Research and Technological Advances Regarding the Study of the Spread of Antimicrobial Resistance Genes and Antimicrobial-Resistant Bacteria Related to Animal Husbandry

 ,
, 

Abstract
:1. Introduction
2. Methods
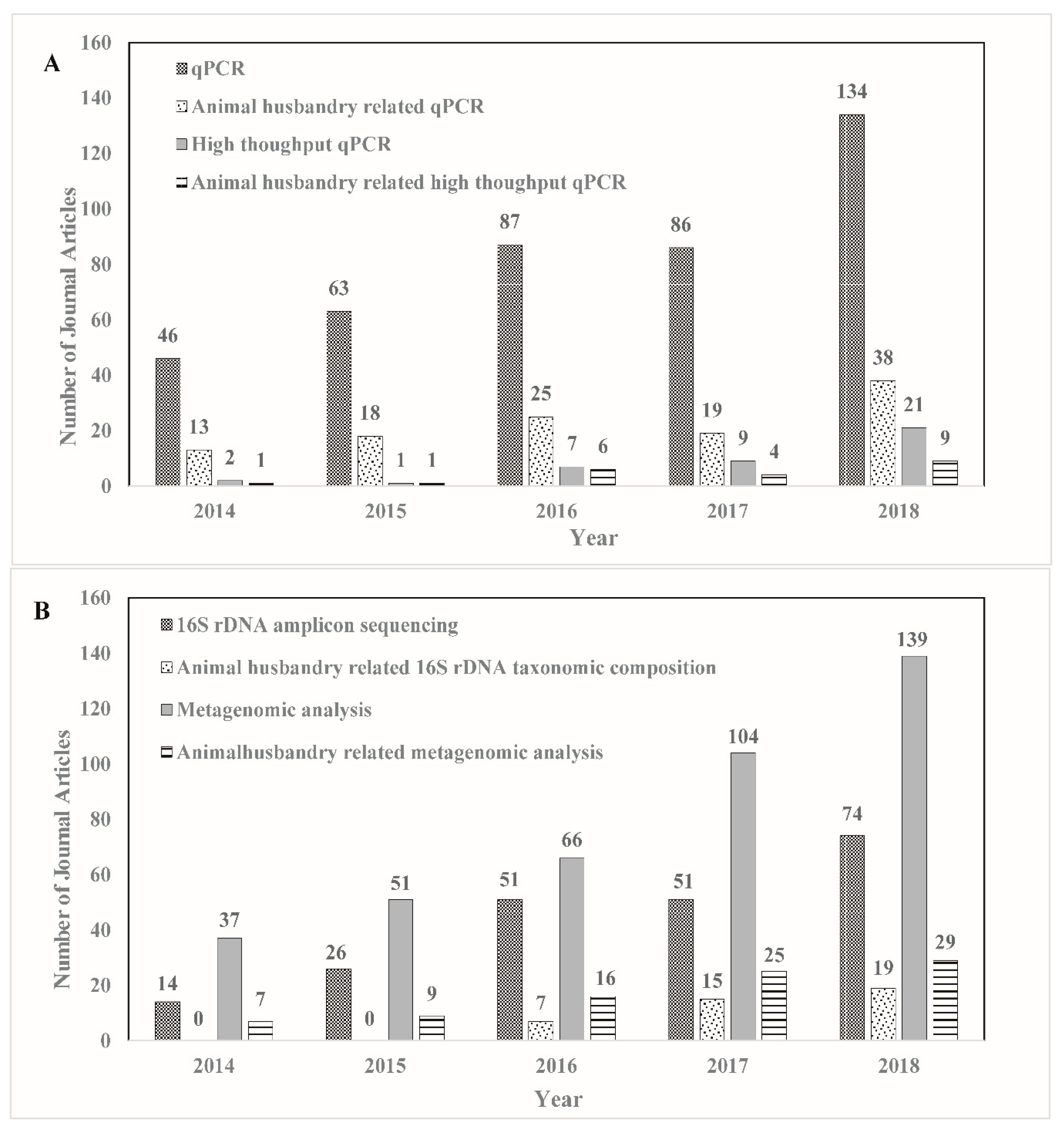
2.1. Metagenomic Analysis
2.2. Animal Husbandry Related Metagenomic Analysis
2.3. Quantitative PCR (qPCR)
2.4. Animal Husbandry Related qPCR
2.5. High-Throughput qPCR
2.6. Animal Husbandry Related High-Throughput qPCR
2.7. -16S rDNA Taxonomic Composition
2.8. Animal-Husbandry-Related 16S rDNA Taxonomic Composition
3. The State of Current Veterinary Research of ARB and ARG Pollution
3.1. Impact of Veterinary Antimicrobials on ARB and ARGs, and Their Correlation with ARGs
3.2. ARG Environmental Transfer
4. Research Methods
4.1. Bacterial Culture
4.2. Molecular Detection
4.2.1. PCR/qPCR and High-Throughput qPCR
4.2.2. Bacterial Taxonomic Composition (High-Throughput 16S rDNA Amplicon Sequencing)
4.2.3. Metagenomic Analysis
4.2.4. Metatranscriptomic Sequencing Technology
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Food and Agriculture Organization of the United Nations (FAO). 2018 Food Outlook—Biannual Report on Global Food Markets; Food and Agriculture Organization of the United Nations: Rome, Italy, 2018; p. 104. [Google Scholar]
- Robinson, T.P.; Thornton, P.K.; Franceschini, G.; Kruska, R.L.; Chiozza, F.; Notenbaert, A.; Cecchi, G.; Herrero, M.; Epprecht, M.; Fritz, S.; et al. Global Livestock Production Systems; Food and Agriculture Organization of the United Nations (FAO) and International Livestock Research Institute (ILRI): Rome, Italy, 2011; 152p. [Google Scholar]
- Wellington, E.M.H.; Boxall, A.B.A.; Cross, P.; Feil, E.J.; Gaze, W.H.; Hawkey, P.M.; Johnson-Rollings, A.S.; Jones, D.L.; Lee, N.M.; Otten, W.; et al. The role of the natural environment in the emergence of antibiotic resistance in Gram-negative bacteria. Lancet Infect. Dis. 2013, 13, 155–165. [Google Scholar] [CrossRef]
- Heuer, H.; Schmitt, H.; Smalla, K. Antibiotic resistance gene spread due to manure application on agricultural fields. Curr. Opin. Microbiol. 2011, 14, 236–243. [Google Scholar] [CrossRef] [PubMed]
- Committee for Medicinal Products for Veterinary Use (CVMP). Reflection Paper on Antimicrobial Resistance in the 4 Environment: Considerations for Current and Future Risk 5 Assessment of Veterinary Medicinal Products; European Medcines Agency-Science Medicines Health: Amsterdam, The Netherlands, 2018; pp. 1–50. [Google Scholar]
- Venglovsky, J.; Sasakova, N.; Placha, I. Pathogens and antibiotic residues in animal manures and hygienic and ecological risks related to subsequent land application. Bioresour. Technol. 2009, 100, 5386–5391. [Google Scholar] [CrossRef] [PubMed]
- Looft, T.; Joghson, T.A.; Allen, H.K.; Bayles, D.O.; Alt, D.P.; Stedtfeld, R.D.; Sul, W.J.; Stedtfeld, T.M.; Chai, B.; Cole, J.R.; et al. In-feed antibiotic effects on the swine intestinal microbiome. Proc. Natl. Acad. Sci. USA 2012, 109, 1691–1696. [Google Scholar] [CrossRef] [Green Version]
- Fang, H.; Wang, H.; Cai, L.; Yu, Y. Prevalence of antibiotic resistance genes and bacterial pathogens in long-term manured greenhouse soils as revealed by metagenomic survey. Environ. Sci. Technol. 2015, 49, 1095–1104. [Google Scholar] [CrossRef]
- Pruden, A.; Larsson, D.G.; Amezquita, A.; Collignon, P.; Brandt, K.K.; Graham, D.W.; Lazorchak, J.M.; Suzuki, S.; Silley, P.; Snape, J.R.; et al. Management options for reducing the release of antibiotics and antibiotic resistance genes to the environment. Environ. Health Perspect. 2013, 121, 878–885. [Google Scholar] [CrossRef]
- Zhang, Z.; Li, B.; Li, N.; Sardar, M.F.; Song, T.; Zhu, C.; Lv, X.; Li, H. Effects of UV disinfection on phenotypes and genotypes of antibiotic-resistant bacteria in secondary effluent from a municipal wastewater treatment plant. Water Res. 2019, 157, 546–554. [Google Scholar] [CrossRef]
- Liang, B.; Ma, J.; Cai, W.; Li, Z.; Liu, W.; Qi, M.; Zhao, Y.; Ma, X.; Deng, Y.; Wang, A.; et al. Response of chloramphenicol-reducing biocathode resistome to continuous electrical stimulation. Water Res. 2019, 148, 398–406. [Google Scholar] [CrossRef]
- Chen, Q.; An, X.; Zheng, B.X.; Ma, Y.B.; Su, J.Q. Long-term organic fertilization increased antibiotic resistome in phyllosphere of maize. Sci. Total Environ. 2018, 645, 1230–1237. [Google Scholar] [CrossRef]
