Metabolite Changes in Maternal and Fetal Plasma Following Spontaneous Labour at Term in Humans Using Untargeted Metabolomics Analysis: A Pilot Study

Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Collection of Samples
2.3. Metabolomic Analysis
2.4. Statistical Analysis
3. Results
3.1. Demographics
3.2. Metabolomics
3.3. Changes in Cord Plasma Metabolome
3.4. Changes in Maternal Plasma Metabolome
3.5. Xenobiotics
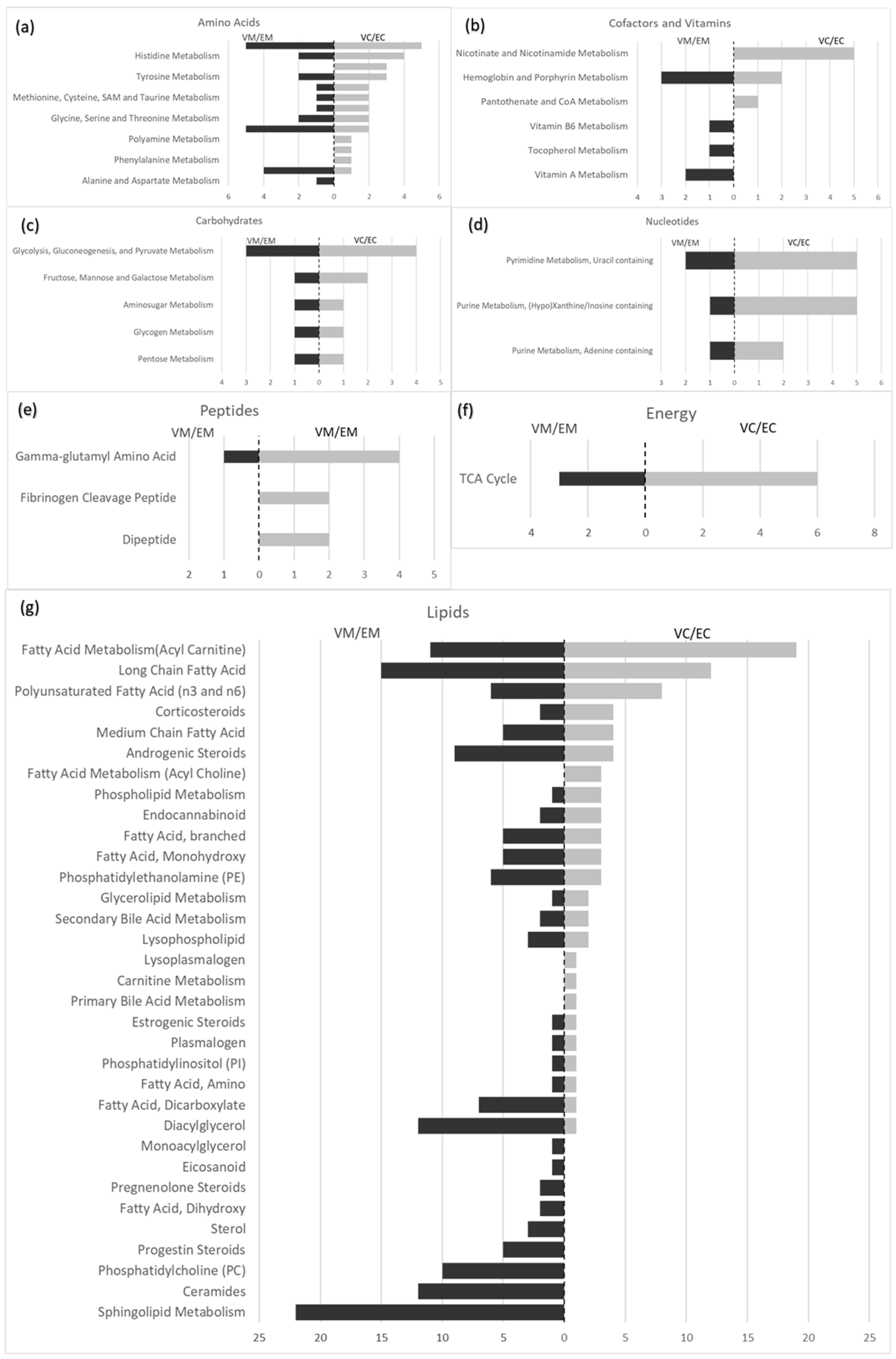
3.6. Lipid Super-Pathway
3.7. Nucleotides
3.8. Amino Acids
3.9. Peptides
3.10. Carbohydrate Metabolism
3.11. Energy Super-Pathway
3.12. Co-Factors and Vitamins
4. Discussion
4.1. Endocannabinoids, Ceramides and Sphingolipids
4.2. Prostaglandins
4.3. Progesterone and Pregnenolone
4.4. Estrogen
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Lopez Bernal, A. Overview. Preterm labour: Mechanisms and management. BMC Pregnancy Childbirth 2007, 7, S2. [Google Scholar] [CrossRef] [PubMed]
- Chang, H.H.; Larson, J.; Blencowe, H.; Spong, C.Y.; Howson, C.P.; Cairns-Smith, S.; Lackritz, E.M.; Lee, S.K.; Mason, E.; Serazin, A.C.; et al. Preventing preterm births: Analysis of trends and potential reductions with interventions in 39 countries with very high human development index. Lancet 2013, 381, 223–234. [Google Scholar] [CrossRef]
- Lozano, R.; Naghavi, M.; Foreman, K.; Lim, S.; Shibuya, K.; Aboyans, V.; Abraham, J.; Adair, T.; Aggarwal, R.; Ahn, S.Y.; et al. Global and regional mortality from 235 causes of death for 20 age groups in 1990 and 2010: A systematic analysis for the Global Burden of Disease Study 2010. Lancet 2012, 380, 2095–2128. [Google Scholar] [CrossRef]
