Critical Review on the Presence of Phthalates in Food and Evidence of Their Biological Impact


 and
and 
Abstract
:
1. Introduction
2. Physical-Chemical Properties and Applications
3. Toxicological Aspects and Human Health Effects
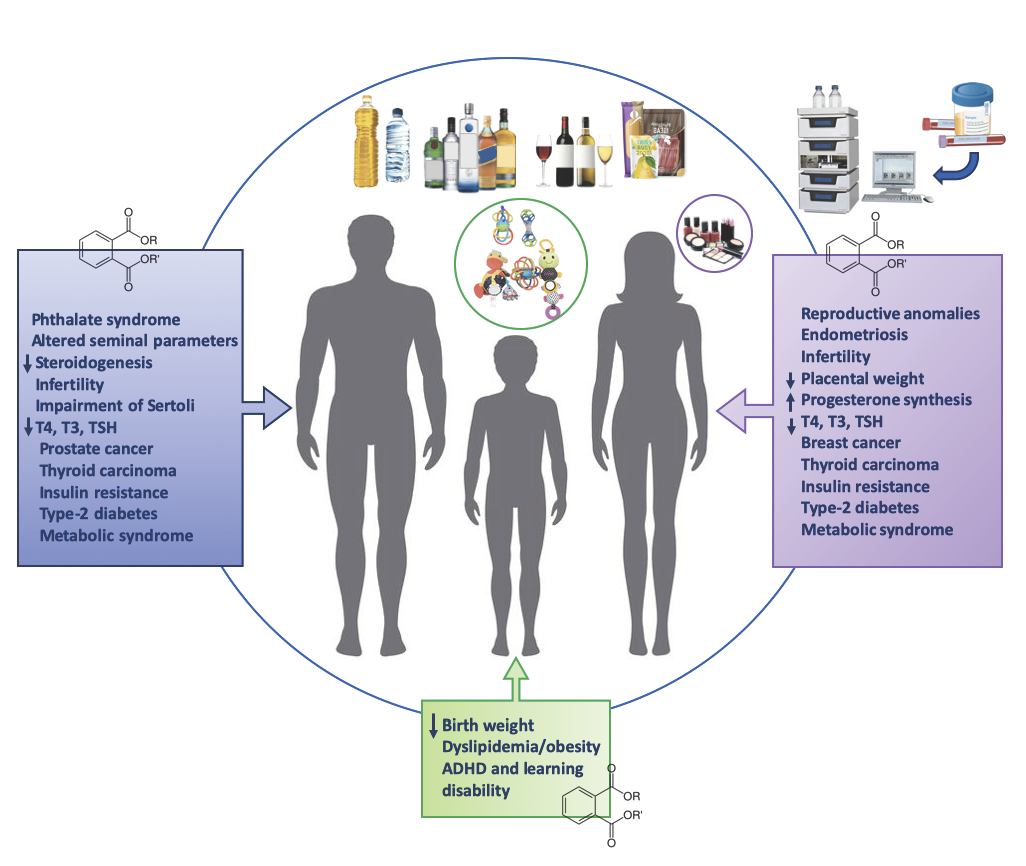
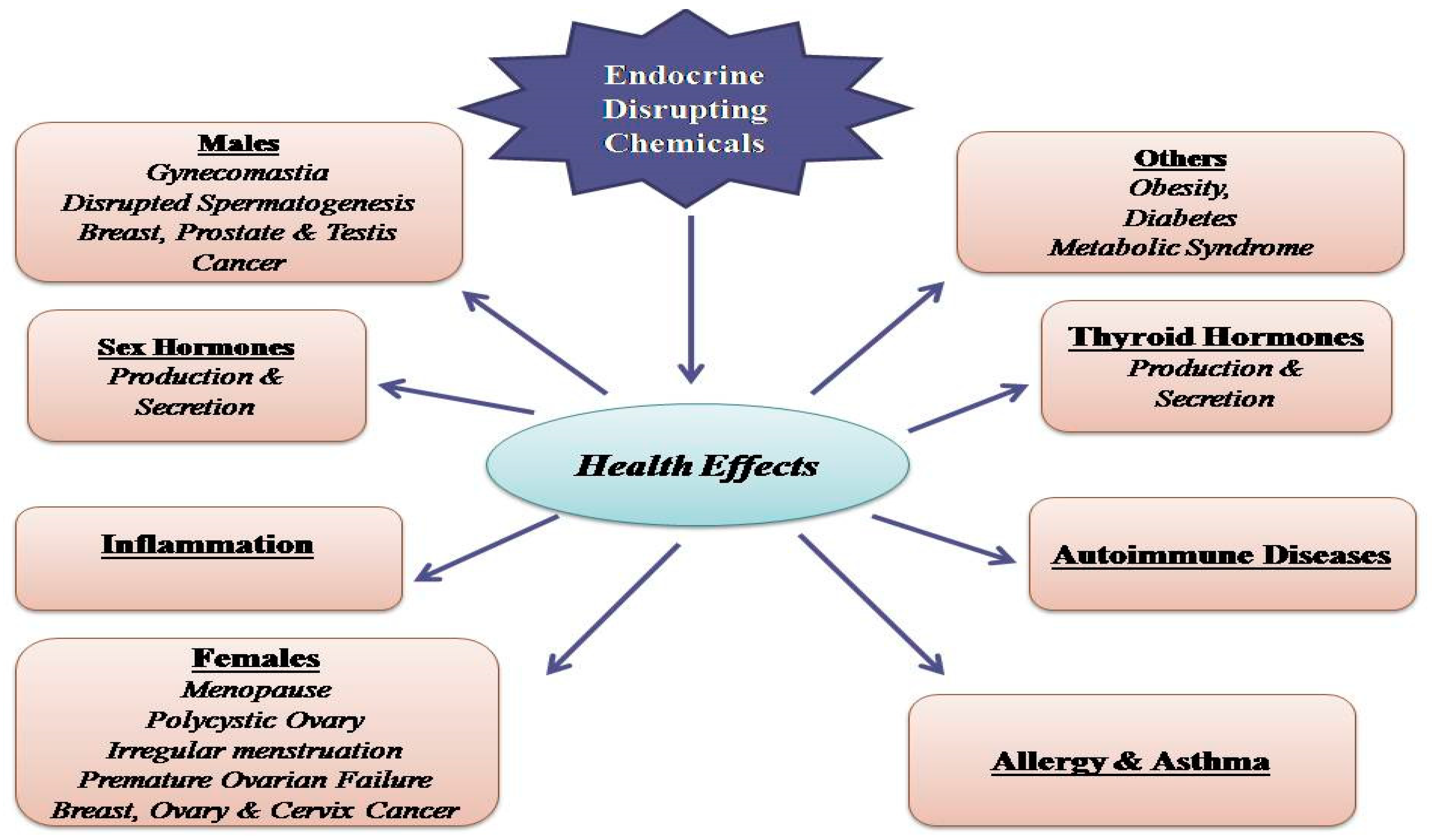
3.1. Phthalates and Endocrine Toxicity
3.2. Phthalates and Cancer
3.3. Phthalates and Metabolic Disorders
3.4. Phthalates and Neurotoxicity
3.5. Phthalates and Immune System
4. Biochemical Regulation of Phthalates Effects
5. Phthalate Regulations
- A: Ethanol 10%, simulates hydrophilic foods,
- B: Acetic Acid 3%, simulates hydrophilic foods with pH < 4.5,
- C: Ethanol 20%, simulates hydrophilic foods with alcohol < 20%,
- D1: Ethanol 50%, simulates lipophilic materials, foods with alcoholic contents > 20%, and oil and water in oil emulsions.
