpetersenii, and L. kirschneri species were identified in the rural Bangladesh rodent population. Almost 1 out of every 7 (61/465) rodents trapped had Leptospira bacteria in their kidneys and where thus potentially capable of shedding leptospires in and around food storage. In our study, no damage or anomalies were found during the dissections, which confirms the role of the animals as a natural reservoir. These results indicate that the risk of human exposure to pathogenic Leptospira is likely to be substantial for the workers of the Rice Milling Station of Comilla, and also for local people, since people can acquire leptospirosis via direct or indirect contact with the urine of an infected host, which in this case can also lead to a risk for the consumers of rice. However, although handling the rice may be dangerous, the risk of contracting leptospirosis via food consumption is limited if the rice is properly cooked.
The prevalence of leptospirosis in humans in Thailand, Malaysia, and India has been reported for some areas with infection rates between 15% and 35% [
55,
56,
57,
58,
59,
60,
61,
62,
63,
64]. Despite the occurrence of leptospirosis in South-East Asia, only a few studies have been performed on
Leptospira prevalence in rodents, and to our knowledge, no studies from Bangladesh have been published before. Moreover, there are only scarce and frequently dated reports about the epidemiology of leptospirosis among citizens in Bangladesh. Research from 1994 showed a human seroprevalence of 38% (
n = 89) in a rural district of Bangladesh close to rivers that regularly flood [
39]. However, no link with risk factors (such as rodents) was made. In 2001, febrile patients (
n = 1297) from two hospitals in Dhaka, Bangladesh were tested on leptospirosis and 63 patients (4.8%) were found to be positive [
23]. More recent research from Bangladesh showed that over 13% of febrile people (
n = 584) were serologically positive for
Leptospira organisms [
65]. One study on the prevalence of
Leptospira in Bangladesh looked at dairy cows in Chittagong and showed that almost 50% of the samples were positive for
Leptospira organisms [
66], which poses a high infection risk to people working on cattle farms.
A study from Cambodia (2012) on
Leptospira in rodents showed an overall infection of 11.1% (
n = 642) [
67] and a study from Malaysia found 11.0% (
n = 357) [
68], which correlates with our findings (13.1%), although we did not use a serological assay but a molecular assay. A serological study from Vietnam showed that 22% of trapped rodents host
L. interrogans and all rodents were trapped in urban areas close to the South-China Sea and in Hanoi City, a city along the Red River [
69]. Research from 2003 conducted on the Andaman Islands showed a seroprevalence of 7.1% in
R. rattus (
n = 85) [
70], which again is in line with our findings (7.9%). Research on rodents from a suburban area in India showed 14.3% of the trapped
R. rattus (
n = 28), and 16.1% of the
B. bengalensis (
n = 58) were serological positive for
Leptospira [
71]. The findings specifically on
R. rattus infections in India by Saravanan et al. were higher than the 7.9% found in the
R. rattus (
n = 191) from the current study in Bangladesh. This difference could be caused by the sensitivity and specificity of the used testing methods (serology vs. molecular diagnostic), or differences in the environment, to the ratio in which rodent species occurred and possibly also to the trapping easiness (or shyness) of each species. Meta-studies on
Leptospira prevalence in rodents in South-East Asia (
Table 5) showed that
B. bengalensis, B. indica, B. savilei, and
R. exulans were infected with
Leptospira, whereas investigated species of the
Mus genus appeared to be uninfected [
60,
61,
72,
73]. In contrast, we found a
Leptospira prevalence of 5.5% in trapped
M. musculus.
For
B. bengalensis and
R. rattus, the ratio of males and females trapped was almost evenly distributed (≈1:1). For
R. exulans (although we trapped only 23 animals of this species), more males than females were trapped (18 out of 23). For the other three species from this study,
B. indica, M. musculus, and
M. terricolor, more female specimens were collected. Research in 2006 from Pakistan found 40% of the trapped
B. bengalensis to be female (
n = 167). This male:female ratio can, in comparison to our results, be explained due to the fact that rodents trapped in this study in Bangladesh were trapped at indoor locations only, whilst in the study of Rana et al. [
74] they were trapped over multiple crop fields, which reflects the difference between male and female behaviour more. Males are more active in their explorations and depredation on food crops, whereas females may be more (trap) shy or are more concealed, or were retracted prior to breeding periods [
74]. Moreover, in our study, there was no correlation between
Leptospira prevalence and the gender of the animals for five of the six rodent species, which was also demonstrated in other earlier studies [
75,
76]. Some studies have shown that in Norway rats (
R. rattus), both in South-East Asia and in the United States of America, male specimens have a higher infection rate than females [
77,
78,
79]. However, in the current study, it was found that female
R. rattus were significantly more prone to a
Leptospira infection than males. Apparently, in our situation, the habitat’s use of rats creates a mechanism where both sexes were more or less equally exposed to infection.
