Antifungal Properties of Two Volatile Organic Compounds on Barley Pathogens and Introduction to Their Mechanism of Action

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fungal Strains
2.2. Evaluation of the Effect of Five VOCs in Gas Phase on F. culmorum and C. sativus Growth Without Direct Contact
2.3. Evaluation of the Effect of Methylprop-2-Enoate and Methylpropanoate on F. culmorum and C. sativus Growth in Direct Contact
2.4. Evaluation of the Fungicidal/Fungistatic Effect of the Most Efficient VOCs on C. sativus and F. culmorum Growth
2.5. Evaluation of the Release of K+ Ions in the Extracellular Medium
2.6. Data Analysis
3. Results
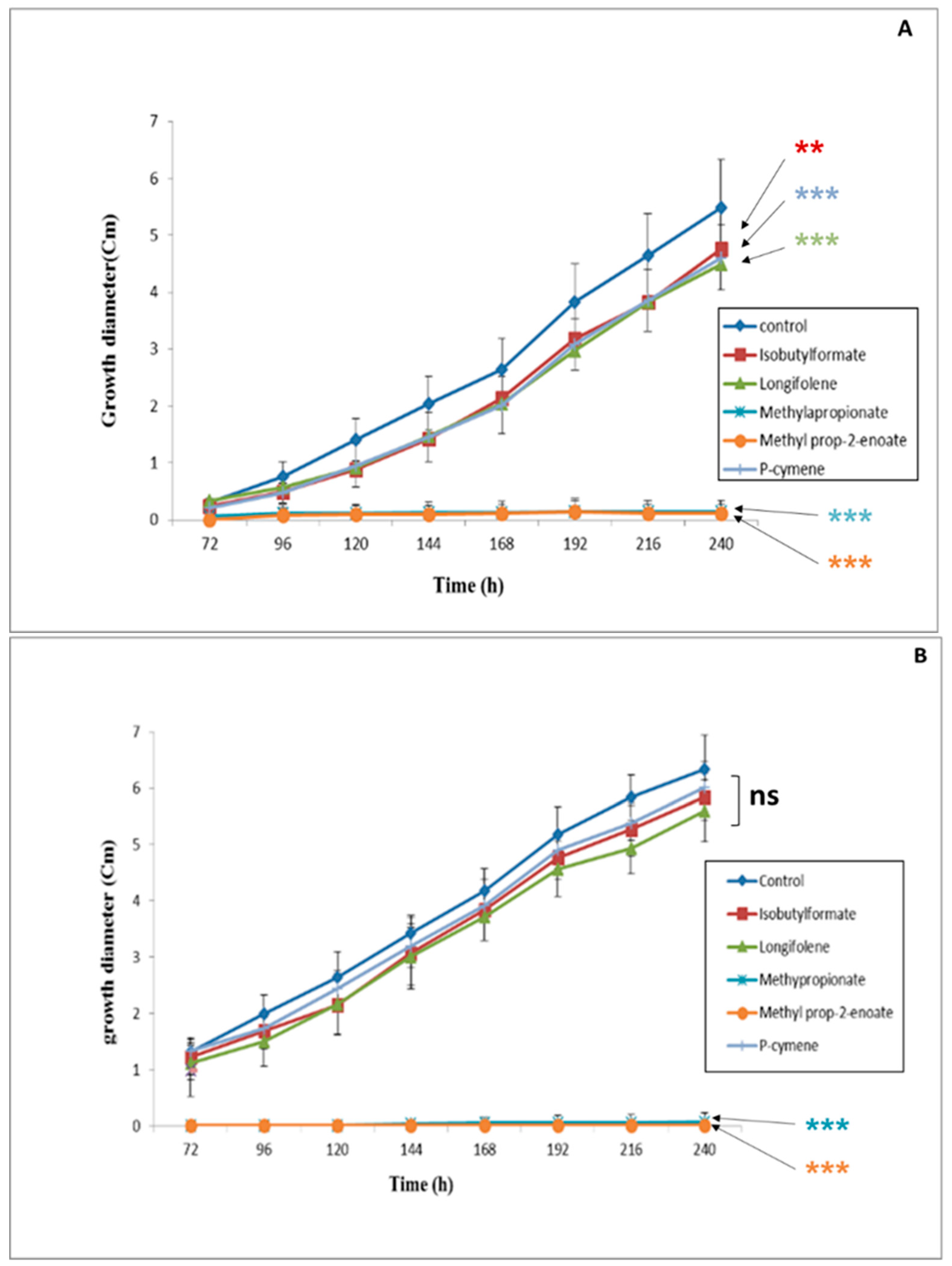
3.1. Evaluation of the Effect of Five VOCs on F. culmorum and C. sativus Growth
3.2. Evaluation of the Inhibitory Effect of the Most Efficient VOCs on Fungal Growth in Direct and Indirect Contact
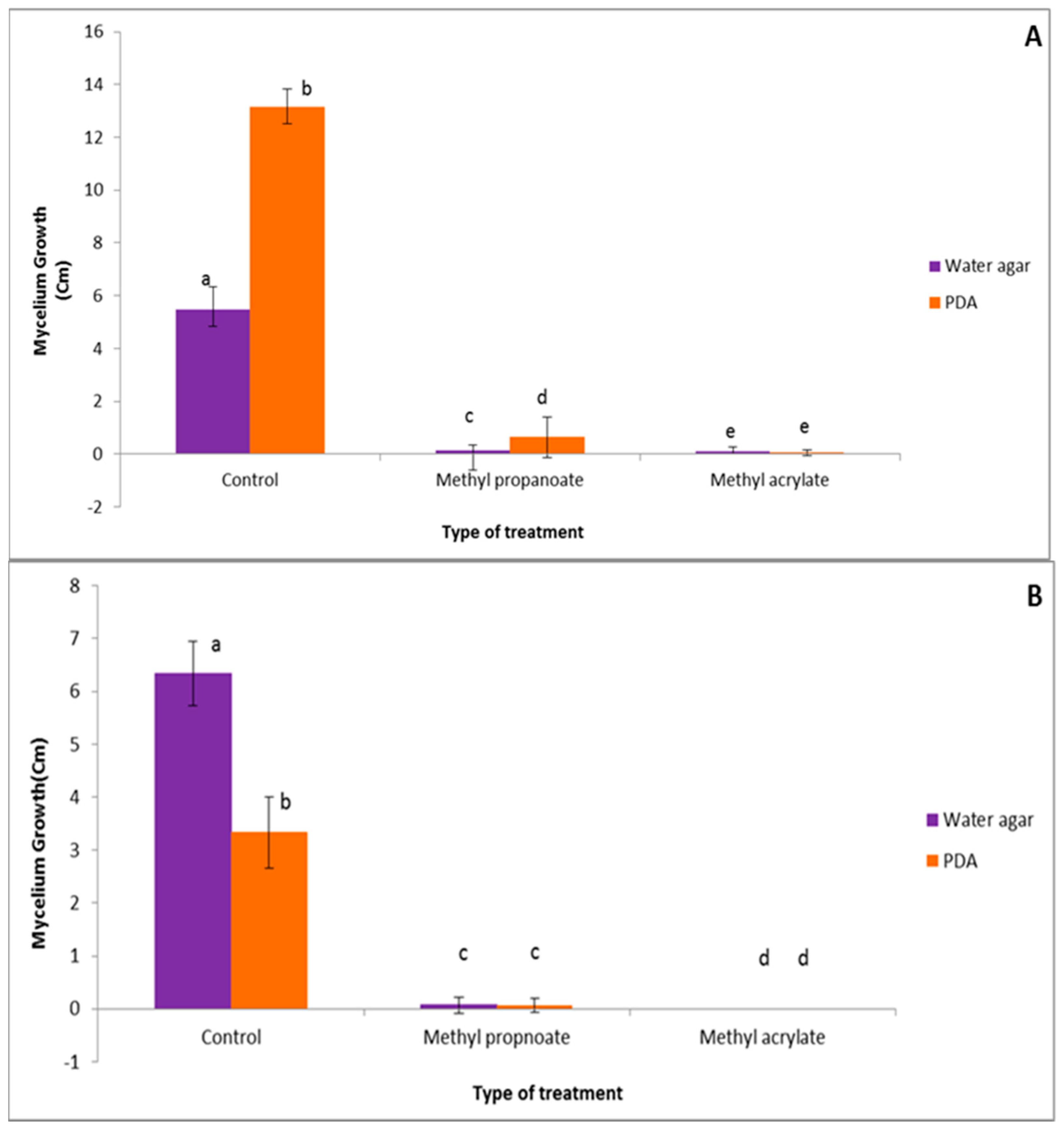
3.2.1. Effect of Methyl Prop-2-Enoate and Methyl Propanoate in Direct Contact
3.2.2. Effect of Methyl Prop-2-Enoate and Methyl Propanoate in Indirect Contact
3.3. Effect of the VOC Concentrations on VOC Antifungal Ability
3.4. Evaluation of the Release of K+ Ions into the Extracellular Medium
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

References
- Karov, I.; Mitrev, S.; Arsov, E. Bipolaris sorokiniana (Teleomorph cochliobolus SATIVUS), causer of barley leaf lessions and root rot in Macedonia. Proc. Nat. Sci. Matica Srpska Novi Sad 2009, 116, 167–174. [Google Scholar] [CrossRef]
- Kirkegaard, J.A.; Simpfendorfer, S.; Holland, J.; Bambach, R.; Moore, K.J.; Rebetzke, G.J. Effect of previous crops on crown rot and yield of durum and bread wheat in northern Nsw. Aust. J. Agric. Res. 2004, 55, 321–334. [Google Scholar] [CrossRef]
- Summerell, B.A.; Leslie, J.F.; Backhouse, D.; Bryden, W.L.; Burgess, L.W. Fusarium: Paul, E. Nelson Memorial Symposium; APS Press: St. Paul, MN, USA, 2001. [Google Scholar]
