Dual-process models have been consistent in their description of severe addictive disorders (such as AUD) [
13,
18,
78]. However, evidence in preclinical populations (e.g., in adolescent BDs without AUD) remains scarce. From a neurodevelopmental perspective, these models have described late adolescence as a period characterised by increased sensation seeking and reduced cognitive inhibitory control [
11]. In this research, we aimed to investigate within the framework of dual-process models the extent to which binge drinking may influence the balance between the reflective and affective-automatic systems in a sample of first-year university students. To this end, the ERPs elicited during a beverage Go/NoGo task were recorded in order to explore the degree to which alcohol-related stimuli may modulate inhibition of a prepotent response in BDs without AUD.
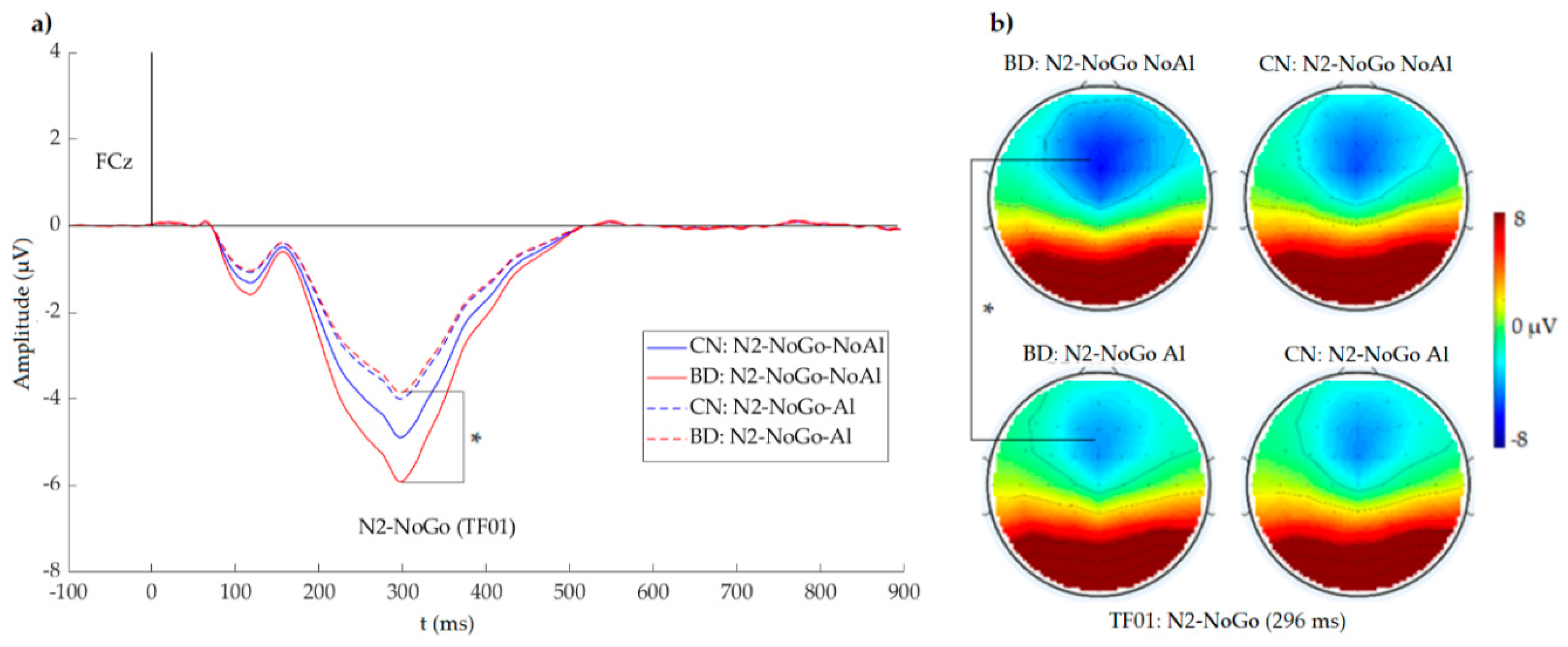
According to our first hypothesis, we expected larger amplitudes of ERP components associated with conflict monitoring and response inhibition (N2-NoGo and/or P3-NoGo) in BDs. Consistently, in our experimental task, the N2-NoGo amplitude was larger in BDs (and not in CNs) in the NoGo-NoAl condition relative to the NoGo-Al condition. Moreover, regarding P3-NoGo, a gender/sex-related effect was observed, as there were no differences between the amplitudes of this component in response to Al and NoAl stimuli in male BDs.