- Gelband, H.; Miller-Petrie, M.; Pant, S.; Gandra, S.; Levinson, J.; Barter, D.; White, A.; Laxminarayan, R. The state of the world’s antibiotics. Wound Heal. S. Afr. 2015, 8, 30–34. [Google Scholar]
- Zhu, Y.; Johnson, T.A.; Su, J.Q.; Qiao, M.; Guo, G.X.; Stedtfeld, R.D.; Hashsham, S.A.; Tiedje, J.M. Diverse and abundant antibiotic resistance genes in Chinese swine farms. Proc. Natl. Acad. Sci. USA 2013, 110, 3435–3440. [Google Scholar] [CrossRef] [Green Version]
- Hvistendahl, M. Public health. China takes aim at rampant antibiotic resistance. Science 2012, 336, 795. [Google Scholar] [CrossRef] [PubMed]
- Ministry of Agriculture and Rural Areas of the People’s Republic of China. Report on the Use of Veterinary Antibiotics of CHINA in 2018; Ministry of Agriculture and Rural Areas of the People’s Republic of China: Beijing, China, 2019. (In Chinese)
- Announcement of the Ministry of Agriculture and Rural Areas of the People’s Republic of China (NO. 194). Index Number: 07Bnull0271201900350; 10 July 2019. Available online: http://www.moa.gov.cn/govpublic/xmsyj/201907/t20190710_6320678.htm (accessed on 23 October 2019). (In Chinese)
- Van Boeckel, T.P.; Brower, C.; Gilbert, M.; Grenfell, B.T.; Levin, S.A.; Robinson, T.P.; Teillant, A.; Laxminarayan, R. Global trends in antimicrobial use in food animals. Proc. Natl. Acad. Sci. USA 2015, 112, 5649–5654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Boeckel, T.P.; Pires, J.; Silvester, R.; Zhao, C.; Song, J.; Criscuolo, N.G.; Gilbert, M.; Bonhoeffer, S.; Laxminarayan, R. Global trends in antimicrobial resistance in animals in low- and middle-income countries. Science 2019, 365, eaaw1944. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du, L.; Liu, W. Occurrence, fate, and ecotoxicity of antibiotics in agro-ecosystems: A review. Agron. Sustain. Dev. 2011, 32, 309–327. [Google Scholar] [CrossRef] [Green Version]
- Xu, M.; Qian, M.; Zhang, H.; Ma, J.; Wang, J.; Wu, H. Simultaneous determination of florfenicol with its metabolite based on modified quick, easy, cheap, effective, rugged, and safe sample pretreatment and evaluation of their degradation behavior in agricultural soils. J. Sep. Sci. 2015, 38, 211–217. [Google Scholar] [CrossRef]
- Qiao, M.; Chen, W.; Su, J.; Zhang, B.; Zhang, C. Fate of tetracyclines in swine manure of three selected swine farms in China. J. Environ. Sci. 2012, 24, 1047–1052. [Google Scholar] [CrossRef]
- Guo, H.; Xue, S.; Nasir, M.; Lv, J.; Gu, J. Role of Bentonite on the Mobility of Antibiotic Resistance Genes, and Microbial Community in Oxytetracycline and Cadmium Contaminated Soil. Front. Microbiol. 2018, 9, 1–11. [Google Scholar] [CrossRef]
- Guo, T.; Lou, C.; Zhai, W.; Tang, X.; Hashmi, M.Z.; Murtaza, R.; Li, Y.; Liu, X.; Xu, J. Increased occurrence of heavy metals, antibiotics and resistance genes in surface soil after long-term application of manure. Sci. Total Environ. 2018, 635, 995–1003. [Google Scholar] [CrossRef]
- Jechalke, S.; Heuer, H.; Siemens, J.; Amelung, W.; Smalla, K. Fate and effects of veterinary antibiotics in soil. Trends Microbiol. 2014, 22, 536–545. [Google Scholar] [CrossRef]
- Larsson, D.G. Antibiotics in the environment. Ups. J. Med. Sci. 2014, 119, 108–112. [Google Scholar] [CrossRef] [PubMed]
- Heuer, H.; Focks, A.; Lamshöft, M.; Smalla, K.; Matthies, M.; Spiteller, M. Fate of sulfadiazine administered to pigs and its quantitative effect on the dynamics of bacterial resistance genes in manure and manured soil. Soil. Biol. Biochem. 2008, 40, 1892–1900. [Google Scholar] [CrossRef]
- Gullberg, E.; Cao, S.; Berg, O.G.; Ilback, C.; Sandegren, L.; Hughes, D.; Andersson, D.I. Selection of resistant bacteria at very low antibiotic concentrations. PLoS. Pathog. 2011, 7, e1002158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martinez, J.L. Environmental pollution by antibiotics and by antibiotic resistance determinants. Environ. Pollut. 2009, 157, 2893–2902. [Google Scholar] [CrossRef] [PubMed]
- Jin, M.; Lu, J.; Chen, Z.; Nguyen, S.H.; Mao, L.; Li, J.; Yuan, Z.; Guo, J. Antidepressant fluoxetine induces multiple antibiotics resistance in Escherichia coli via ROS-mediated mutagenesis. Environ. Int. 2018, 120, 421–430. [Google Scholar] [CrossRef]
- Liao, J.; Huang, H.; Chen, Y. CO2 promotes the conjugative transfer of multiresistance genes by facilitating cellular contact and plasmid transfer. Environ. Int. 2019, 129, 333–342. [Google Scholar] [CrossRef]