- Middleton, P.; Shepherd, E.; Crowther, C.A. Induction of labour for improving birth outcomes for women at or beyond term. Cochrane Database Syst. Rev. 2018, 5, Cd004945. [Google Scholar] [CrossRef] [PubMed]
- National Collaborating Centre for Women’s and Children’s Health. National Collaborating Centre for Women’s and Children’s Health. National Institute for Health and Care Excellence: Clinical Guidelines. In Preterm Labour and Birth; National Institute for Health and Care Excellence (UK) Copyright (c) 2015; National Collaborating Centre for Women’s and Children’s Health: London, UK, 2015. [Google Scholar]
- National Collaborating Centre for Women’s and Children’s Health. National Collaborating Centre for Women’s and Children’s Health. National Institute for Health and Care Excellence: Clinical Guidelines. In Intrapartum Care: Care of Healthy Women and Their Babies During Childbirth; National Institute for Health and Care Excellence (UK) Copyright (c) 2014; National Collaborating Centre for Women’s and Children’s Health: London, UK, 2014. [Google Scholar]
- Menon, R.; Bonney, E.A.; Condon, J.; Mesiano, S.; Taylor, R.N. Novel concepts on pregnancy clocks and alarms: Redundancy and synergy in human parturition. Hum Reprod Update 2016, 22, 535–560. [Google Scholar] [CrossRef] [PubMed]
- National Health Service Digital. NHS Maternity Statistics 2016–2017; Health and Social Care Information Centre: Leeds, UK, 2017.