4.1. BDs Displayed Increased Conflict Monitoring Activity in a Context of Frequent Al Stimuli
N2-NoGo is considered an index of detection of conflict between competing responses (Go and NoGo), with neural sources located in the IFC [
36,
37] and ACC [
38,
39]. Following the conflict monitoring theory of Botvinick, Cohen, and Carter [
79], ACC acts as an online indicator of the degree of response conflict, conveying this information to other systems (such as the prefrontal cortex, including IFC) devoted more directly in the implementation of cognitive control strategies (attentional selection, response priming, goal maintenance, etc.). Consequently, N2-NoGo can be interpreted as an indicator of the previous cognitive processes required to implement inhibitory control rather than the actual inhibitory brake.
According to our N2-NoGo findings and conflict monitoring theory, it is possible that, in BDs, the Al stimuli may enhance activation of the prepotent Go pre-response so that the conflict will thus increase during NoGo trials. BDs would therefore have to expend greater cognitive effort and to recruit extra neural resources to successfully inhibit the primed Go response. In terms of the dual-process model framework, during the Go-Al vs. NoGo-NoAl condition, BDs may display overactivation of the automatic system (due to the higher motivational value of Go-Al stimuli). Extra neural resources may be allocated to the reflective system (as reflected by larger N2-NoGo amplitudes) to overcome this overactivation and to successfully resolve the conflict.
Previous neurocognitive research on binge drinking has already shown that alcohol-related content may modulate N2-NoGo amplitudes of prepotent response inhibition. As we discuss below, the direction and intensity of these modulations differ between studies, possibly due to methodological factors, such as different task designs or sample characteristics. In those studies reporting greater activation in the absence of behavioural differences, the results were interpreted within the context of the compensatory hypothesis.
In an fMRI design, Ames et al. [
51] observed greater activity in the right dorsolateral and medial prefrontal cortex, anterior cingulate cortex, and anterior insula in BDs during NoGo-Al trials of a beverage Go/NoGo task. These authors suggested that, in BDs, the increased incentive value or salience of the alcohol pictures during NoGo trials may have served as an attentional bias cue, resulting in greater effort being required to withhold a response. Although the task only included the Go-NoAl vs. NoGo-Al condition under a different design and therefore we cannot fully compare their results with ours, the increased activation also indicates compensatory activity of neural regions related to N2-NoGo (medial prefrontal cortex and ACC).
In a Go/NoGo task with geometrical shapes used as Go stimuli and a combination of other geometrical shapes, Al pictures, and NoAl pictures as NoGo stimuli, Watson et al. [
47] found that the NoAl stimuli generally elicited larger N2-NoGo than the Al stimuli. However, in those subjects who reported more episodes of intoxication in the last six months, the N2-NoGo amplitude was larger in response to Al stimuli. This difference relative to our findings may be associated with the task design, as the Go trials consisted entirely of nonbeverage, neutral stimuli so that conflict may emerge in a different way.
Task design may also explain the absence of N2-NoGo differences in a letter Go/NoGo task with pictures of alcoholic beverages and neutral pictures as a background [
48]. The fact that alcohol pictures were not explicitly related to task instructions may have led to the lack of differences, as shown by Lannoy et al. [
49] in a study in which group differences in N2-NoGo amplitudes and behavioural performance only emerged when Al pictures were explicitly related to the task instructions. Furthermore, in contrast to the compensatory hypothesis, the aforementioned research found smaller N2-NoGo amplitudes for Al pictures in BDs (relative to CNs), suggesting that BDs may experience some difficulties at the attentional level in inhibiting a prepotent response when alcohol pictures are present. In comparison with our study, the level of alcohol consumption (mean AUDIT score: 17.20 ± 5.32) in the previous sample may suggest a more advanced state of deleterious cerebral effects as a result of a higher rate of consumption. It is therefore possible that the compensatory mechanisms that may be acting in our sample may have been unavailable in the subjects of the previous research. A follow-up study of our sample would be necessary to confirm this explanation.