- Lamprecht, S.C.; Marasas, W.F.O.; Hardy, M.B.; Calitz, F.J. Effect of crop rotation on crown rot and the incidence of Fusarium pseudograminearum in wheat in the western cape, South Africa. Australas. Plant. Pathol. 2006, 35, 419–426. [Google Scholar] [CrossRef]
- Evans, M.L.; Hollaway, G.J.; Dennis, J.I.; Correll, R.; Wallwork, H. Crop sequence as a tool for managing populations of Fusarium pseudograminearum and F. Culmorum in South-Eastern Australia. Australas. Plant. Pathol. 2010, 39, 376–382. [Google Scholar] [CrossRef]
- Bateman, G.L.; Kwaśna, H. Effects of number of winter wheat crops grown successively on fungal communities on wheat roots. Appl. Soil Ecol. 1999, 13, 271–282. [Google Scholar] [CrossRef]
- Bateman, G.L.; Murray, G. Seasonal variations in populations of fusarium species in wheat-field soil. Appl. Soil Ecol. 2001, 18, 117–128. [Google Scholar] [CrossRef]
- Sturz, A.V.; Bernier, C.C. Influence of crop rotations on winter wheat growth and yield in relation to the dynamics of pathogenic crown and root rot fungal complexes. Can. J. Plant. Pathol. 1989, 11, 114–121. [Google Scholar] [CrossRef]
- Felton, W.L.; Marcellos, H.; Alston, C.; Martin, R.J.; Backhouse, D.; Burgess, L.W.; Herridge, D.F. Chickpea in wheat-based cropping systems of Northern New South Wales. II. influence on biomass, grain yield, and crown rot in the following wheat crop. Aust. J. Agric. Res. 1998, 49, 401–408. [Google Scholar] [CrossRef]
- Kaddes, A.; Parisi, O.; Berhal, C.; Ben Kaab, S.; Fauconnier, M.L.; Nasraoui, B.; De Clerck, C. Evaluation of the effect of two volatile organic compounds on barley pathogens. Molecules 2016, 21, 1124. [Google Scholar] [CrossRef]
- Dudareva, N.; Negre, F.; Nagegowda, D.A.; Orlova, I. Plant volatiles: Recent advances and future perspectives. Crit. Rev. Plant Sci. 2006, 25, 417–440. [Google Scholar] [CrossRef]
- Morath, S.U.; Hung, R.; Bennett, J.W. Fungal volatile organic compounds: A review with emphasis on their biotechnological potential. Fungal Biol. Rev. 2012, 26, 73–83. [Google Scholar] [CrossRef]