- Grobman, W.A.; Rice, M.M.; Reddy, U.M.; Tita, A.T.N.; Silver, R.M.; Mallett, G.; Hill, K.; Thom, E.A.; El-Sayed, Y.Y.; Perez-Delboy, A.; et al. Labor Induction versus Expectant Management in Low-Risk Nulliparous Women. N. Engl. J. Med. 2018, 379, 513–523. [Google Scholar] [CrossRef]
- Keirse, M.J. Elective induction, selective deduction, and cesarean section. Birth 2010, 37, 252–256. [Google Scholar] [CrossRef] [PubMed]
- Mealing, N.M.; Roberts, C.L.; Ford, J.B.; Simpson, J.M.; Morris, J.M. Trends in induction of labour, 1998-2007: A population-based study. Aust. N. Z. J. Obstet. Gynaecol. 2009, 49, 599–605. [Google Scholar] [CrossRef] [PubMed]
- National Institute for Clinical and Health Excellence. Inducing Labour Clinical Guideline CG70; National Institute for Clinical and Health Excellence: Manchester, UK, 2008. [Google Scholar]
- Georgiou, H.M.; Di Quinzio, M.K.; Permezel, M.; Brennecke, S.P. Predicting Preterm Labour: Current Status and Future Prospects. Dis. Markers 2015, 2015, 435014. [Google Scholar] [CrossRef]
- Kruit, H.; Heikinheimo, O.; Sorsa, T.; Juhila, J.; Paavonen, J.; Rahkonen, L. Cervical biomarkers as predictors of successful induction of labour by Foley catheter. J. Obstet. Gynaecol. 2018, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Roberts, L.D.; Souza, A.L.; Gerszten, R.E.; Clish, C.B. Targeted metabolomics. Curr. Protoc. Mol. Biol. 2012. [Google Scholar] [CrossRef] [PubMed]
- Lindon, J.C.; Holmes, E.; Nicholson, J.K. Metabonomics in pharmaceutical R&D. FEBS J. 2007, 274, 1140–1151. [Google Scholar] [CrossRef] [PubMed]
- Nicholson, J.K.; Lindon, J.C.; Holmes, E. ‘Metabonomics’: Understanding the metabolic responses of living systems to pathophysiological stimuli via multivariate statistical analysis of biological NMR spectroscopic data. Xenobiotica 1999, 29, 1181–1189. [Google Scholar] [CrossRef] [PubMed]
- Wishart, D.S. Emerging applications of metabolomics in drug discovery and precision medicine. Nat. Rev. Drug Discov. 2016, 15, 473–484. [Google Scholar] [CrossRef] [PubMed]
- Atzori, L.; Barberini, L.; Santoru, M.L.; Antonucci, R.; Fanos, V. Metabolomics explained to perinatologists and pediatricians. J. Matern. Fetal Neonatal Med. 2012, 25, 10–12. [Google Scholar] [CrossRef]
- Caboni, P.; Meloni, A.; Lussu, M.; Carta, E.; Barberini, L.; Noto, A.; Deiana, S.F.; Mereu, R.; Ragusa, A.; Paoletti, A.M.; et al. Urinary metabolomics of pregnant women at term: A combined GC/MS and NMR approach. J. Matern. Fetal Neonatal Med. 2014, 27, 4–12. [Google Scholar] [CrossRef]
- Virgiliou, C.; Gika, H.G.; Witting, M.; Bletsou, A.A.; Athanasiadis, A.; Zafrakas, M.; Thomaidis, N.S.; Raikos, N.; Makrydimas, G.; Theodoridis, G.A. Amniotic Fluid and Maternal Serum Metabolic Signatures in the Second Trimester Associated with Preterm Delivery. J. Proteome Res. 2017, 16, 898–910. [Google Scholar] [CrossRef]
- Wu, Z.; Jin, L.; Zheng, W.; Zhang, C.; Zhang, L.; Chen, Y.; Guan, J.; Fei, H. NMR-based serum metabolomics study reveals a innovative diagnostic model for missed abortion. Biochem. Biophys. Res. Commun. 2018, 496, 679–685. [Google Scholar] [CrossRef]
- Fattuoni, C.; Pietrasanta, C.; Pugni, L.; Ronchi, A.; Palmas, F.; Barberini, L.; Dessi, A.; Pintus, R.; Fanos, V.; Noto, A.; et al. Urinary metabolomic analysis to identify preterm neonates exposed to histological chorioamnionitis: A pilot study. PLoS ONE 2017, 12, e0189120. [Google Scholar] [CrossRef]