- D2: Vegetable oil and lipophilic foods.
- E: Tenax and dry foods.
6. Occurrence of PAEs in Food
6.1. Alcoholic Beverages
6.2. Mineral Water
6.3. Soft Drinks

{kind=link}
{kind=link}
| Matrix | Phthalate | Average Concentration | Reference |
|---|---|---|---|
| Water in PET bottles | DEHP | 0.196 | [213] |
| DBP | 0.046 | ||
| DEP | 0.432 | ||
| Mineral water | DMP | ND | [217] |
| DBP | 11.33 | ||
| DEHP | 8.79 | ||
| Soft drink K sorbate | DMP | 759.80 | |
| DBP | 9.00 | ||
| DEHP | 36.60 | ||
| Soft drink K sorbate and Na benzoate | DMP | 500.88 | |
| DBP | 26.75 | ||
| DEHP | 15 | ||
| Water in PET bottles | DBP | 0.21 | [212] |
| DEP | 0.17 | ||
| DEHP | 0.02 | ||
| Bottled water | DEHP | 0.35 | [203] |
| DBP | 0.044 | ||
| DEP | 0.033 | ||
| Water in PET bottles, non-carbonated | DEHP | 0.016–1.7 | [210] |
| DBP | 0.007–0.08 | ||
| DIBP | 0.003–0.02 | ||
| Water in PET bottles | DEHP | 0.217 | [17] |
| DBP | 0.135 | ||
| Room temperature | DEHP | 0.411 | |
| DBP | 0.116 | ||
| Refrigerator | DEHP | 0.423 | |
| DBP | 0.124 | ||
| Freezing | DEHP | 0.317 | |
| DBP | 0.079 | ||
| 40 °C | DEHP | 0.917 | |
| DBP | 0.303 | ||
| Mineral water | DEHP | 248 | [218] |
| Orange flavored soda | DMP | 74 | |
| DEP | 91 | ||
| Cola | DMP | 105 | |
| DEHP | 1123 | ||
| Sport drinks | DEHP | 15–98 | [219] |
| Tea | DEHP | 16–1263 | |
| Coffee | DEHP | 28–159 | |
| Fruit juices | DEHP | 22–126 | |
| Espresso coffee surrogates from pre-packed metal capsule | DEHP | 220 | [1] |
| DIBP | 240 | ||
| DEP | 230 | ||
| DBP | 4 | ||
| Espresso coffee surrogates from pre-packed plastic capsule | DEHP | 1560 | |
| DIBP | 7 | ||
| DBP | 7 | ||
| Espresso coffee surrogates from pre-packed biodegradable capsule | DEHP | 830 | |
| DIBP | 330 | ||
| DBP | 120 |
6.4. Edible Oils and Fats
6.5. Dairy Products
6.6. Meat and Poultry
6.7. Edible Plants
7. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- De Toni, L.; Tisato, F.; Seraglia, R.; Roverso, M.; Gandin, V.; Marzano, C.; Padrini, R.; Foresta, C. Phthalates and heavy metals as endocrine disruptors in food: A study on pre-packed coffee products. Toxicol. Rep. 2017, 4, 234–239. [Google Scholar] [CrossRef]
- Schettler, T. Human exposure to phthalates via consumer products. Int. J. Androl. 2006, 29, 134–139. [Google Scholar] [CrossRef]
- Fang, H.; Wang, J.; Lynch, A.R. Migration of di(2-ethylhexyl)phthalate (DEHP) and di-n-butylphthalate (DBP) from polypropylene food containers. Food Control. 2016, 73, 1298–1302. [Google Scholar] [CrossRef]
- Net, S.; Sempéré, R.; Delmont, A.; Paluselli, A.; Ouddane, B. Occurrence, fate, behavior and ecotoxicological state of phthalates in different environmental matrices. Environ. Sci. Tech. 2015, 49, 4019–4035. [Google Scholar] [CrossRef]
- Pan, T.L.; Wang, P.W.; Aljuffali, I.A.; Hung, Y.Y.; Lin, C.F.; Fang, J.Y. Dermal toxicity elicited by phthalates: Evaluation of skin absorption, immunohistology, and functional proteomics. Food Chem. Toxicol. 2014, 65, 105–114. [Google Scholar] [CrossRef]
- Katsikantami, I.; Tzatzarakis, M.N.; Karzi, V.; Stavroulaki, A.; Xezonaki, P.; Vakonaki, E.; Alegakis, A.K.; Sifakis, S.; Rizos, A.K.; Tsatsakis, A.M. Biomonitoring of bisphenols A and S and phthalate metabolites in hair from pregnant women in Crete. Sci. Total Environ. 2020, 712, 135651. [Google Scholar] [CrossRef]
- Yang, J.; Li, Y.; Wang, Y.; Ruan, J.; Zhang, J.; Sun, C. Recent advances in analysis of phthalate esters in foods. TrAC Trends in Analy. Chemistry 2015, 72, 10–26. [Google Scholar]
- He, M.; Yang, C.; Geng, R.; Zhao, X.; Hong, L.; Piao, X.; Chen, T.; Quinto, M.; Li, D. Monitoring of phthalates in foodstuffs using gas purge microsyringe extraction coupled with GC–MS. Anal. Chim. Acta 2015, 879, 63–68. [Google Scholar] [CrossRef]
- Notardonato, I.; Salimei, E.; Russo, M.V.; Avino, P. Simultaneous determination of organophosphorus pesticides and phthalates in baby food samples by ultrasound-vortex-assisted liquid-liquid microextraction and GC-IT/MS. Anal. Bioanal. Chem. 2018, 410, 3285–3296. [Google Scholar] [CrossRef] [PubMed]
- Russo, M.V.; Avino, P.; Notardonato, I. Fast analysis of phthalates in freeze-dried baby foods by ultrasound-vortex-assisted liquid-liquid microextraction coupled with gas chromatography-ion trap/mass spectrometry. J. Chromatogr. A 2016, 25, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Russo, M.V.; Avino, P.; Perugini, L.; Notardonato, I. Extraction and GC-MS analysis of phthalate esters in food matrices: A review. RSC Adv. 2015, 5, 37023–37043. [Google Scholar] [CrossRef]
- Russo, M.V.; Notardonato, I.; Cinelli, G.; Avino, P. Evaluation of an analytical method for determining phthalate esters in wine samples by solid-phase extraction and gas chromatography coupled with ion-trap mass spectrometer detector. Anal. Bioanal. Chem. 2012, 402, 1373–1381. [Google Scholar] [CrossRef] [PubMed]
- Russo, M.V.; Notardonato, I.; Avino, P.; Cinelli, G. Determination of phthalate esters at trace levels in light alcoholic drinks and soft drinks by XAD-2 adsorbent and gas chromatography coupled with ion trap-mass spectrometry detection. Anal. Methods-UK 2014, 6, 7030–7037. [Google Scholar] [CrossRef]
- Notardonato, I.; Protano, C.; Vitali, M.; Bhattacharya, B.; Avino, P. A method validation for simultaneous determination of phthalates and bisphenol A released from plastic water containers. Appl. Sci. 2019, 9, 2945. [Google Scholar] [CrossRef] [Green Version]