- Fiers, M.; Lognay, G.; Fauconnier, M.L.; Jijakli, M.H. Volatile compound-mediated interactions between barley and pathogenic fungi in the soil. PLoS ONE 2013, 8, e66805. [Google Scholar] [CrossRef]
- Kaddes, A.; Fauconnier, M.L.; Sassi, K.; Nasraoui, B.; Jijakli, M.H. Endophytic fungal volatile compounds as solution for sustainable agriculture. Molecules 2019, 24, 1065. [Google Scholar] [CrossRef]
- Bryman, A.; Duncan, C. Quantitative Data Analysis with Minitab: A Guide for Social Scientists; Psychology Press: London, UK, 1996. [Google Scholar]
- Dunnett, C.W. New tables for multiple comparisons with a control. Biometrics 1964, 20, 482–491. [Google Scholar] [CrossRef]
- Sewake, K.T.; Uchida, J.Y. Diseases of Heliconia in Hawaii. Associate county extension agent and associate professor of plant pathology respectively, CTAHR, University of Hawaii. Human Res. 1991, 52, 5R47. [Google Scholar]
- Meddah, N.; Touhami, A.O.; Benkirane, R.; Douira, A. Pouvoir pathogène de Cochliobolus australiensis et Cochliobolus spicifer vis-à-vis de Musa accuminata, L. variété «grande naine» au Maroc. J. Anim. Plant. Sci. 2014, 21, 3263–3272. [Google Scholar]
- Arabi, M.I.E.; Jawhar, M. Pathotypes of Cochliobolus sativus (Spot blotch) on barley in Syria. J. Plant. Pathol. 2003, 85, 193–196. [Google Scholar]
- Neched, H. Étude comparative des traitements de semences sans fongicide chez les céréales À L’aide De L’ozone Et De L’oxygène pur; Université Laval: Quebec City, QC, Canada, 2015. [Google Scholar]
- Zirihi, G.N.; Soro, S.; Kone, D.; Kouadio, Y.J. Activité antifongique de l’extrait naturel de combretum sp. in vitro sur 3 espèces fongiques telluriques des cultures de tomate en Côte D’Ivoire. Rev. Ivoir. Sci. Technol. 2008, 11, 131–142. [Google Scholar]
- Sikkema, J.; de Bont, J.A.; Poolman, B. Mechanisms of membrane toxicity of hydrocarbons. Microbiol. Mol. Biol. Rev. 1995, 59, 201–222. [Google Scholar]
- Portillo, F. Regulation of plasma membrane H+−ATPase in fungi and plants. Biochim. Et Biophys. Acta (BPA)-Rev. Biomembr. 2000, 1469, 31–42. [Google Scholar] [CrossRef]