When looking at the results from the rodent population from our study sites, it showed
R. exulans (
n = 23) were present in lower numbers compared to both
B. bengalensis (
n = 140) and
R. rattus (
n = 191), while the population of
R. exulans consisted mainly of males. This raises the question of whether
R. exulans competes with one of the other rodent species present. It is not unusual that species compete with each other. An early report from Bombay and Calcutta (1966) showed that
B. bengalensis increased in population size enormously and displaced the
R. rattus in urban areas [
80]. One of the underlying mechanisms was the high reproductive capacity of
B. bengalensis [
81,
82,
83], and the aggressive behaviour that dominant males exhibit [
82,
83]. Other research from India reported
B. bengalensis to be the most aggressive field rodent [
84] and that the females confine most of their time to hoarding food and are less active than males [
83]. Furthermore, although bandicoots are generally nocturnal, the
B. bengalensis is known to also become active during daytime when conditions are undisturbed [
83]. This could be advantageous in comparison to other rodent species. Unfortunately, almost no scientific studies are published on the competition of
B. bengalensis with other rodent species. We found only one report from India that claimed that female
R. rattus are submissive to
B. bengalensis [
85]. Sridhara et al. [
85] postulate that there is less aggression in closely related rodent species (e.g., within species from the
Rattus genus) compared to the violent interaction of rodents more distantly related from each other (e.g.,
Rattus genus vs.
Bandicota genus). Unfortunately, rodents of the
Rattus exulans species were not researched. This finding might explain the higher number of
Bandicota specimens and the higher
Leptospira infection rate compared to
Rattus specimens trapped in our study.
R. rattus is amongst the most omnipresent rodents in the world, and has a strong potential to displace other (native) rodent species [
86,
87]. In Australian ecosystems, it was shown that the invasive
R. rattus was dominant over the native
R. fuscipes [
88,
89]. On Madagascar,
R. rattus competes for resources with the native rodents and replaces native rodent species [
90]. Because
B. bengalensis and
R. rattus are larger than
R. exulans [
91], we expect
B. bengalensis and
R. rattus to be superior to
R. exulans. In New Caledonia, Perez et al. [
92] most frequently trapped
R. Rattus specimens (>60%,
n = 140) and very rarely trapped
R. exulans rodents (<5%,
n = 11), which supports our hypothesis that
R. rattus is dominant over
R. exulans.
In Bangladesh,
Leptospira prevalence (in %) in rodents was significantly higher in the dry season than in the rainy season. These findings are not in line with the findings from Malaysia and also from Cambodia, where rodents showed a lower infection rate in the dry (6.3%) than in the wet (26.7%) season [
67,
68]. In a study on febrile patients (
n = 1297) from Dhaka, a peak in the occurrence of
Leptospira in patients was found in October and November, shortly after the monsoon season in Bangladesh [
23]. These findings confirm the relation between floods or excessive rainfall and Leptospirosis outbreaks [
21,
32,
70]. Besides this seasonal influence on human infection, the risk of
Leptospira infection also depends on the geographic location, as well as on other risk factors, such as the risk of flooding, contaminated surface waters, and proximity to rubbish dumps (attractive for rodents) [
93,
94,
95]. Easterbrook et al. [
77] stated that seasonal fluctuations in
Leptospira infections in rodents do not occur due to the fact that once infected, the antibodies remain in the animal and the animal will test positive. This can explain our results and findings from other countries in South-East Asia, which show that rodent species living in households have a stable infection level, regardless of the geography and season [
67,
78].
Our study has some limitations. The extracted DNA from the kidneys was diluted before being added to the PCR mix in order to obtain better results by reducing potential inhibitors present in the samples. Inhibitors reduce the activity of the DNA polymerase enzyme and as a result a false negative result will be observed. As pathogenic
Leptospira colonize the kidney of the rodent, using this tissue as a source for leptospires DNA is critical; however, very high concentrations of host DNA are present. The abundant DNA of the host in the PCR reaction could lead to false-positive results as SYBR green dye is able to bind to any double-stranded DNA present in a very high concentration, thus rodent DNA can also be bound. By sequencing all samples, it was possible to filter out all false positive qPCR results. Furthermore, the effect of potential residual inhibitors in the samples could possibly result in false negative qPCR results. Because of this, we consider the 13.1% prevalence to be a prudent estimate of the actual prevalence in the rodent population in the study areas in Comilla. However, the asset of this study is that molecular diagnostics were used instead, which gives an indication of the carriership of the animal. Serologic methods used by most of the other studies conducted on
Leptospira in South-East Asia indicate that the animal did have contact with leptospires, but it remains unclear whether the animal is still a possible carrier and therefore a potential reservoir [
27,
60].