- Notardonato, I.; Protano, C.; Vitali, M.; Avino, P. Phthalates and bisphenol-A determination and release from different beverage plastic containers by dispersive liquid-liquid microextraction and GC-IT/MS analysis. Food Anal. Methods 2019, 12, 2562–2571. [Google Scholar] [CrossRef]
- Notardonato, I.; Passarella, S.; Ianiri, G.; Di Fiore, C.; Russo, M.V.; Avino, P. Analytical Method Development and Chemometric Approach for Evidencing Presence of Plasticizer Residues in Nectar Honey Samples. Int. J. Environ. Res. Public Health 2020, 17, 1692. [Google Scholar] [CrossRef] [Green Version]
- Jeddi, M.Z.; Rastkari, N.; Ahmadkhaniha, R.; Yunesian, M. Concentrations of phthalates in bottled water under common storage conditions: Do they pose a health risk to children? Food Res. Int. 2015, 69, 256–265. [Google Scholar] [CrossRef]
- López-Carrillo, L.; Hernández, R.R.U.; Calafat, A.M.; Torres, S.L.; Galván, P.M.; Needham, L.L.; Ruiz, R.R.; Cebrián, M.E. Exposure to phthalates and breast cancer risk in northern Mexico. Environ. Health Perspect. 2010, 118, 539–544. [Google Scholar]
- Mu, D.; Gao, F.; Fan, Z.; Shen, H.; Peng, H.; Hu, J. Levels of phthalate metabolites in urine of pregnant women and risk of clinical pregnancy loss. Environ. Sci. Technol. Lett. 2015, 49, 10651–10657. [Google Scholar] [CrossRef]
- Petrović, E.K. How substances get regulated against in the building industry: Formaldehyde, phthalate plasticizers in polyvinyl chloride/vinyl. In Materials for a Healthy, Ecological and Sustainable Built Environment; Woodhead Publishing: Swaston, UK, 2017; pp. 175–189. [Google Scholar]
- Harunarashid, N.Z.I.H.; Lim, L.H.; Harunsani, M.H. Phthalate sample preparation methods and analysis in food and food packaging: A review. Food Anal. Methods 2017, 10, 3790–3814. [Google Scholar] [CrossRef]
- Wypych, A. Databook of Plasticizers, 2nd ed.; ChemTec Publishing: Scarborough, ON, Canada, 2017; pp. 435–560. [Google Scholar]
- Chiellini, F.; Ferri, M.; Morelli, A.; Dipaola, L.; Latini, G. Perspectives on alternatives to phthalate plasticized poly(vinyl chloride) in medical devices applications. Prog. Polym. Sci. 2013, 38, 1067–1088. [Google Scholar] [CrossRef] [Green Version]
- Casas-Hernàndez, A.M.; Paz, A.C.M.; Gómez, H.A. Application of time-resolved luminescence methodology to the determination of phthalate esters. Anal. Lett. 2003, 36, 1017–1027. [Google Scholar] [CrossRef]
- Huang, P.C.; Tien, C.J.; Sun, Y.M.; Hsieh, C.Y.; Lee, C.C. Occurrence of phthalates in sediment and biota: Relationship to aquatic factors and the biota-sediment accumulation factor. Chemosphere 2008, 73, 539–544. [Google Scholar] [CrossRef] [PubMed]
- Staples, C.A.; Peterson, D.R.; Parkerton, T.F.; Adams, W.J. The environmental fate of phthalate esters: A literature review. Chemosphere 1997, 35, 667–749. [Google Scholar] [CrossRef]
- Vikelsøe, J.; Thomsen, M.; Carlsen, L. Phthalates and nonylphenols in profiles of differently dressed soils. Sci. Total Environ. 2002, 296, 105–116. [Google Scholar] [CrossRef]
- Zheng, X.; Zhang, B.T.; Teng, Y. Distribution of phthalate acid esters in lakes of Beijing and its relationship with anthropogenic activities. Sci. Total Environ. 2014, 476, 107–113. [Google Scholar] [CrossRef] [PubMed]
- Hammer, J.; Tukker, A.M.; Postma, J.F.; Haftka, J.J.H.; Hermens, J.L.M.; De, V.P.; Kraak, M.H.S. Solubility Constraints on Aquatic Ecotoxicity Testing of Anionic Surfactants. Bull. Environ. Contam. Toxicol. 2018, 101, 99–104. [Google Scholar] [CrossRef] [Green Version]
- Cousins, A.; Remberger, M.; Kaj, L.; Ekheden, Y.; Dusan, B.; Brorström, L.E. Results from the Swedish National Screening Programme 2006: Subreport 1: Phthalates; IVL Swedish Environmental Research Institute: Stockholm, Sweeden, 2007. [Google Scholar]
- Repouskou, A.; Panagiotidou, E.; Panagopoulou, L.; Bisting, P.L.; Tuck, A.R.; Sjödin, M.O.D.; Lindberg, J.; Bozas, E.; Rüegg, J.; Gennings, C.; et al. Gestational exposure to an epidemiologically defined mixture of phthalates leads to gonadal dysfunction in mouse offspring of both sexes. Sci. Rep. 2019, 9, 6424. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Wang, W.; Zhu, J.; Li, Y.; Luo, L.; Huang, Y.; Zhang, W. Di(2-ethylhexyl) phthalate (DEHP) influences follicular development in mice between the weaning period and maturity by interfering with ovarian development factors and microRNAs. Environ. Toxicol. 2018, 33, 535–544. [Google Scholar] [CrossRef]
- Casals-Casas, C.; Desvergne, B. Endocrine disruptors: From endocrine to metabolic disruption. Annu. Rev. Physiol. 2011, 73, 135–162. [Google Scholar] [CrossRef] [Green Version]
- Hatch, E.E.; Nelson, J.W.; Qureshi, M.M.; Weinberg, J.; Moore, L.L.; Singer, M.; Webster, T.F. Association of urinary phthalate metabolite concentrations with body mass index and waist circumference: A cross-sectional study of NHANES data, 1999–2002. Environ. Health 2008, 7, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muscogiuri, G.A.A.C. Phthalates: New cardiovascular health disruptors? Arch. Toxicol. 2017, 91, 1513–1517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ventrice, P.; Ventrice, D.; Russo, E.; De, S.G. Phthalates: European regulation, chemistry, pharmacokinetic and related toxicity. Envt. Toxic. Pharmc. 2013, 36, 88–96. [Google Scholar] [CrossRef] [PubMed]
- Jaakkola, J.J.; Knight, T.L. The role of exposure to phthalates from polyvinyl chloride products in the development of asthma and allergies: A systematic review and meta-analysis. Environ. Health Perspect. 2008, 116, 845–853. [Google Scholar] [CrossRef]