- Zhu, L.; Zhao, Y.; Yang, K.; Chen, J.; Zhou, H.; Chen, X.; Liu, Q.; Wei, Z. Host bacterial community of MGEs determines the risk of horizontal gene transfer during composting of different animal manures. Environ. Pollut. 2019, 250, 166–174. [Google Scholar] [CrossRef]
- Asante, J.; Osei Sekyere, J. Understanding antimicrobial discovery and resistance from a metagenomic and metatranscriptomic perspective: Advances and applications. Environ. Microbiol. Rep. 2019, 11, 62–86. [Google Scholar] [CrossRef]
- Cadena, M.; Durso, L.M.; Miller, D.N.; Waldrip, H.M.; Castleberry, B.L.; Drijber, R.A.; Wortmann, C. Tetracycline and sulfonamide antibiotic resistance genes in soils from Nebraska organic farming operations. Front. Microbiol. 2018, 9, 1283. [Google Scholar] [CrossRef]
- Piddock, L.J.V. Clinically relevant chromosomally encoded multidrug resistance efflux pumps in bacteria. Clin. Microbiol. Rev. 2006, 19, 382–402. [Google Scholar] [CrossRef] [Green Version]
- Pan, M.; Chu, L.M. Occurrence of antibiotics and antibiotic resistance genes in soils from wastewater irrigation areas in the Pearl River Delta region, southern China. Sci. Total Environ. 2018, 624, 145–152. [Google Scholar] [CrossRef]
- World Health Organization (WHO). Antimicrobial Resistance: Global Report on Surveillance; World Health Organization: Geneva, Switzerland, 2014; pp. 1–256. [Google Scholar]
- Van den Bergh, K.; Huizinga, P.; Bonten, M.J.; Bos, M.; Bruyne, K.D.; Friedrich, A.W.; Rossen, J.W.; Savelkoul, P.H.; Kluytmans, J.A. Presence of mcr-1-positive Enterobacteriaceae in retail chicken meat but not in humans in the Netherlands since 2009. EuroSurveillance 2016, 21, 30149. [Google Scholar] [CrossRef] [PubMed]
- Rhouma, M.; Thériault, W.; Rabhi, N.; Duchaine, C.; Quessy, S.; Fravalo, P. First identification of mcr-1/mcr-2 genes in the fecal microbiota of Canadian commercial pigs during the growing and finishing period. Vet. Med. Res. Rep. 2019, 10, 65–67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, J.; Tian, Z.; Yu, J.; Yang, M.; Zhang, Y. Distribution and Abundance of Antibiotic Resistance Genes in Sand Settling Reservoirs and Drinking Water Treatment Plants across the Yellow River, China. Water 2018, 10, 246. [Google Scholar] [CrossRef] [Green Version]
- Von Wintersdorff, C.J.H.; Penders, J.; Stobberingh, E.E.; Lashof, A.M.L.O.; Hoebe, C.J.P.A.; Savelkoul, P.H.M.; Wolffs, P.F.G. High rates of antimicrobial drug resistance gene acquisition after international travel, the Netherlands. Emerg. Infect. Dis. 2014, 20, 649–657. [Google Scholar] [CrossRef] [PubMed]
- Schmithausen, R.M.; Schulze-Geisthoevel, S.V.; Heinemann, C.; Bierbaum, G.; Exner, M.; Petersen, B.; Steinhoff-Wagner, J. Reservoirs and Transmission Pathways of Resistant indicator bacteria in the biotope pig stable and along the food chain: A review from a one health perspective. Sustainability 2018, 10, 3967. [Google Scholar] [CrossRef] [Green Version]
- Food and Agricultural Organization of the United Nations (FAO). Sub-Committee on Aquaculture: Antimicrobial Resistance (AMR) in Aquaculture; FAO: Rome, Italy, 2017. [Google Scholar]
- Wang, N.; Yang, X.; Jiao, S.; Zhang, J.; Ye, B.; Gao, S. Sulfonamide-resistant bacteria and their resistance genes in soils fertilized with manures from Jiangsu Province, Southeastern China. PLoS ONE 2014, 9, e112626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singer, A.C.; Shaw, H.; Rhodes, V.; Hart, A. Review of Antimicrobial Resistance in the Environment and Its Relevance to Environmental Regulators. Front. Microbiol. 2016, 7, 1728. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, X.M.; Hu, H.W.; Chen, Q.L.; Yang, L.Y.; Li, H.L.; Zhu, Y.G.; Li, X.Z.; Ma, Y.B. Antibiotic resistance genes and associated bacterial communities in agricultural soils amended with different sources of animal manures. Soil. Biol. Biochem. 2018, 126, 91–102. [Google Scholar] [CrossRef]
- D’Costa, V.M.; King, C.E.; Kalan, L.; Morar, M.; Sung, W.W.; Schwarz, C.; Froese, D.; Zazula, G.; Calmels, F.; Debruyne, R.; et al. Antibiotic resistance is ancient. Nature 2011, 477, 457–461. [Google Scholar] [CrossRef]
- Durso, L.M.; Miller, D.N.; Wienhold, B.J. Distribution and quantification of antibiotic resistant genes and bacteria across agricultural and non-agricultural metagenomes. PLoS ONE 2012, 7, e48325. [Google Scholar] [CrossRef] [PubMed]
- Viana, A.T.; Caetano, T.; Covas, C.; Santos, T.; Mendo, S. Environmental superbugs: The case study of Pedobacter spp. Environ. Pollut. 2018, 241, 1048–1055. [Google Scholar] [CrossRef] [PubMed]