We found
L. interrogans as well as
L. borgpetersenii DNA in the samples from all five rodent species that tested positive (
B. bengalensis,
B. indica,
M. musculus,
R. rattus,
R. exulans). Other studies confirm the relation between
R. exulans and
L. interrogans [
78,
92], and in Thailand,
R. exulans was also found to be infected with both
L. interrogans and
L. borgpetersenii [
60,
96]. These findings are in line with the fact that
Leptospira species,
borgpetersenii and
interrogans, contribute a great deal to human disease in Asia [
49,
61,
78,
79]. Also, in Europe,
L. borgpetersenii and
L. interrogans are the most observed
Leptospira genomospecies present in rodents; however, in Europe, a third genomospecies is also commonly found in rodents:
L. kirschneri [
97,
98]. It is interesting to note that we also found one rodent sample infected with
L. kirschneri from a
R. exulans sample. Our findings are the first to confirm the presence of
L. kirschneri in the rodent species,
R. exulans, in Bangladesh.
Bandicota is an Asian genus of rodents, consisting of three species:
B. bengalensis,
B. indica, and
B. savilei [
103,
104,
105,
106,
107]. The few reports on
Leptospira prevalence in the
Bandicota genus indicate that all three species are potential carriers of the same leptospira species (
L. interrogans, L. borgpetersenii, L. weilli, L. inadai). Our study found that the probability of an infection with
Leptospira was significantly higher for rodents of the
Bandicota genus. Thus,
Bandicota rats could be an important host in the epidemiology of leptospirosis in Comilla. Previous studies on urban rodents identify
R. rattus to be the main reservoir host for human pathogenic
Leptospira [
94,
108,
109]. However, due to the limited information available, it is not possible to link the strain or serovar infection to a specific host species. This is unfortunate, as such information could give insight into a possible co-evolution of serovars with specific rodent species. Other studies from countries with a similar climate and cropping season as Bangladesh have used mostly serological and culturing methods, and unfortunately no specification is made into specific serogroups for each rodent species. Therefore, the only comparison possible is to see which strains are found and if this correlates with the findings in Bangladesh (
Table 5).
Cosson et al. [
78] postulate that
Leptospira species show an ecological niche; they found
L. borgpetersenii to be more abundant in rodents from dry habitats (non-floodable lands) than
L. interrogans, which implies that the infection of rodents can be linked to ecology. The
B. indica (
n = 172) is more common in the field (74%) than in or around houses [
110,
111], and in Vietnam, the
B. bengalensis was found only in grass habitats (1969). The fact that the
Bandicota species from Bangladesh were found to be infected with
L. interrogans (
Table 5) is thus in line with expectations.
Our study provides new data on rodent species as carriers of pathogenic
Leptospira in South-East Asia. From our results and the literature research (
Table 5), we can state that
L. interrogans and
L. borgpetersenii are the most common species found in rodents in South-East Asia. However, to find out whether specific strains/serovars adapt to specific reservoir hosts in specific habitats, more in-depth research with different diagnostics needs to be conducted. Although our results confirmed the importance of
Bandicota spp. and
Rattus spp. as hosts of leptospires for human health and our findings indicate that human exposure to pathogenic
Leptospira may be considerable in Comilla, the impact of leptospires on human health continues to be under recognised. In many Asian human populations, including populations in Bangladesh, the burden of undifferentiated feverish illness is substantial [
26]. One way to minimise this problem of recognising the disease is to conduct broader diagnostic tests to determine the cause of these illnesses and to inform people on the preventive measures they can take to prevent leptospirosis. Our findings highlight the complex multi-host epidemiology of leptospirosis and the importance of considering the role of rodents, and other animal hosts in the maintenance and transmission of infection when evaluating human risks. One of the key actions to minimise the public health impacts of leptospirosis in Bangladesh is to improve rodent management. A key question is the percentage to which the rodent population should be reduced to and which species should be diminished to prevent infection. In any case, preventive measures should be taken for rodent control, such as preventing rodents from accessing domestic areas and food storage to prevent pathogen transmission to humans. Rodents could be denied access to food and drinking water by taking rodent-proofing measures to existing buildings or by constructing rodent-proof warehouses. Furthermore, rodents could be discouraged from visiting domestic areas by keeping the environments clean, removing potential nesting sites, and by installing adequate sanitation and waste disposal.
With more knowledge about the rodent species present, leptospirosis, and its consequences, local people can be informed about the need for better rodent management practices. This could lead to a reduction of the impact of rodent-borne zoonotic disease in Bangladesh.

 ,
, 

{kind=link}
{kind=link}
{kind=link}