- Genuis, S.J.; Beesoon, S.; Lobo, R.A.; Birkholz, D. Human elimination of phthalate compounds: Blood, urine, and sweat (BUS) study. Sci. World J. 2012, 2012, 615068. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Zhu, H.; Kannan, K. A review of biomonitoring of phthalate exposures. Toxics 2019, 7, 21. [Google Scholar] [CrossRef] [Green Version]
- Weiss, J.M.; Gustafsson, Å.; Gerde, P.; Bergman, Å.; Lindh, C.H.; Krais, A.M. Daily intake of phthalates, MEHP, and DINCH by ingestion and inhalation. Chemosphere 2018, 208, 40–49. [Google Scholar] [CrossRef]
- Wittassek, M.; Angerer, J. Phthalates: Metabolism and exposure. Int. J. Androl. 2008, 31, 131–138. [Google Scholar] [CrossRef]
- Minguez-Alarcon, L.W.; Chiu, Y.H.; Gaskins, A.J.; Nassan, F.L.; Dadd, R.; Petrozza, J.; Hauser, R.; Chavarro, J.E. Secular trends in semen parameters among men attending a fertility center between 2000 and 2017, Identifying potential predictors. Environ. Int. 2018, 121, 1297–1303. [Google Scholar] [CrossRef]
- Meehan, T.; Schlatt, S.; O’Bryan, M.K. Regulation of germ cell and Sertoli cell development byactivin, follistatin, and FSH. Dev. Biol. 2000, 220, 225–237. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.X.; Zhou, B.; Chen, Y.-J.; Liu, C.; Huang, L.-L.; Liao, J.-Q.; Hu, X.-J.; Lu, W.-Q.; Zeng, Q.; Pan, A. Thyroid function, phthalate exposure and semen quality: Exploring associations and mediation effects in reproductive-aged men. Environ. Int. 2018, 116, 278–285. [Google Scholar] [CrossRef] [PubMed]
- Willems, A.; De, G.K.; Allemeersch, J.; Smith, L.B.; Welsh, M.; Swinnen, J.V.; Verhoeven, G. Early effects of Sertoli cell-selective androgen receptor ablation on testicular gene expression. Int. J. Androl. 2010, 33, 507–517. [Google Scholar] [CrossRef] [PubMed]
- Caporossi, L.; Alteri, A.; Campo, G.; Paci, E.; Tranfo, G.; Capanna, S.; Papaleo, E.; Pigini, D.; Viganò, P.; Papaleo, B. Cross Sectional Study on Exposure to BPA and Phthalates and Semen Parameters in Men Attending a Fertility Center. Int. J. Environ. Res. Pub. Health 2020, 17, 489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Hond, E.T.H.; De, S.P.; Ombelet, W.; Baeyens, W.; Covaci, A.; Cox, B.; Nawrot, T.S.; Larebeke, N.V.; D’Hooghe, T. Human exposure to endocrine disrupting chemicals and fertility: A case control study in male subfertility patients. Environ. Int. 2015, 84, 154–160. [Google Scholar] [CrossRef]
- Botelho, G.; Golin, M.; Bufalo, A.; Morais, R.; Dalsenter, P.; Martino-Andrade, A. Reproductive effects of di(2-ethylhexyl)phthalate in immature male rats and its relation to cholesterol, testosterone, and thyroxin levels. Arch. Environ. Contam. Toxicol. 2009, 57, 777–784. [Google Scholar] [CrossRef]
- Ghisari, M.; Bonefeld-Jorgensen, E.C. Effects of plasticizers and their mixtures on estrogen receptor and thyroid hormone functions. Toxicol. Lett. 2009, 189, 67–77. [Google Scholar] [CrossRef]
- Sheikh, I.A. Stereoselectivity and the potential endocrine disrupting activity of di-(2-ethylhexyl) phthalate (DEHP) against human progesterone receptor: A computational perspective. J. Appl. Toxicol. 2016, 36, 741–747. [Google Scholar] [CrossRef]
- Sharpe, R.M.; Skakkebaek, N.E. Testicular dysgenesis syndrome: Mechanistic insights and potential new downstream effects. Fertil. Steril. 2008, 89, 33–38. [Google Scholar] [CrossRef]
- Pocar, P.; Fiandanese, N.; Secchi, C.; Berrini, A.; Fischer, B.; Schmidt, J.S.; Schaedlich, K.; Borromeo, V. Exposure to di (2-ethyl-hexyl) phthalate (DEHP) in utero and during lactation causes long-term pituitary-gonadal axis disruption in male and female mouse offspring. Endocrinology 2011, 153, 937–948. [Google Scholar] [CrossRef] [Green Version]
- Sobarzo, C.M.; Rosana, N.D.M.; Livia, L.; Berta, D.; Schteingart, H.F. Mono-(2-ethylhexyl) phthalate (MEHP) affects intercellular junctions of Sertoli cell: A potential role of oxidative stress. Reprod. Toxicol. 2015, 58, 203–212. [Google Scholar] [CrossRef]
- Ge, R.S.; Chen, G.R.; Tanrikut, C.; Hardy, M.P. Phthalate ester toxicity in Leydig cells: Developmental timing and dosage considerations. Reprod. Toxicol. 2007, 23, 366–373. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Fang, E.F.; Scheibye-Knudsen, M.; Cui, H.; Qiu, L.; Li, J.; He, Y.; Huang, J.; Bohr, V.A.; Ng, T.B.; et al. Di-(2 ethylhexyl) phthalate inhibits DNA replication leading to hyperPARylation, SIRT1 attenuation, and mitochondrial dysfunction in the testis. Sci. Rep. 2014, 4, 6434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aly, H.A.; Hassan, M.H.; El-Beshbishy, H.A.; Alahdal, A.M.; Osman, A.M. Dibutylphthalate induces oxidative stress and impairs spermatogenesis in adult rats. Toxicol. Ind. Health 2016, 32, 1467–1477. [Google Scholar] [CrossRef]
- Lenzi, A.; Gandini, L.; Maresca, V.; Rago, R.; Sgrò, P.; Dondero, F.; Picardo, M. Fatty acid composition of spermatozoa and immature germ cells. Mol. Human Reprod. 2000, 6, 226–231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amaral, A.; Lourenço, B.; Marques, M.; Ramalho-Santos, J. Mitochondria functionality and sperm quality. Reproduction 2013, 146, 163–174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, G.X.; Lian, Q.Q.; Ge, R.S.; Hardy, D.O.; Li, X.K. Phthalate-induced testicular dysgenesis syndrome: Leydig cell influence. Trends Endocrinol. Metab. TEM 2009, 20, 139–145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Saleh, I.; Coskun, S.; Al-Doush, I.; Al-Rajudi, T.; Abduljabbar, M.; Al-Rouqi, R.; Palawan, H.; Al-Hassan, S. The relationships between urinary phthalate metabolites, reproductive hormones and semen parameters in men attending in vitro fertilization clinics. Sci. Total Environ. 2019, 658, 982–995. [Google Scholar] [CrossRef]