- Plumridge, A.; Hesse, S.J.; Watson, A.J.; Lowe, K.C.; Stratford, M.; Archer, D.B. The weak acid preservative sorbic acid inhibits conidial germination and mycelial growth of Aspergillus niger through intracellular acidification. Appl. Env. Microbiol. 2004, 70, 3506–3511. [Google Scholar] [CrossRef]
- Hopkins, W.G. Physiologie Végétale; De Boeck Supérieur: Quebec City, QC, Canada, 2003. [Google Scholar]
- Bartlett, D.W.; Clough, J.M.; Godwin, J.R.; Hall, A.A.; Hamer, M.; Parr-Dobrzanski, B. The strobilurin fungicides. Pest. Manag. Sci. Former. Pestic. Sci. 2002, 58, 649–662. [Google Scholar] [CrossRef]
- Kunova, A.; Pizzatti, C.; Cortesi, P. Impact of tricyclazole and azoxystrobin on growth, sporulation and secondary infection of the rice blast fungus, Magnaporthe oryzae. Pest. Manag. Sci. 2013, 69, 278–284. [Google Scholar] [CrossRef]
- Kaur, S.; Mishra, P. Dimorphism-associated changes in plasma membrane H+-ATPase activity of Candida albicans. Arch. Microbiol. 1991, 156, 412–415. [Google Scholar] [CrossRef]
- Bhatia, R.; Shreaz, S.; Khan, N.; Muralidhar, S.; Basir, S.F.; Manzoor, N.; Khan, L.A. Proton pumping ATPase mediated fungicidal activity of two essential oil components. J. Basic Microbiol. 2012, 52, 504–512. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| A | ||||
| VOCs | Fusarium culmorum | Cochliobolus sativus | ||
| MP | MA | MP | MA | |
| 500 µM | - | -- | -- | -- |
| - | -- | -- | -- | |
| - | -- | -- | -- | |
| 1000 µM | - | -- | -- | -- |
| - | --- | -- | -- | |
| - | --- | -- | -- | |
| Controls | + | + | + | + |
| + | + | + | + | |
| + | + | + | + | |
| B | ||||
| VOCs | Fusarium culmorum | Cochliobolus sativus | ||
| MA | MP | MP | MA | |
| 500 µM | - | + | + | - |
| - | + | + | - | |
| - | + | + | - | |
| 1000 µM | - | + | + | - |
| - | + | + | - | |
| - | + | + | - | |
 Means “Absence of mycelium of the pathogen in vials”;
Means “Absence of mycelium of the pathogen in vials”;  Means “Presence of mycelium of the pathogen in vials”.