- Walsh, F. Investigating antibiotic resistance in non-clinical environments. Front. Microbiol. 2013, 4, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knapp, C.W.; Dolfing, J.; Ehlert, P.A.; Graham, W.D. Evidence of increasing antibiotic resistance gene abundances in archived soils since 1940. Environ. Sci. Technol. 2010, 44, 580–587. [Google Scholar] [CrossRef] [PubMed]
- Peak, N.; Knapp, C.W.; Yang, R.K.; Hanfelt, M.M.; Smith, M.S.; Aga, D.S.; Graham, D.W. Abundance of six tetracycline resistance genes in wastewater lagoons at cattle feedlots with different antibiotic use strategies. Environ. Microbiol. 2007, 9, 143–151. [Google Scholar] [CrossRef]
- Chang, C.W.; Chung, H.; Huang, C.F.; Su, H.J. Exposure of workers to airborne microorganisms in open-air swine houses. Appl. Environ. Microbiol. 2001, 67, 155–161. [Google Scholar] [CrossRef] [Green Version]
- Biswas, S.N.M.; Pandey, P.; Appuhamy, J.A.D.R.N.; Leytem, A.B.; Kebreab, E.; Dungan, R.S. Effect of Dairy Manure Storage Conditions on the Survival of E. coli O157:H7 and Listeria. J. Environ. Qual. 2018, 47, 185–189. [Google Scholar] [CrossRef] [Green Version]
- Huerta, B.; Marti, E.; Gros, M.; Lopez, P.; Pompeo, M.; Armengol, J.; Barceló, D.; Balcázar, J.L.; Rodríguez-Mozaz, S.; Marcé, R. Exploring the links between antibiotic occurrence, antibiotic resistance, and bacterial communities in water supply reservoirs. Sci. Total Environ. 2013, 456, 161–170. [Google Scholar] [CrossRef]
- Mendes, L.W.; Tsai, S.M.; Navarrete, A.A.; de Hollander, M.; van Veen, J.A.; Kuramae, E.E. Soil-borne microbiome: Linking diversity to function. Microb. Ecol. 2015, 70, 255–265. [Google Scholar] [CrossRef]
- Cui, E.; Wu, Y.; Zuo, Y.; Chen, H. Effect of different biochars on antibiotic resistance genes and bacterial community during chicken manure composting. Bioresour. Technol. 2016, 203, 11–17. [Google Scholar] [CrossRef]
- Li, N.; Zhu, C.; Liu, C.; Zhang, X.; Ding, J.; Zandi, P.; Li, H. The persistence of antimicrobial resistance and related environmental factors in abandoned and working swine feedlots. Environ. Pollut. 2019, 255, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Li, B.; Zhang, Z.; Zhu, C.; Tian, Y.; Ye, J. Evolution of microbial communities during electrokinetic treatment of antibiotic-polluted soil. Ecotoxicol. Environ. Saf. 2018, 148, 842–850. [Google Scholar] [CrossRef] [PubMed]
- Hong, P.Y.; Li, X.; Yang, X.; Shinkai, T.; Zhang, Y.; Wang, X.; Mackie, R.I. Monitoring airborne biotic contaminants in the indoor environment of pig and poultry confinement buildings. Environ. Microbiol. 2012, 14, 1420–1431. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.; Wang, Y.; Liu, S.; Huang, J.; Zhai, Z.; He, C.; Ding, J.; Wang, J.; Wang, H.; Fan, W.; et al. The dynamic distribution of porcine microbiota across different ages and gastrointestinal tract segments. PLoS ONE 2015, 10, e0117441. [Google Scholar] [CrossRef] [Green Version]
- Nelson, K.E. An Update on the Status of Current Research on the Mammalian Microbiome. ILAR J. 2015, 56, 163–168. [Google Scholar] [CrossRef] [Green Version]
- Wichmann, F.; Udikovic-Kolic, N.; Andrew, S.; Handelsman, J. Diverse antibiotic resistance genes in dairy cow manure. MBio. 2014, 5, e01017. [Google Scholar] [CrossRef] [Green Version]
- Forsberg, K.J.; Patel, S.; Gibson, M.K.; Lauber, C.L.; Knight, R.; Fierer, N.; Dantas, G. Bacterial phylogeny structures soil resistomes across habitats. Nature 2014, 509, 612–616. [Google Scholar] [CrossRef] [Green Version]
- Fan, X.; Li, H.; Chen, Q.; Zhang, Y.; Ye, J.; Zhu, Y.; Su, J. Fate of antibiotic resistant Pseudomonas putida and broad host range plasmid in natural soil microcosms. Front. Microbiol. 2009, 10, 194. [Google Scholar] [CrossRef] [Green Version]
- He, L.Y.; Ying, G.G.; Liu, Y.S.; Su, H.C.; Chen, J.; Liu, S.S.; Zhao, J.L. Discharge of swine wastes risks water quality and food safety: Antibiotics and antibiotic resistance genes from swine sources to the receiving environments. Environ. Int. 2016, 92, 210–219. [Google Scholar] [CrossRef]
- Dungan, R.S.; McKinney, C.W.; Leytem, A.B. Tracking antibiotic resistance genes in soil irrigated with dairy wastewater. Sci. Total. Environ. 2018, 635, 1477–1483. [Google Scholar] [CrossRef]