- Singh, S.L.; Li, S.S.-L. Epigenetic Effects of Environmental Chemicals Bisphenol and Phthalates. Int. J. Mol. Sci. 2012, 13, 10143–10153. [Google Scholar] [CrossRef]
- Tian, M.L.L.; Zhang, J.; Huang, Q.; Shen, H. Positive Association of Low-level Environmental Phthalate Exposure with Sperm Motility was Mediated by DNA Methylation: A Pilot Study. Chemosphere 2019, 220, 459–467. [Google Scholar] [CrossRef]
- Houshdaran, S.; Cortessis, V.; Siegmund, K.; Yang, A.; Laird, P.W.; Sokol, R.Z. Widespread epigenetic abnormalities suggest a broad DNA methylation erasure defect in abnormal human sperm. PLoS ONE 2007, 2, e1289. [Google Scholar] [CrossRef] [Green Version]
- Rahiminia, T.; Yazd, E.F.; Fesahat, F.; Moein, M.; Mirjalili, A.; Talebi, R. Sperm chromatin and DNA integrity, methyltransferase mRNA levels, and global DNA methylation in oligoasthenoteratozoospermia. Clin. Exp. Reprod. Med. 2018, 45, 17–24. [Google Scholar] [CrossRef] [PubMed]
- Santi, D.; De Vincentis, S.; Magnani, E.; Spaggiari, G. Impairment of sperm DNA methylation in male infertility: A meta-analytic study. Andrology 2017, 5, 695–703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valinluck, V.; Tsai, H.H.; Rogstad, D.K.; Burdzy, A.; Bird, A.; Sowers, L.C. Oxidative damage to methyl-CpG sequences inhibits the binding of the methyl-CpG binding domain (MBD) of methyl-CpG binding protein 2 (MeCP2). Nucleic Acids Res. 2004, 32, 4100–4108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martinez-Arguelles, D.; Culty, M.; Zirkin, B.R.; Papadopoulos, V. In utero exposure to di-(2-ethylhexyl) phthalate decreases mineralocorticoid receptor expression in the adult testis. Endocrinology 2009, 150, 5575–5585. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kariyazono, Y.; Taura, J.; Hattori, Y.; Ishii, Y.; Narimatsu, S.; Fujimura, M.; Takeda, T.; Yamada, H. Effect of in utero exposure to endocrine disruptors on fetal steroidogenesis governed by the pituitary-gonad axis: A study in rats using different ways of administration. J. Toxic. Sci. 2015, 40, 909–916. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herr, C.; Zur, N.A.; Koch, H.M.; Schuppe, H.C.; Fieber, C.; Angerer, J.; Eikmann, T.; Stilianakis, N.I. Urinary di (2-ethylhexyl) phthalate (DEHP)-metabolites and male human markers of reproductive function. Int. J. Hyg. Environ. Health 2009, 212, 648–653. [Google Scholar] [CrossRef]
- Strakovsky, R.S.S.; Schantz, S.L. Impacts of bisphenol A (BPA) and phthalate exposures on epigenetic outcomes in the human placenta. Environ. Epigenet. 2018, 4, dvy022. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.W.F.; Zhang, J.; Hao, L.; Jiang, J.; Dang, L.; Mei, D.; Fan, S.S.; Yu, Y.; Jiang, L. Bisphenol A and estrogen induce proliferation of human thyroid tumor cells via an estrogen-receptor-dependent pathway. Biochem. Biophys. 2017, 633, 29–39. [Google Scholar] [CrossRef]
- Long, M.G.M.; Kjeldsen, L.; Wielsøe, M.; Nørgaard, P.B.; Mortensen, E.L.; Abdallah, M.W.; Bonefeld, J.E.C. Autism spectrum disorders, endocrine disrupting compounds, and heavy metals in amniotic fluid: A case-control study. Mol. Autism 2019, 10, 1. [Google Scholar] [CrossRef]
- Latini, G.; De Felice, C.; Presta, G.; Del Vecchio, A.; Paris, I.; Ruggieri, F.; Mazzeo, P. In utero exposure to di-(2-ethylhexyl) phthalate and duration of human pregnancy. Environ. Health Perspect. 2003, 111, 1783–1785. [Google Scholar] [CrossRef] [Green Version]
- Agarwal, A.; Gupta, S.; Sharma, R.K. Role of oxidative stress in female reproduction. Reprod. Biol. Endocrin. 2005, 3, 28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Upson, K.; Sathyanarayana, S.; De Roos, A.J.; Thompson, M.L.; Scholes, D.; Dills, R.; Holt, V.L. Phthalates and risk of endometriosis. Environ. Res. 2013, 126, 91–97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Absalan, F.; Saremy, S.; Mansouri, E.; Moghadam, M.T.; Moghadam, A.R.E.; Ghanavati, R. Effects of mono-(2-ethylhexyl) phthalate and Di-(2-ethylhexyl) phthalate administrations on oocyte meiotic maturation, apoptosis and gene quantification in mouse model. Cell 2016, 18, 503–513. [Google Scholar]
- Frederiksen, H.; Sørensen, K.; Mouritsen, A.; Aksglaede, L.; Hagen, C.P.; Petersen, J.H.; Skakkebaek, N.E.; Andersson, A.M.; Juul, A. High urinary phthalate concentration associated with delayed pubarche in girls. Int. J. Androl. 2012, 35, 216–226. [Google Scholar] [CrossRef]
- Hauser, R.; Gaskins, A.J.; Souter, I.; Smith, K.W.; Dodge, L.E.; Ehrlich, S.; Meeker, J.D.; Calafat, A.M.; Williams, P.L. Urinary phthalate metabolite concentrations and reproductive outcomes among women undergoing in vitro fertilization: Results from the EARTH study. Environ. Health Perspect 2016, 124, 831–839. [Google Scholar] [CrossRef] [Green Version]
- Shen, R.; Zhao, L.L.; Yu, Z.; Zhang, C.; Chen, Y.H.; Wang, H.; Zhang, Z.H.; Xu, D.X. Maternal di-(2-ethylhexyl) phthalate exposure during pregnancy causes fetal growth restriction in a stage-specific but gender-independent manner. Reprod. Toxicol. 2017, 67, 117–124. [Google Scholar] [CrossRef]
- Zhang, S.; Sun, C.; Zhao, S.; Wang, B.; Wang, H.; Zhang, J.; Wang, Y.; Cheng, H.; Zhu, L.; Shen, R.; et al. Exposure to DEHP or its metabolite MEHP promotes progesterone secretion and inhibits proliferation in mouse placenta or JEG-3 cells. Environ. Pollut. 2020, 257, 113593. [Google Scholar] [CrossRef]
- Mose, T.; Mortensen, G.K.; Hedegaard, M.; Knudsen, L.E. Phthalate monoesters in perfusate from a dual placenta perfusion system, the placenta tissue and umbilical cord blood. Reprod. Toxicol. 2007, 23, 83–91. [Google Scholar] [CrossRef]