- Fotiou, M.; Fotakis, C.; Tsakoumaki, F.; Athanasiadou, E.; Kyrkou, C.; Dimitropoulou, A.; Tsiaka, T.; Chatziioannou, A.C.; Sarafidis, K.; Menexes, G.; et al. (1)H NMR-based metabolomics reveals the effect of maternal habitual dietary patterns on human amniotic fluid profile. Sci. Rep. 2018, 8, 4076. [Google Scholar] [CrossRef]
- Yuan, W.; Heesom, K.; Phillips, R.; Chen, L.; Trinder, J.; Lopez Bernal, A. Low abundance plasma proteins in labour. Reproduction 2012, 144, 505–518. [Google Scholar] [CrossRef] [Green Version]
- Montrose, D.C.; Zhou, X.K.; Kopelovich, L.; Yantiss, R.K.; Karoly, E.D.; Subbaramaiah, K.; Dannenberg, A.J. Metabolic profiling, a noninvasive approach for the detection of experimental colorectal neoplasia. Cancer Prev. Res. 2012, 5, 1358–1367. [Google Scholar] [CrossRef]
- Hiltunen, T.P.; Rimpela, J.M.; Mohney, R.P.; Stirdivant, S.M.; Kontula, K.K. Effects of four different antihypertensive drugs on plasma metabolomic profiles in patients with essential hypertension. PLoS ONE 2017, 12, e0187729. [Google Scholar] [CrossRef]
- Shin, S.Y.; Fauman, E.B.; Petersen, A.K.; Krumsiek, J.; Santos, R.; Huang, J.; Arnold, M.; Erte, I.; Forgetta, V.; Yang, T.P.; et al. An atlas of genetic influences on human blood metabolites. Nat. Genet. 2014, 46, 543–550. [Google Scholar] [CrossRef] [Green Version]
- Dehaven, C.D.; Evans, A.M.; Dai, H.; Lawton, K.A. Organization of GC/MS and LC/MS metabolomics data into chemical libraries. J. Cheminform. 2010, 2, 9. [Google Scholar] [CrossRef]
- Lehmann, R.; Zhao, X.; Weigert, C.; Simon, P.; Fehrenbach, E.; Fritsche, J.; Machann, J.; Schick, F.; Wang, J.; Hoene, M.; et al. Medium chain acylcarnitines dominate the metabolite pattern in humans under moderate intensity exercise and support lipid oxidation. PLoS ONE 2010, 5, e11519. [Google Scholar] [CrossRef]
- Peake, J.M.; Tan, S.J.; Markworth, J.F.; Broadbent, J.A.; Skinner, T.L.; Cameron-Smith, D. Metabolic and hormonal responses to isoenergetic high-intensity interval exercise and continuous moderate-intensity exercise. Am. J. Physiol. Endocrinol. Metab. 2014, 307, E539–E552. [Google Scholar] [CrossRef]
- Yin, P.; Lehmann, R.; Xu, G. Effects of pre-analytical processes on blood samples used in metabolomics studies. Anal. Bioanal. Chem. 2015, 407, 4879–4892. [Google Scholar] [CrossRef] [Green Version]
- Manolopoulos, K.N.; O’Reilly, M.W.; Bujalska, I.J.; Tomlinson, J.W.; Arlt, W. Acute Hypercortisolemia Exerts Depot-Specific Effects on Abdominal and Femoral Adipose Tissue Function. J. Clin. Endocrinol. Metab. 2017, 102, 1091–1101. [Google Scholar] [CrossRef] [Green Version]
- Lindsay, K.L.; Hellmuth, C.; Uhl, O.; Buss, C.; Wadhwa, P.D.; Koletzko, B.; Entringer, S. Longitudinal Metabolomic Profiling of Amino Acids and Lipids across Healthy Pregnancy. PLoS ONE 2015, 10, e0145794. [Google Scholar] [CrossRef]
- Hill, M.; Paskova, A.; Kanceva, R.; Velikova, M.; Kubatova, J.; Kancheva, L.; Adamcova, K.; Mikesova, M.; Zizka, Z.; Koucky, M.; et al. Steroid profiling in pregnancy: A focus on the human fetus. J. Steroid. Biochem. Mol. Biol. 2014, 139, 201–222. [Google Scholar] [CrossRef] [PubMed]
- Petraglia, F.; Imperatore, A.; Challis, J.R. Neuroendocrine mechanisms in pregnancy and parturition. Endocr. Rev. 2010, 31, 783–816. [Google Scholar] [CrossRef] [PubMed]