- Chen, Z.; Yang, L.; Stedtfeld, R.D.; Peng, A.; Gu, C.; Boyd, S.A.; Li, H. Antibiotic resistance genes and bacterial communities in cornfield and pasture soils receiving swine and dairy manures. Environ. Pollut. 2019, 248, 947–957. [Google Scholar] [CrossRef]
- Agga, G.E.; Cook, K.L.; Netthisinghe, A.M.P.; Gilfillen, R.A.; Woosley, P.B.; Sistani, K.R. Persistence of antibiotic resistance genes in beef cattle backgrounding environment over two years after cessation of operation. PLoS ONE 2019, 14, e0212510. [Google Scholar] [CrossRef] [Green Version]
- Peng, S.; Wang, Y.; Zhou, B.; Lin, X. Long-term application of fresh and composted manure increase tetracycline resistance in the arable soil of eastern China. Sci. Total Environ. 2015, 506, 279–286. [Google Scholar] [CrossRef]
- Tang, X.; Lou, C.; Wang, S.; Lu, Y.; Liu, M.; Hashmi, M.Z.; Liang, X.; Li, Z.; Liao, Y.; Qin, W.; et al. Effects of long-term manure applications on the occurrence of antibiotics and antibiotic resistance genes (ARGs) in paddy soils: Evidence from four field experiments in south of China. Soil. Biol. Biochem. 2015, 90, 179–187. [Google Scholar] [CrossRef]
- Huang, X.; Liu, C.; Li, K.; Liu, F.; Liao, D.; Liu, L.; Zhu, G.; Liao, J. Liao. Occurrence and distribution of veterinary antibiotics and tetracycline resistance genes in farmland soils around swine feedlots in Fujian Province, China. Environ. Sci. Pollut. Res. Int. 2013, 20, 9066–9074. [Google Scholar] [CrossRef]
- Li, B.; Yang, Y.; Ma, L.; Ju, F.; Guo, F.; Tiedje, J.M.; Zhang, T. Metagenomic and network analysis reveal wide distribution and co-occurrence of environmental antibiotic resistance genes. ISME J. 2015, 9, 2490–2502. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Liu, C.; Chen, Y.; Huang, H.; Ren, T. Antibiotic residues in liquid manure from swine feedlot and their effects on nearby groundwater in regions of North China. Environ. Sci. Pollut. Res. 2018, 25, 11565–11575. [Google Scholar] [CrossRef]
- Liu, C.L.; Zheng, G.D.; Wang, L.; Chen, T.B.; Shao, Z.Z.; Chen, L. Sources and pollution characteristics of antibiotic resistance genes and conditional pathogenic bacteria in concentrated swine feeding operation. Chin. J. Appl. Ecol. 2018, 8, 1–10. (In Chinese) [Google Scholar]
- Lim, J.A.; Hong, J.; Kim, J.; Heu, S.; Roh, E. OmpF of Pectobacterium carotovorum subsp. carotovorum Pcc3 is required for carocin D sensitivity. FEMS. Microbiol. Lett. 2016, 363. [Google Scholar] [CrossRef] [Green Version]
- McEachran, A.D.; Blackwell, B.R.; Hanson, J.D.; Wooten, K.J.; Mayer, G.D.; Cox, S.B.; Smith, P.N. Antibiotics, bacteria, and antibiotic resistance genes: Aerial transport from cattle feed yards via particulate matter. Environ. Health Perspect. 2015, 123, 337–343. [Google Scholar] [CrossRef] [Green Version]
- Kempf, I.; Kerouanton, A.; Bougeard, S.; Nagard, B.; Rose, V.; Mourand, G.; Osterberg, J.; Denis, M.; Bengtsson, B.O. Campylobacter coli in Organic and Conventional Pig Production in France and Sweden: Prevalence and Antimicrobial Resistance. Front. Microbiol. 2017, 8, 955. [Google Scholar] [CrossRef]
- Su, J.; Cui, L.; Chen, Q.; An, X.; Zhu, Y. Application of genomic technologies to measure and monitor antibiotic resistance in animals. Ann. N. Y. Acad. Sci. 2017, 1388, 121–135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Enne, V.I.; Cassar, C.; Sprigings, K.; Woodward, M.J.; Bennett, P.M. A high prevalence of antimicrobial resistant Escherichia coli isolated from pigs and a low prevalence of antimicrobial resistant E. coli from cattle and sheep in Great Britain at slaughter. FEMS. Microbiol. Lett. 2008, 278, 193–199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bech, T.B.; Johnsen, K.; Dalsgaard, A.; Laegdsmand, M.; Jacobsen, O.H.; Jacobsen, C.S. Transport and distribution of Salmonella enterica serovar Typhimurium in loamy and sandy soil monoliths with applied liquid manure. Appl. Environ. Microbiol. 2010, 76, 710–714. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keer, J.T.; Birch, L. Molecular methods for the assessment of bacterial viability. J. Microbiol. Methods. 2003, 53, 175–183. [Google Scholar] [CrossRef]
- Kibbee, R.J.; Ormeci, B. Development of a sensitive and false-positive free PMA-qPCR viability assay to quantify VBNC Escherichia coli and evaluate disinfection performance in wastewater effluent. J. Microbiol. Methods 2017, 132, 139–147. [Google Scholar] [CrossRef]
- Vondrakova, L.T.H.; Scholtz, V.; Pazlarova, J.; Demnerova, K. Impact of various killing methods on EMA/PMA-qPCR efficacy. Food. Control. 2018, 85, 23–28. [Google Scholar] [CrossRef]