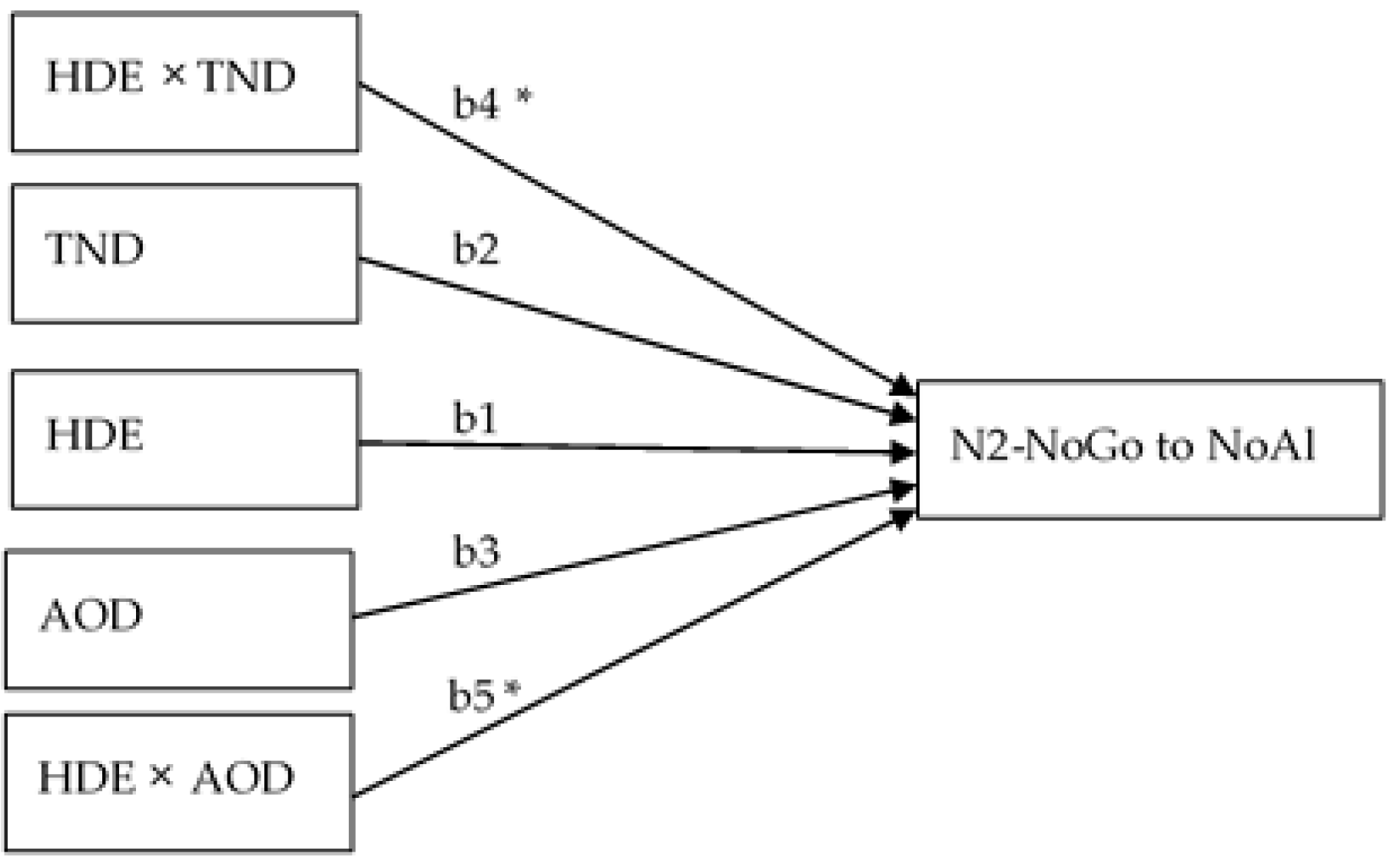
Considering our findings, the amplitude of N2-NoGo elicited in response to NoAl stimuli also appeared to be related to the rate of alcohol consumption. Correlational and regression-based moderation analyses revealed that N2-NoGo in response to NoAl was significantly correlated with the number of BDEs in the last six months and with the total number of alcoholic drinks in the last six months. These correlations showed that, in BDs, more BDEs and a greater number of drinks were related to larger amplitudes of this ERP component. Regression-based moderation analysis showed that, in this group, the amplitude of N2-NoGo in response to NoAl was predicted by the number of BDEs specifically when this variable interacted with the total amount of drinks and with the age of onset of drinking. In other words, N2-NoGo amplitudes were largest in subjects with a short history of binge alcohol consumption but with a high frequency and intensity.