- Immenschuh, S.; Vijayan, V.; Janciauskiene, S.; Gueler, F. Heme as a Target for Therapeutic Interventions. Front. Pharmacol. 2017, 8, 146. [Google Scholar] [CrossRef] [PubMed]
- Rapino, C.; Battista, N.; Bari, M.; Maccarrone, M. Endocannabinoids as biomarkers of human reproduction. Hum. Reprod. Update 2014, 20, 501–516. [Google Scholar] [CrossRef] [Green Version]
- Cianchi, F.; Papucci, L.; Schiavone, N.; Lulli, M.; Magnelli, L.; Vinci, M.C.; Messerini, L.; Manera, C.; Ronconi, E.; Romagnani, P.; et al. Cannabinoid receptor activation induces apoptosis through tumor necrosis factor alpha-mediated ceramide de novo synthesis in colon cancer cells. Clin. Cancer Res. 2008, 14, 7691–7700. [Google Scholar] [CrossRef] [PubMed]
- Marczylo, T.H.; Lam, P.M.; Amoako, A.A.; Konje, J.C. Anandamide levels in human female reproductive tissues: Solid-phase extraction and measurement by ultraperformance liquid chromatography tandem mass spectrometry. Anal. Biochem. 2010, 400, 155–162. [Google Scholar] [CrossRef]
- Maccarrone, M.; Valensise, H.; Bari, M.; Lazzarin, N.; Romanini, C.; Finazzi-Agro, A. Relation between decreased anandamide hydrolase concentrations in human lymphocytes and miscarriage. Lancet 2000, 355, 1326–1329. [Google Scholar] [CrossRef]
- Nallendran, V.; Lam, P.M.; Marczylo, T.H.; Bankart, M.J.; Taylor, A.H.; Taylor, D.J.; Konje, J.C. The plasma levels of the endocannabinoid, anandamide, increase with the induction of labour. Bjog 2010, 117, 863–869. [Google Scholar] [CrossRef] [Green Version]
- Dennedy, M.C.; Friel, A.M.; Houlihan, D.D.; Broderick, V.M.; Smith, T.; Morrison, J.J. Cannabinoids and the human uterus during pregnancy. Am. J. Obstet. Gynecol. 2004, 190, 2–9. [Google Scholar] [CrossRef]
- Melland-Smith, M.; Ermini, L.; Chauvin, S.; Craig-Barnes, H.; Tagliaferro, A.; Todros, T.; Post, M.; Caniggia, I. Disruption of sphingolipid metabolism augments ceramide-induced autophagy in preeclampsia. Autophagy 2015, 11, 653–669. [Google Scholar] [CrossRef] [Green Version]
- Habayeb, O.M.; Taylor, A.H.; Evans, M.D.; Cooke, M.S.; Taylor, D.J.; Bell, S.C.; Konje, J.C. Plasma levels of the endocannabinoid anandamide in women—A potential role in pregnancy maintenance and labor? J. Clin. Endocrinol. Metab. 2004, 89, 5482–5487. [Google Scholar] [CrossRef]
- Signorelli, P.; Avagliano, L.; Reforgiato, M.R.; Toppi, N.; Casas, J.; Fabrias, G.; Marconi, A.M.; Ghidoni, R.; Caretti, A. De novo ceramide synthesis is involved in acute inflammation during labor. Biol. Chem. 2016, 397, 147–155. [Google Scholar] [CrossRef]
- Keirse, M.J.; Mitchell, M.D.; Turnbull, A.C. Changes in prostaglandin F and 13,14-dihydro-15-keto-prostaglandin F concentrations in amniotic fluid at the onset of and during labour. Br. J. Obstet. Gynaecol. 1977, 84, 743–746. [Google Scholar] [CrossRef]
- Sellers, S.M.; Hodgson, H.T.; Mitchell, M.D.; Anderson, A.B.; Turnbull, A.C. Raised prostaglandin levels in the third stage of labor. Am. J. Obstet. Gynecol. 1982, 144, 209–212. [Google Scholar] [CrossRef]
- Lee, D.C.; Romero, R.; Kim, J.S.; Yoo, W.; Lee, J.; Mittal, P.; Kusanovic, J.P.; Hassan, S.S.; Yoon, B.H.; Kim, C.J. Evidence for a spatial and temporal regulation of prostaglandin-endoperoxide synthase 2 expression in human amnion in term and preterm parturition. J. Clin. Endocrinol. Metab. 2010, 95, E86–E91. [Google Scholar] [CrossRef]