- Negreanu, Y.P.Z.; Jurkevitch, E.; Cytryn, E. Impact of treated wastewater irrigation on antibiotic resistance in agricultural soils. Environ. Sci. Technol. 2012, 46, 4800–4808. [Google Scholar] [CrossRef]
- Prosser, J.I. Dispersing misconceptions and identifying opportunities for the use of ‘omics’ in soil microbial ecology. Nat. Rev. Microbiol. 2015, 13, 439–446. [Google Scholar] [CrossRef]
- Walsh, F.; Ingenfeld, A.; Zampicolli, M.; Hilber-Bodmer, M.; Frey, J.E.; Duffy, B. Real-time PCR methods for quantitative monitoring of streptomycin and tetracycline resistance genes in agricultural ecosystems. J. Microbiol. Methods 2011, 86, 150–155. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Tuo, X.; Gu, J.; Wang, X.; Sun, Y.; Duan, M.; Sun, W.; Yin, Y.; Guo, A.; Zhang, L. Prevalence of quinolone resistance genes, copper resistance genes, and the bacterial communities in a soil-ryegrass system co-polluted with copper and ciprofloxacin. Chemosphere 2018, 197, 643–650. [Google Scholar] [CrossRef] [PubMed]
- Cheng, W.; Chen, H.; Su, C.; Yan, S. Abundance and persistence of antibiotic resistance genes in livestock farms: A comprehensive investigation in eastern China. Environ. Int. 2013, 61, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Pu, C.; Liu, H.; Ding, G.; Sun, Y.; Yu, X.; Chen, J.; Ren, J.; Gong, X. Impact of direct application of biogas slurry and residue in fields: In situ analysis of antibiotic resistance genes from pig manure to fields. J. Hazard. Mater. 2018, 344, 441–449. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.; Rieke, E.L..; Moorman, T.B.; Soupir, M.L.; Allen, H.K.; Smith, S.D.; Howe, A. Practical implications of erythromycin resistance gene diversity on surveillance and monitoring of resistance. FEMS. Microbiol. Ecol. 2018, 94, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Thanner, S.; Drissner, D.; Walsh, F. Antimicrobial Resistance in Agriculture. MBio 2016, 7, e02227-15. [Google Scholar] [CrossRef] [Green Version]
- Zhu, B.; Chen, Q.; Chen, S.; Zhu, Y. Does organically produced lettuce harbor higher abundance of antibiotic resistance genes than conventionally produced? Environ. Int. 2017, 98, 152–159. [Google Scholar] [CrossRef]
- Zhu, Y.; Zhao, Y.; Li, B.; Huang, C.; Zhang, S.; Yu, S.; Chen, Y.; Zhang, T.; Gillings, M.R.; Su, J. Continental-scale pollution of estuaries with antibiotic resistance genes. Nat. Microbiol. 2017, 2, 16270. [Google Scholar] [CrossRef]
- Zhang, J.; Wang, Z.; Wang, Y.; Zhong, H.; Sui, Q.; Zhang, C.; Wei, Y. Effects of graphene oxide on the performance, microbial community dynamics and antibiotic resistance genes reduction during anaerobic digestion of swine manure. Bioresour. Technol. 2017, 245, 850–859. [Google Scholar] [CrossRef]
- Su, Y.; Wang, J.; Xia, H.; Xie, B. Comparative network analysis revealing the mechanisms of antibiotic resistance genes removal by leachate recirculation under different hydraulic loadings. Sci. Total. Environ. 2019, 649, 318–326. [Google Scholar] [CrossRef]
- Zhang, S.; Han, B.; Gu, J.; Wang, C.; Wang, P.; Ma, Y.; Gao, J.; He, Z. Fate of antibiotic resistant cultivable heterotrophic bacteria and antibiotic resistance genes in wastewater treatment processes. Chemosphere. 2015, 135, 138–145. [Google Scholar] [CrossRef] [PubMed]
- Sharpton, T.J. An introduction to the analysis of shotgun metagenomic data. Front. Plant. Sci. 2014, 5, 209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aguiar-Pulido, V.; Huang, W.; Suarez-Ulloa, V.; Cickovski, T.; Mathee, K.; Narasimhan, G. Metagenomics, metatranscriptomics, and metabolomics approaches for microbiome analysis. Evol. Bioinform. Online 2016, 12, 5–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tietgen, M.; Semmler, T.; Riedel-Christ, S.; Kempf, V.A.J.; Molinaro, A.; Ewers, C.; Gottig, S. Impact of the colistin resistance gene mcr-1 on bacterial fitness. Int. J. Antimicrob. Agents 2018, 51, 554–561. [Google Scholar] [CrossRef] [PubMed]
- Moura, A.; Soares, M.; Pereira, C.; Leitao, N.; Henriques, I.; Correia, A. INTEGRALL: A database and search engine for integrons, integrases and gene cassettes. Bioinformatics 2009, 25, 1096–1098. [Google Scholar] [CrossRef] [Green Version]