4.2. Male BDs Did Not Show Differences in Evaluation/Closure of the Inhibition Process Between Alcohol and Nonalcohol Stimuli
P3-NoGo has been related to closure or the evaluation of inhibition process [
43], and its neural source has been located in IFC [
36]. Results showed a group × sex × type of stimulus interaction regarding the amplitude of this component. During a context of predominant Al stimuli, NoGo-NoAl trials elicit larger P3-NoGo, but this difference was not significant in male BDs. It seems that, in male BDs, the evaluation/closure of the inhibition process may be altered by the motivational value of the stimuli.
To our knowledge, no gender/sex-related differences in P3-NoGo have previously been reported in research on binge drinking. Gender/sex differences related to BD in young samples have been explained in terms of neurodevelopmental delay in men relative to women [
80], together with differences in alcohol consumption patterns, expectations, and effects [
81]. Nevertheless, replication of these results and longitudinal data are required to further clarify these gender/sex-related differences and how they evolve over time.
Regardless of gender/sex differences and alcohol-related content, previous electrophysiological research with neutral Go/NoGo tasks has detected P3-NoGo compensatory alterations related to binge drinking. A longitudinal study by our research group reported larger P3-NoGo amplitudes (in response to neutral stimuli) in BDs than in CNs, together with greater activation of the right inferior frontal cortex, only at the second evaluation (20–21 years old and at least two years of binge drinking) [
55]. As there are great similarities between the sample used in the aforementioned study during the first evaluation and the sample of our study, it is possible that anomalies in P3-NoGo amplitude will emerge in subsequent follow-ups of our sample if the binge drinking pattern persists.
4.3. Increased Salience of Al Stimuli.
P100 is an early ERP component that is sensitive to variations in stimulus parameters but that also can be modulated by attentional influences [
54]. Our second hypothesis posits that greater salience of Al stimuli for BDs will be reflected by larger amplitude of the P1 component. The results showed that Al stimuli elicited larger P1 amplitudes; however, no differences in relation to groups were found, except for a second-order interaction (group × type of trial × type of stimulus
). Post hoc analysis did not reveal any group-based differences in P1 amplitudes, so at least in our sample, BDs did not show increased responses to alcohol pictures at the perceptual level.
Interestingly, correlational analysis showed that an early onset of drinking was correlated with larger P1 amplitudes in the context of prepotent Al stimuli. This is consistent with previous electrophysiological research (in oddball tasks) showing increased P1 amplitudes to Al stimuli in BDs [
51,
52]. However, these findings are not consistent with those of a longitudinal study in which a reduction in P1 amplitude was observed in response to both Al and NoAl stimuli after only one year of a binge drinking pattern of consumption [
82]. This incongruence between the findings of both studies may be due to aspects of sample selection, such as the inclusion, in the longitudinal research [
82], of subjects with family history of alcohol consumption. Further replication of these results from a longitudinal approach is required.
4.4. An Integrated View of Beverage Go/NoGo Tasks Through the Cognitive Stream
From all of the above, the findings related to the modulations of response inhibition by motivational (alcohol-related) stimuli in BDs seem to indicate anomalies in ERP components at different stages of the cognitive stream, which may depend on the history and intensity of binge drinking. In early perceptual components, the scarce findings range from subtle correlations (as in our research) to larger [
52,
53] or smaller [
82] P1 amplitudes. Regarding later cognitive ERP components, larger N2-NoGo amplitudes in response to nonalcohol stimuli (as we observed) may reflect compensatory conflict monitoring processes that overcome bias to an alcohol preponderant context; however, such compensatory mechanisms may be overcome by a longer history of consumption (or more difficult task), as reflected by lower N2-NoGo or poorer behavioural execution reported in other studies [
45,
49]. The same seems to occur with the P3-NoGo component, as larger amplitudes were reported in a longitudinal research [
55] only during the second evaluation. In summary, during the first years of a binge drinking pattern of consumption, compensatory mechanisms still seem to be available in fairly simple inhibitory tasks and Al stimuli seem to affect conflict monitoring rather than inhibitory processes per se. Longitudinal data are needed to confirm whether this compensatory mechanism will no longer be available if the binge drinking pattern is maintained as well as to explore whether alcohol bias or response inhibition alterations will occur.

 ,
, 




{kind=link}
{kind=link}
{kind=link}
{kind=link}