- Kurki, T.; Viinikka, L.; Ylikorkala, O. Urinary excretion of prostacyclin and thromboxane metabolites in threatened preterm labor: Effect of indomethacin and nylidrin. Am. J. Obstet. Gynecol. 1992, 166, 150–154. [Google Scholar] [CrossRef]
- Ylikorkala, O.; Paatero, H.; Suhonen, L.; Viinikka, L. Vaginal and abdominal delivery increases maternal urinary 6-keto-prostaglandin F1 alpha excretion. Br. J. Obstet. Gynaecol. 1986, 93, 950–954. [Google Scholar] [CrossRef] [PubMed]
- Hill, M.; Parizek, A.; Kancheva, R.; Duskova, M.; Velikova, M.; Kriz, L.; Klimkova, M.; Paskova, A.; Zizka, Z.; Matucha, P.; et al. Steroid metabolome in plasma from the umbilical artery, umbilical vein, maternal cubital vein and in amniotic fluid in normal and preterm labor. J. Steroid. Biochem. Mol. Biol. 2010, 121, 594–610. [Google Scholar] [CrossRef]
- Hill, M.; Parizek, A.; Cibula, D.; Kancheva, R.; Jirasek, J.E.; Jirkovska, M.; Velikova, M.; Kubatova, J.; Klimkova, M.; Paskova, A.; et al. Steroid metabolome in fetal and maternal body fluids in human late pregnancy. J. Steroid. Biochem. Mol. Biol. 2010, 122, 114–132. [Google Scholar] [CrossRef]
- Pavlicev, M.; Norwitz, E.R. Human Parturition: Nothing More Than a Delayed Menstruation. Reprod. Sci. 2018, 25, 166–173. [Google Scholar] [CrossRef]
- Phillips, R.J.; Heesom, K.J.; Trinder, J.; Bernal, A.L. Human maternal plasma proteomic changes with parturition. EuPA Open Proteom. 2014, 5, 10–20. [Google Scholar] [CrossRef] [Green Version]
- Diczfalusy, E. Endocrine functions of the human fetoplacental unit. 1964. Am. J. Obstet. Gynecol. 2005, 193, 2024–2025. [Google Scholar] [CrossRef]
- Rock, R.C. Endocrinology of the Fetoplacental Unit: Role of Estrogen Assays. Lab. Med. 1984, 15, 95–97. [Google Scholar] [CrossRef]
- Smith, R.; Butler, T.; Chan, E.-C. Do estrogen receptor variants explain the enigma of human birth? EBioMedicine 2018, 39, 25–26. [Google Scholar] [CrossRef]
- Smith, R.; Smith, J.I.; Shen, X.; Engel, P.J.; Bowman, M.E.; McGrath, S.A.; Bisits, A.M.; McElduff, P.; Giles, W.B.; Smith, D.W. Patterns of plasma corticotropin-releasing hormone, progesterone, estradiol, and estriol change and the onset of human labor. J. Clin. Endocrinol. Metab. 2009, 94, 2066–2074. [Google Scholar] [CrossRef]
- Hudson, C.A.; McArdle, C.A.; Lopez Bernal, A. Steroid receptor co-activator interacting protein (SIP) mediates EGF-stimulated expression of the prostaglandin synthase COX2 and prostaglandin release in human myometrium. Mol. Hum. Reprod. 2016, 22, 512–525. [Google Scholar] [CrossRef] [Green Version]
- Anamthathmakula, P.; Kyathanahalli, C.; Ingles, J.; Hassan, S.S.; Condon, J.C.; Jeyasuria, P. Estrogen receptor alpha isoform ERdelta7 in myometrium modulates uterine quiescence during pregnancy. EBioMedicine 2019, 39, 520–530. [Google Scholar] [CrossRef]


{kind=link}
| Demographic | VD n = 9 | Elective CS n = 10 | p Value (at ≤ 0.05) |
|---|---|---|---|
| Age (years) | 32.4 (24–39) a | 34.6 (30–40) a | NS |
| BMI at pregnancy booking | 21.6 (19.9–23.1) a | 25.56 (21.7–38.3) a | 0.02 |
| Ethnicity | White British: 9 | White British: 9 White Romanian: 1 | NS |
| Smoking history: | NS | ||
| Never | 8 | 4 | |
| Ex | 0 | 5 | |
| Current | 1 (4/day) | 1 (10/day) | |