- Silva, M.J.; Reidy, J.A.; Herbert, A.R.; Preau, J.L.; Needham, L.L.; Calafat, A.M. Detection of phthalate metabolites in human amniotic fluid. Bull. Environ. Contam. Toxicol. 2004, 72, 1226–1231. [Google Scholar] [CrossRef]
- Petit, J.; Wakx, A.; Gil, S.; Fournier, T.; Auzeil, N.; Rat, P.; Laprevote, O. Lipidome-wide disturbances of human placental JEG-3 cells by the presence of MEHP. Biochimie 2018, 149, 1–8. [Google Scholar] [CrossRef]
- Main, K.M.; Mortensen, G.K.; Kaleva, M.; A Boisen, K.; Damgaard, I.N.; Chellakooty, M.; Schmidt, I.M.; Suomi, A.-M.; Virtanen, H.E.; Petersen, D.V.H.; et al. Human breast milk contamination with phthalates and alterations of endogenous reproductive hormones in infants three months of age. Environ. Health Perspect. 2006, B114, 270–276. [Google Scholar] [CrossRef] [PubMed]
- Hannon, P.R.; Brannick, K.E.; Wang, W.; Flaws, J.A. Mono(2-ethylhexyl) phthalate accelerates early folliculogenesis and inhibits steroidogenesis in cultured mouse whole ovaries andantralfollicles. Biol. Reprod. 2015, 92, 120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keuchi, S.; Iida, M.; Kobayashi, S.; Jin, K.; Matsuda, T.; Kojima, H. Differential effects of phthalate esters on transcriptional activities via human estrogen receptors alpha and beta, and androgen receptor. Toxicology 2005, 210, 223–233. [Google Scholar]
- Gupta, R.K.; Singh, J.M.; Leslie, T.C.; Meachum, S.; Flaws, J.A.; Yao, H.H. Di-(2-ethylhexyl) phthalate and mono-(2- ethylhexyl) phthalate inhibit growth and reduce estradiol levels of antral follicles in vitro. Toxicol. Appl. Pharm. 2010, 242, 224–230. [Google Scholar] [CrossRef] [Green Version]
- Lovekamp, S.T.; Davis, B.J. Mechanisms of phthalate ester toxicity in the female reproductive system. Environ. Health Perspect. 2003, 111, 139–145. [Google Scholar] [CrossRef] [Green Version]
- Shanle, E.K.; Xu, W. Endocrine disrupting chemicals targeting estrogen receptor signaling: Identification and mechanisms of action. Chem. Res. Toxicol. 2010, 24, 6–19. [Google Scholar] [CrossRef] [Green Version]
- Dong, X.; Dong, J.; Zhao, Y.; Guo, J.; Wang, Z.; Liu, M.; Zhang, Y.; Na, X. Effects of long-term in vivo exposure to di-2-ethyl-hexylphthalate on thyroid hormones and the TSH/TSHR signaling pathways in wistar rats. Int. J. Environ. Res. Public Health 2017, 14, 44. [Google Scholar] [CrossRef] [Green Version]
- Kim, M.; Jeong, J.S.; Kim, H.; Hwang, S.; Park, I.H.; Lee, B.C.; Yoon, S.I.; Jee, S.H.; Nam, K.T.; Lim, K.M. Low Dose Exposure to Di-2-Ethylhexyl Phthalate in Juvenile Rats Alters the Expression of Genes Related with Thyroid Hormone Regulation. Biomol. Ther. 2018, 26, 512–519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meeker, J.D.; Ferguson, K.K. Relationship between Urinary Phthalate and Bisphenol A Concentrations and Serum Thyroid Measures in U.S. Adults and Adolescents from the National Health and Nutrition Examination Survey (NHANES) 2007–2008. Environ. Health Perspect. 2011, 119, 1396–1402. [Google Scholar] [CrossRef] [PubMed]
- Baralić, K.; Buha, D.A.; Živančević, K.; Antonijević, E.; Anđelković, M.; Javorac, D.; Ćurčić, M.; Bulat, Z.; Antonijević, B.; Đukić, Ć.D. Toxic Effects of the Mixture of Phthalates and Bisphenol A-Subacute Oral Toxicity Study in Wistar Rats. Int. J. Environ. Res. Public Health 2020, 17, 746. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, D.; Zhou, L.; Wang, S.; Liu, T.; Zhu, J.; Jia, Y.; Xu, J.; Chen, H.; Wang, Q.; Xu, F.; et al. Effect of Di-(2-ethylhexyl) phthalate on the hypothalamus-pituitary-thyroid axis in adolescent rat. Endocr. J. 2018, 65, 261–268. [Google Scholar] [CrossRef] [Green Version]
- Fiore, M.; Oliveri, C.G.; Caltabiano, R.; Buffone, A.; Zuccarello, P.; Cormaci, L.; Cannizzaro, M.A.; Ferrante, M. Role of Emerging Environmental Risk Factors in Thyroid Cancer: A Brief Review. Int. J. Environ. Res. Public Health 2019, 16, 1185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miao, Y.; Wang, R.; Lu, C.; Zhao, J.; Deng, Q. Lifetime cancer risk assessment for inhalation exposure to di(2-ethylhexyl) phthalate (DEHP). Environ. Sci. Pollut. Res. Int. 2017, 24, 312–320. [Google Scholar] [CrossRef] [PubMed]
- Zhu, M.; Huang, C.; Ma, X.; Wu, R.; Zhu, W.; Li, X.; Liang, Z.; Deng, F.; Wu, J.; Geng, S.; et al. Phthalates promote prostate cancer cell proliferation through activation of ERK5 and p38. Environ. Toxicol. Pharmacol. 2018, 63, 29–33. [Google Scholar] [CrossRef] [PubMed]
- Buteau-Lozano, H.; Velasco, G.; Cristofari, M.; Balaguer, P.; Perrot, A.M. Xenoestrogens modulate vascular endothelial growth factor secretion in breast cancer cells through an estrogen receptor-dependent mechanism. J. Endocrinol. 2008, 196, 399–412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodgers, K.M.; Udesky, J.O.; Rudel, R.A.; Brody, J.G. Environmental chemicals and breast cancer: An updated review of epidemiological literature informed by biological mechanisms. Environ. Res. 2018, 160, 152–182. [Google Scholar] [CrossRef]
- Rusyn, I.; Corton, J.C. Mechanistic considerations for human relevance of cancer hazard of di (2-ethylhexyl) phthalate. Mutat. Res. 2012, 750, 141–158. [Google Scholar] [CrossRef] [Green Version]
- Chen, F.P.; Chien, M.H. Lower concentrations of phthalates induce proliferation in human breast cancer cells. Climacteric 2014, 17, 377–384. [Google Scholar] [CrossRef]