- Siguier, P.; Perochon, J.; Lestrade, L.; Mahillon, J.; Chandler, M. ISfinder: The reference centre for bacterial insertion sequences. Nucleic. Acids. Res. 2006, 34, D32–D36. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Yang, G.; Hou, S.; Zhang, T.; Li, Z.; Liang, F. Distribution of ARGs and MGEs among glacial soil, permafrost, and sediment using metagenomic analysis. Environ. Pollut. 2018, 234, 339–346. [Google Scholar] [CrossRef]
- Gupta, S.K.; Padmanabhan, B.R.; Diene, S.M.; Lopez-Rojas, R.; Kempf, M.; Landraud, L.; Rolain, J.M. ARG-ANNOT, a new bioinformatic tool to discover antibiotic resistance genes in bacterial genomes. Antimicrob. Agents Chemother. 2014, 58, 212–220. [Google Scholar] [CrossRef] [Green Version]
- Rowe, W.P.M.; Baker-Austin, C.; Verner-Jeffreys, D.W.; Ryan, J.J.; Micallef, C.; Maskell, D.J.; Pearce, G.P. Overexpression of antibiotic resistance genes in hospital effluents over time. J. Antimicrob. Chemother. 2017, 72, 1617–1623. [Google Scholar] [CrossRef]
- Arango-Argoty, G.; Garner, E.; Pruden, A.; Heath, L.S.; Vikesland, P.; Zhang, L. DeepARG: A deep learning approach for predicting antibiotic resistance genes from metagenomic data. Microbiome 2018, 6, 23. [Google Scholar] [CrossRef] [Green Version]
- Hadjadj, L.; Baron, S.A.; Diene, S.M.; Rolain, J.M. How to discover new antibiotic resistance genes? Expert. Rev. Mol. Diagn. 2019, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; He, Z.; Yang, Y.; Deng, Y.; Tringe, S.G.; Alvarez-Cohen, L. High-throughput metagenomic technologies for complex microbial community analysis: Open and closed formats. Mbio 2015, 6, e02288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rhoads, A.; Au, K.F. PacBio Sequencing and its applications. Genom. Proteom. Bioinf. 2015, 13, 278–289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wenger, A.M.; Peluso, P.; Rowell, W.J.; Chang, P.C.; Hall, R.J.; Concepcion, G.T.; Ebler, J.; Fungtammasan, A.; Kolesnikov, A.; Olson, N.D.; et al. Accurate circular consensus long-read sequencing improves variant detection and assembly of a human genome. Nat. Biotechnol. 2019. [CrossRef] [PubMed]
- Nostrand, J.D.; He, Z.; Zhou, J. Use of functional gene arrays for elucidating in situ biodegradation. Front. Microbiol. 2012, 3, 339. [Google Scholar] [CrossRef] [Green Version]
- Low, A.; Ng, C.; He, J. Identification of antibiotic resistant bacteria community and a GeoChip based study of resistome in urban watersheds. Water. Res. 2016, 106, 330–338. [Google Scholar] [CrossRef]
- Zhang, B.; Xia, Y.; Wen, X.; Wang, X.; Yang, Y.; Zhou, J.; Zhang, Y. The composition and spatial patterns of bacterial virulence factors and antibiotic resistance genes in 19 wastewater treatment plants. PLoS. ONE 2016, 11, e0167422. [Google Scholar] [CrossRef] [Green Version]
- Wei, S.T.S.; Higgins, C.M.; Adriaenssens, E.M.; Cowan, D.A.; Pointing, S.B. Genetic signatures indicate widespread antibiotic resistance and phage infection in microbial communities of the McMurdo Dry Valleys, East Antarctica. Polar. Biol. 2015, 38, 919–925. [Google Scholar] [CrossRef]
- Jiang, X.; Hall, A.B.; Arthur, T.D.; Plichta, D.R.; Covington, C.T.; Poyet, M.; Crothers, J.; Moses, P.L.; Tolonen, A.C.; Vlamakis, H.; et al. Invertible promoters mediate bacterial phase variation, antibiotic resistance, and host adaptation in the gut. Science 2019, 363, 181–187. [Google Scholar] [CrossRef]


{kind=link}
| Sources | Environmental Matrices | Methods | ARG or Antimicrobial-Resistant Bacteria (ARB) Pollution Results | Reference |
|---|---|---|---|---|
| Dairy farm | Agricultural soil | Quantitative PCR (qPCR) | Wastewater significantly increased the relative ARG abundance in soil | [67] |
| Pig, chicken, and cow manure | Soil | High-throughput qPCR and 16S rDNA taxonomic composition | Manure fertilizer significantly increased the ARG abundance | [68] |
| A semi-intensive beef cattle farm | Soil in feeding and grazing area | qPCR | ARG abundance was negatively correlated with distance from feeding area and abandonment time. Two years after abandonment of cattle farm, ARG pollution still existed | [69] |
| Pig manure | Soil | qPCR | 2×10−5 to 0.0374 ARG copies/16S rDNA | [70] |
| Manure | Soil | PCR | Layering over 15 cm did not distinguish the vertical ARG distribution | [34] |
| Manure | Soil | qPCR | 10−7 to 10−3 ARG copies/16S rDNA | [71] |
| Pig manure | Soil | qPCR | Antimicrobial residues and ARGs were found at 60–80 cm depths | [72] |
| Livestock and others | Multiple environmental matrices | Metagenomic sequencing | Relative ARG abundance: Animal manure > WWTP > river water, soil, and fish pond sediments | [73] |