| Maternal past medical history | Eczema: 1 ICSI pregnancy (own sperm/egg): 1 | Previous postnatal depression:1 Well controlled asthma: 1 (MTHFR gene 1): 1 | NA |
| Maternal employment status at pregnancy booking: | NS | ||
| Employed | 8 | 8 | |
| Home maker | 1 | 2 | |
| Gravida | 1.2 (0–2) a | 2.4 (0–5) a | NS |
| Parity | 0.6 (0–2) a | 1.7 (0–7) a | NS |
| Significant pregnancy complications | Nil | Nil | NA |
| Indication for elective CS | NA | Breech: 3 Previous CS: 3 Maternal request: 1 Previous 3rd degree vaginal tear: 1 Previous traumatic delivery: 1 Tocophobia: 1 | NA |
| Duration of labour in minutes | 312 min (70–650) a | NA | NA |
| Gestation at delivery (weeks+days) | 40 (38+1–41+6) a | 39+2 (38+4–40+3) a | 0.04 |
| Apgar scores at 1, 5 and 10 min: | NS | ||
| 8,9,10 | 1 | 0 | |
| 9,10,10 | 8 | 10 | |
| Management of third stage: | NS | ||
| Syntometrine or Carbetocin | 8 | 10 | |
| Physiological | 1 | 0 | |
| Interval between delivery and freezing of sample (minutes) | 38 (36–60) b | 28.5 (24–45) b | 0.02 |
| Gender of fetus: | NS | ||
| Female | 5 | 4 | |
| Male | 4 | 6 | |
| Apgar scores at 1, 5 and 10 min: | NS | ||
| 8,9,10 | 1 | 0 | |
| 9,10,10 | 8 | 10 | |
| Birth weight (Kilograms): | 3.4 (3.2–3.9) a | 3.6 (3.1–4.1) a | NS |
| Super Pathway | Number Metabolites Measured | Number Significant Changes VM/EM | Number Significant Changes VC/EC | ||||
|---|---|---|---|---|---|---|---|
| ↑ | ↓ | Total (%) | ↑ | ↓ | Total (%) | ||
| Lipid | 405 | 139 | 14 | 153 (37.8) | 73 | 12 | 85 (21.0) |
| Amino Acid | 177 | 9 | 15 | 24 (13.6) | 15 | 14 | 29 (16.4) |
| Xenobiotics | 108 | 18 | 3 | 21 (19.4) | 16 | 5 | 21 (19.4) |
| Carbohydrate | 29 | 7 | 0 | 7 (24.1) | 9 | 0 | 9 (31.0) |
| Cofactors and Vitamins | 31 | 3 | 4 | 7 (22.6) | 3 | 5 | 8 (25.8) |
| Energy | 11 | 3 | 0 | 3 (27.3) | 6 | 0 | 6 (54.6) |
| Nucleotide | 38 | 2 | 2 | 4 (10.5) | 9 | 3 | 12 (31.6) |
| Peptide | 24 | 0 | 1 | 1 (4.2) | 3 | 5 | 8 (33.3) |
| Partially characterised | 3 | 2 | 0 | 2 | 0 | ||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Birchenall, K.A.; Welsh, G.I.; López Bernal, A. Metabolite Changes in Maternal and Fetal Plasma Following Spontaneous Labour at Term in Humans Using Untargeted Metabolomics Analysis: A Pilot Study. Int. J. Environ. Res. Public Health 2019, 16, 1527. https://doi.org/10.3390/ijerph16091527
Birchenall KA, Welsh GI, López Bernal A. Metabolite Changes in Maternal and Fetal Plasma Following Spontaneous Labour at Term in Humans Using Untargeted Metabolomics Analysis: A Pilot Study. International Journal of Environmental Research and Public Health. 2019; 16(9):1527. https://doi.org/10.3390/ijerph16091527
Chicago/Turabian StyleBirchenall, Katherine A., Gavin I. Welsh, and Andrés López Bernal. 2019. "Metabolite Changes in Maternal and Fetal Plasma Following Spontaneous Labour at Term in Humans Using Untargeted Metabolomics Analysis: A Pilot Study" International Journal of Environmental Research and Public Health 16, no. 9: 1527. https://doi.org/10.3390/ijerph16091527
APA StyleBirchenall, K. A., Welsh, G. I., & López Bernal, A. (2019). Metabolite Changes in Maternal and Fetal Plasma Following Spontaneous Labour at Term in Humans Using Untargeted Metabolomics Analysis: A Pilot Study. International Journal of Environmental Research and Public Health, 16(9), 1527. https://doi.org/10.3390/ijerph16091527