- Vacher, S.; Castagnet, P.; Chemlali, W.; Lallemand, F.; Meseure, D.; Pocard, M.; Bieche, I.; Perrot, A.M. High AHR expression in breast tumors correlates with expression of genes from several signaling pathways namely inflammation and endogenous tryptophan metabolism. PLoS ONE 2018, 13, e0190619. [Google Scholar] [CrossRef] [Green Version]
- Hsieh, T.H.; Tsai, C.F.; Hsu, C.Y.; Kuo, P.L.; Lee, J.N.; Chai, C.Y.; Wang, S.C.; Tsai, E.M. Phthalates induce proliferation and invasiveness of estrogen receptor-negative breast cancer through the AhR/HDAC6/c-Myc signaling pathway. FASEB J. 2012, 26, 778–787. [Google Scholar] [CrossRef] [Green Version]
- Ito, Y.; Yamanoshita, O.; Asaeda, N.; Tagawa, Y.; Lee, C.H.; Aoyama, T.; Ichihara, G.; Furuhashi, K.; Kamijima, M.; Gonzalez, F.J.; et al. Di (2-ethylhexyl) phthalate induces hepatic tumorigenesis through a peroxisome proliferator-activated receptor- independent Pathway. J. Occup. Health 2007, 49, 172–182. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Xin, H.; Fen, T. Function of MicroRNA-141 in Human Breast Cancer through Cytotoxic Cd4+ T Cells Regulated by MAP4K4 Expression. Mol. Med. Rep. 2018, 17, 7893–7901. [Google Scholar] [CrossRef] [PubMed]
- Zuccarello, P.; Olivieri, C.G.; Cavallaro, F.; Copat, C.; Cristaldi, A.; Fiore, M.; Ferrante, M. Implication of dietary phthalates in breast cancer. A systematic review. Food Chem. Toxicol. 2018, 118, 667–674. [Google Scholar] [CrossRef] [PubMed]
- Maradonna, F.; Carnevali, O. Lipid Metabolism Alteration by Endocrine Disruptors in Animal Models: An Overview. Front. Endocr. 2018, 9, 654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Samuel, V.T.; Shulman, G.I. Mechanisms for insulin resistance: Common threads and missing links. Cell 2012, 148, 852–871. [Google Scholar] [CrossRef] [Green Version]
- Lind, P.M.; Zethelius, B.; Lind, L. Circulating levels of phthalate metabolites are associated with prevalent diabetes in the elderly. Diabetes Care 2012, 35, 1519–1524. [Google Scholar] [CrossRef] [Green Version]
- Salgado, A.L.; Carvalho, L.; Oliveira, A.C.; Santos, V.N.; Vieira, J.G.; Parise, E.R. Insulin resistance index (HOMA-IR) in the differentiation of patients with nonalcoholic fatty liver disease and healthy individuals. Arq. Gastroenterol. 2010, 47, 165–169. [Google Scholar] [CrossRef] [Green Version]
- Dales, R.E.; Kauri, L.M.; Cakmak, S. The associations between phthalate exposure and insulin resistance, β-cell function and blood glucose control in a population-based sample. Sci. Total Environ. 2018, 612, 1287–1292. [Google Scholar] [CrossRef]
- Milošević, N.; Milanović, M.; Sudji, J.; Živanović, D.B.; Stojanoski, S.; Vuković, B.; Milić, N.; Stojanoska, M.M. Could phthalates exposure contribute to the development of metabolic syndrome and liver disease in humans? Environ. Sci. Pollution Res. 2020, 27, 772–784. [Google Scholar] [CrossRef]
- Harada, S.; Miyagi, K.; Obata, T.; Morimoto, Y.; Nakamoto, K.; Kim, K.I.; Kim, S.K.; Kim, S.R.; Tokuyama, S. Influence of hyperglycemia on liver inflammatory conditions in the early phase of non-alcoholic fatty liver disease in mice. J. Pharm. Pharmacol. 2017, 69, 698–705. [Google Scholar] [CrossRef]
- Unger, G.; Benozzi, S.F.; Perruzza, F.; Pennacchiotti, G.L. Triglycerides and glucose index: A useful indicator of insulin resistance. Endocrinol. Nutr. 2014, 61, 533–540. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Du, T.; Li, M.; Jia, J.; Lu, H.; Lin, X.; Yu, X. Triglyceride glucose-body mass index is effective in identifying nonalcoholic fatty liver disease in non-obese subjects. Medicine 2017, 96, e7041. [Google Scholar] [CrossRef] [PubMed]
- Lambrinoudaki, I.; Kazani, M.V.; Armeni, E.; Georgiopoulos, G.; Tampakis, K.; Rizos, D.; Augoulea, A.; Kaparos, G.; Alexandrou, A.; Stamatelopoulos, K. The TyG index as a marker of subclinical atherosclerosis and arterial stiffness in lean and overweight postmenopausal women. Heart Lung Circ. 2018, 27, 716–724. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Park, H.Y.; Bae, S.; Lim, Y.H.; Hong, Y.C. Diethylhexyl phthalates is associated with insulin resistance via oxidative stress in the elderly: A panel study. PLoS ONE 2013, 8, e71392. [Google Scholar] [CrossRef]
- Zhang, Y.; Meng, X.; Chen, L.; Li, D.; Zhao, L.; Zhao, Y.; Li, L.; Shi, H. Age and sex-specific relationships between phthalate exposures and obesity in Chinese children at puberty. PLoS ONE 2014, 9, e104852. [Google Scholar] [CrossRef]
- Buser, M.C.; Murray, H.E.; Scinicariello, F. Age and sex differences in childhood and adulthood obesity association with phthalates: Analyses of NHANES 2007–2010. Int. J. Hyg. Environ. Health 2014, 217, 687–694. [Google Scholar] [CrossRef]
- Wang, H.; Zhou, Y.; Tang, C.; He, Y.; Wu, J.; Chen, Y.; Jiang, Q. Urinary Phthalate Metabolites Are Associated with Body Mass Index and Waist Circumference in Chinese School Children. PLoS ONE 2013, 8, e56800. [Google Scholar] [CrossRef] [Green Version]
- Xia, B.; Zhu, Q.; Zhao, Y.; Ge, W.; Zhao, Y.; Song, Q.; Zhou, Y.; Shi, H.; Zhang, Y. Phthalate exposure and childhood overweight and obesity: Urinary metabolomic evidence. Environ. Int. 2018, 121, 159–168. [Google Scholar] [CrossRef]
- Aguilar-Morales, I.; Colin, R.E.; Rivera, M.S.; Vallejo, M.; Vázquez, A.C. Performance of waist-to-height ratio, waist circumference, and body mass index in discriminating cardio-metabolic risk factors in a sample of school-aged Mexican Children. Nutrients 2018, 10, 1850. [Google Scholar] [CrossRef] [Green Version]
- Medic, S.M.; Milosevic, N.; Milic, N.; Abenavoli, L. The influence of phthalates and bisphenol A on the obesity development and glucose metabolism disorders. Endocrine 2017, 55, 666–681. [Google Scholar]