| Pig farm | Multiple environmental matrices | qPCR | Antimicrobials and ARGs could penetrate into groundwater, resulting in groundwater pollution | [74] |
| Pig farms | Aerosols and pig manure | qPCR | The ermB, ermF, and tetW in pig manure >109 copies·g−1; ARGs in aerosols were 104 to 107 copies·m− | [75] |
| Poultry farms | Aerosol | PCR | 360 strains of E. coli were isolated; 47 strains were non-resistant, many were multiply-resistant | [76] |
| Beef cattle feed yards | Aerosol | qPCR | ARGs were more abundant downwind compared to upwind PM of feed yards | [77] |
| Pig, layer, and turkey farms | Aerosol | 16S rDNA taxonomic composition and qPCR | The abundance of tetracycline ARGs were 102 to 106 copies/ng DNA | [60] |
| Method | Advantages | Disadvantages | Application Scope |
|---|---|---|---|
| Culture method | Able to determine the MIC of culturable bacteria and phenotypic changes under antimicrobial selective pressure. | Tedious process; unable for analyzing comprehensive ARG transmission risk on a community level | Ex situ phenotypic characterization; selecting and determining MIC of ARB; constructing gene library |
| PCR/qPCR | Able to determine ARG presence or abundance, offering ARG basic transmission risk | Low-throughput; PCR bias exists; Cannot distinguish between live and dead cell or ARG hosts in a complex community | Determining the presence or abundance of certain interested ARGs with knowing host or obtaining the gene pool |
| High-throughput qPCR | High-throughput format plus the advantages of qPCR | PCR bias exists; rough abundance data due to the same protocol set for multiple primers | Determines multiple ARGs in environmental microbiota |
| 16S rDNA amplicon sequencing | Allows analysis for bacterial taxa in ecosystems and co-occurrence analysis for bacteria and ARGs | Unable to link ARGs to hosts; PCR bias before sequencing can sometimes influence results | Analyzes bacterial community structure, and potential ARG hosts via co-occurrence analysis |
| Metagenomic analysis | Allows annotation of all functional genes, making it possible to predict ARG hosts | Poor repeatability and high cost; PCR bias exists; advanced computer system capable of processing huge data sets needed; not sensitive to test low abundance taxa. | Open format analysis allows query of broad characteristics and can identify novel genes; determines the community wide ARG pool |
| GeoChip | PCR-independent; excellent repeatability and accuracy; high sensitivity enabling detection of low abundance species and genes | Unable to explore novel species or functional genes; potentially underestimating the diversity of microbial taxa and functional genes | Closed format analysis (towards known species and target genes) for phylogenetic and functional genes |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, N.; Liu, C.; Zhang, Z.; Li, H.; Song, T.; Liang, T.; Li, B.; Li, L.; Feng, S.; Su, Q.; et al. Research and Technological Advances Regarding the Study of the Spread of Antimicrobial Resistance Genes and Antimicrobial-Resistant Bacteria Related to Animal Husbandry. Int. J. Environ. Res. Public Health 2019, 16, 4896. https://doi.org/10.3390/ijerph16244896
Li N, Liu C, Zhang Z, Li H, Song T, Liang T, Li B, Li L, Feng S, Su Q, et al. Research and Technological Advances Regarding the Study of the Spread of Antimicrobial Resistance Genes and Antimicrobial-Resistant Bacteria Related to Animal Husbandry. International Journal of Environmental Research and Public Health. 2019; 16(24):4896. https://doi.org/10.3390/ijerph16244896
Chicago/Turabian StyleLi, Na, Chong Liu, Zhiguo Zhang, Hongna Li, Tingting Song, Ting Liang, Binxu Li, Luyao Li, Shuo Feng, Qianqian Su, and et al. 2019. "Research and Technological Advances Regarding the Study of the Spread of Antimicrobial Resistance Genes and Antimicrobial-Resistant Bacteria Related to Animal Husbandry" International Journal of Environmental Research and Public Health 16, no. 24: 4896. https://doi.org/10.3390/ijerph16244896
APA StyleLi, N., Liu, C., Zhang, Z., Li, H., Song, T., Liang, T., Li, B., Li, L., Feng, S., Su, Q., Ye, J., & Zhu, C. (2019). Research and Technological Advances Regarding the Study of the Spread of Antimicrobial Resistance Genes and Antimicrobial-Resistant Bacteria Related to Animal Husbandry. International Journal of Environmental Research and Public Health, 16(24), 4896. https://doi.org/10.3390/ijerph16244896