Means “Presence of mycelium of the pathogen in vials”.| Concentration | 0 h | 1 h | 2 h | 3 h | 4 h | 5 h |
|---|---|---|---|---|---|---|
| Control | 400 | 400 | 400 | 400 | 400 | 400 |
| MA 100 µM | 400 | 400 | 400 | 400 | 400 | 400 |
| MA 500 µM | 400 | 400 | 400 | 700 | 700 | 700 |
| MA 1000 µM | 400 | 400 | 400 | 700 | 700 | 1000 |
| MP 100 µM | 400 | 400 | 400 | 400 | 400 | 400 |
| MP 500 µM | 400 | 400 | 400 | 400 | 400 | 700 |
| MP 1000 µM | 400 | 400 | 400 | 700 | 700 | 700 |
 400 mg/L K+ present in the extracellular medium of a spore suspension of Fusarium culmorum.
400 mg/L K+ present in the extracellular medium of a spore suspension of Fusarium culmorum.  700 mg/L K+ present in the extracellular medium of a spore suspension of Fusarium culmorum.
700 mg/L K+ present in the extracellular medium of a spore suspension of Fusarium culmorum.  1000 mg/L K+ present in the extracellular medium of a spore suspension of Fusarium culmorum.
1000 mg/L K+ present in the extracellular medium of a spore suspension of Fusarium culmorum.| Concentration | 0 h | 1 h | 2 h | 3 h | 4 h | 5 h |
|---|---|---|---|---|---|---|
| Control | 400 | 400 | 400 | 400 | 400 | 400 |
| MA 100 µM | 400 | 400 | 400 | 400 | 400 | 700 |
| MA 500 µM | 400 | 400 | 400 | 400 | 1000 | 1000 |
| MA 1000 µM | 400 | 400 | 400 | 400 | 1000 | 1000 |
| MP 100 µM | 400 | 400 | 400 | 400 | 400 | 700 |
| MP 500 µM | 400 | 400 | 400 | 400 | 700 | 1000 |
| MP 1000 µM | 400 | 400 | 400 | 400 | 700 | 1000 |
400 mg/L K+ present in the extracellular medium of a spore suspension of Cochliobolus sativus. 700 mg/L K+ present in the extracellular medium of a spore suspension of Cochliobolus sativus. 1000 mg/L K+ present in the extracellular medium of a spore suspension of Cochliobolus sativus.© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kaddes, A.; Fauconnier, M.-L.; Sassi, K.; Nasraoui, B.; Jijakli, M.H. Antifungal Properties of Two Volatile Organic Compounds on Barley Pathogens and Introduction to Their Mechanism of Action. Int. J. Environ. Res. Public Health 2019, 16, 2866. https://doi.org/10.3390/ijerph16162866
Kaddes A, Fauconnier M-L, Sassi K, Nasraoui B, Jijakli MH. Antifungal Properties of Two Volatile Organic Compounds on Barley Pathogens and Introduction to Their Mechanism of Action. International Journal of Environmental Research and Public Health. 2019; 16(16):2866. https://doi.org/10.3390/ijerph16162866
Chicago/Turabian StyleKaddes, Amine, Marie-Laure Fauconnier, Khaled Sassi, Bouzid Nasraoui, and M. Haïssam Jijakli. 2019. "Antifungal Properties of Two Volatile Organic Compounds on Barley Pathogens and Introduction to Their Mechanism of Action" International Journal of Environmental Research and Public Health 16, no. 16: 2866. https://doi.org/10.3390/ijerph16162866
APA StyleKaddes, A., Fauconnier, M.-L., Sassi, K., Nasraoui, B., & Jijakli, M. H. (2019). Antifungal Properties of Two Volatile Organic Compounds on Barley Pathogens and Introduction to Their Mechanism of Action. International Journal of Environmental Research and Public Health, 16(16), 2866. https://doi.org/10.3390/ijerph16162866