- Gaston, S.A.; Tulve, N.S. Urinary phthalate metabolites and metabolic syndrome in U.S. adolescents: Cross-sectional results from the National Health and Nutrition Examination Survey (2003–2014) data. Int. J. Hyg. Environ. Health 2019, 222, 195–204. [Google Scholar] [CrossRef] [PubMed]
- Samuel, V.T.; Shulman, G.I. The pathogenesis of insulin resistance: Integrating signaling pathways and substrate flux. J. Clin. Invest. 2016, 126, 12–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Evans, R.M.; Barish, G.D.; Wang, Y.X. PPARs and the complex journey to obesity. Nat. Med. 2004, 10, 355–361. [Google Scholar] [CrossRef]
- Grun, F.; Blumberg, B. Minireview: The case for obesogens. Mol. Endocrinol. 2009, 23, 1127–1134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Desvergne, B.; Feige, J.N.; Casals, C.C. PPAR-mediated activity of phthalates: A link to the obesity epidemic? Mol. Cell Endocrinol. 2009, 304, 43–48. [Google Scholar] [CrossRef]
- Johns, L.E.; Ferguson, K.K.; Soldin, O.P.; Cantonwine, D.E.; Rivera, G.L.O.; Del, T.L.V.A.; Calafat, A.M.; Ye, X.; Alshawabkeh, A.N.; Cordero, J.F.; et al. Urinary phthalate metabolites in relation to maternal serum thyroid and sex hormone levels during pregnancy: A longitudinal analysis. Reprod. Biol. Endocrinol. 2015, 13, 4. [Google Scholar] [CrossRef] [Green Version]
- Radke, E.G.; Braun, J.M.; Nachman, R.M.; Cooper, G.S. Phthalate exposure and neurodevelopment: A systematic review and meta-analysis of human epidemiological evidence. Environ. Int. 2020, 137, 105408. [Google Scholar] [CrossRef]
- Li, N.; Papandonatos, G.D.; Calafat, A.M.; Yolton, K.; Lanphear, B.P.; Chen, A.; Braun, J.M. Identifying periods of susceptibility to the impact of phthalates on children’s cognitive abilities. Environ. Res. 2019, 172, 604–614. [Google Scholar] [CrossRef]
- Polanska, K.; Ligocka, D.; Sobala, W.; Hanke, W. Phthalate exposure and child development: The Polish Mother and Child Cohort Study. Early Hum. Dev. 2014, 90, 477–485. [Google Scholar] [CrossRef]
- Lin, H.; Yuan, K.; Li, L.; Liu, S.; Li, S.; Hu, G.; Lian, Q.Q.; Ge, R.S. In utero exposure to diethyl-hexyl phthalate affects rat brain development: A behavioral and genomic approach. Int. J. Environ. Res. Public Health 2015, 12, 13696–13710. [Google Scholar] [CrossRef]
- Xu, Y.; Agrawal, S.; Cook, T.J.; Knipp, G.T. Di-(2-ethylhexyl)-phthalate affects lipid profiling in fetal rat brain upon maternal exposure. Arch. Toxicol. 2007, 81, 57–62. [Google Scholar] [CrossRef] [PubMed]
- Dai, Y.; Yang, Y.; Xu, X.; Hu, Y. Effects of uterine and lactational exposure to di-(2-ethylhexyl) phthalate on spatial memory and NMDA receptor of hippocampus in mice. Horm. Behav. 2015, 71, 41–48. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.N.; Cho, S.C.; Kim, Y.; Shin, M.S.; Yoo, H.J.; Kim, J.W.; Yang, Y.H.; Kim, H.-W.; Bhang, S.Y.; Hong, Y.-C. Phthalates exposure and attention-deficit/hyperactivity disorder in school-age children. Biol. Psychiatry 2009, 66, 958–963. [Google Scholar] [CrossRef] [PubMed]
- Bennett, D.; Bellinger, D.C.; Birnbaum, L.S.; Bradman, A.; Chen, A.; Cory-Slechta, D.A.; Engel, S.M.; Fallin, M.D.; Halladay, A.; Hauser, R.; et al. Project TENDR: Targeting Environmental Neuro-Developmental Risks The TENDR Consensus Statement. Environ. Health Perspect 2016, 124, A118–A122. [Google Scholar] [CrossRef]
- Engel, S.M.; Villanger, G.D.; Nethery, R.C.; Thomsen, C.; Sakhi, A.K.; Drover, S.S.M.; Hoppin, J.A.; Zeiner, P.; Knudsen, G.P.; Reichborn-Kjennerud, T.; et al. Prenatal Phthalates, Maternal Thyroid Function, and Risk of Attention-Deficit Hyperactivity Disorder in the Norwegian Mother and Child Cohort. Environ. Health Perspect. 2018, 126, 057004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chopra, V.; Harley, K.; Lahiff, M.; Eskenazi, B. Association between phthalates and attention deficit disorder and learning disability in US children, 6-15 years. Environ. Res. 2014, 128, 64–69. [Google Scholar] [CrossRef] [Green Version]
- Gao, H.; Wu, W.; Xu, Y.; Jin, Z.; Bao, H.; Zhu, P.; Su, P.; Sheng, J.; Hao, J.; Tao, F. Effects of prenatal phthalate exposure on thyroid hormone concentrations beginning at the embryonic stage. Sci. Rep. 2017, 7, 13106. [Google Scholar] [CrossRef] [Green Version]
- Murray, E.; Pearson, R.; Fernandes, M.; Santos, I.S.; Barros, F.C.; Victora, C.G.; Stein, A.; Matijasevich, A. Are fetal growth impairment and preterm birth causally related to child attention problems and ADHD? Evidence from a comparison between high-income and middle-income cohorts. J. Epidemiol. Comm. Health 2016, 70, 704–709. [Google Scholar] [CrossRef] [Green Version]
- Smith, C.A.; Holahan, M.R. Reduced hippocampal dendritic spine density and BDNF expression following acute postnatal exposure to Di(2-Ethylhexyl) phthalate in male long evans rat. PLoS ONE 2014, 9, e109522. [Google Scholar] [CrossRef] [Green Version]
- Butala, J.H.; David, R.M.; Gans, G.; McKee, R.H.; Guo, T.L.; Peachee, V.L.; White, J.K.L. Phthalate treatment does not influence levels of IgE or Th2 cytokines in B6C3F1 mice. Toxicology 2004, 201, 77–85. [Google Scholar] [CrossRef]
- Larsen, S.T.; Nielsen, G.D. Structure-activity relationship of immunostimulatory effects of phthalates. BMC Immunol. 2008, 9, 61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, M.H.; Park, J.; Chung, S.W.; Kang, B.Y.; Kim, S.H.; Kim, T.S. Enhancement of interleukin 4 production in activated CD4+ T cells by di phthalate plasticizers via increased NF-AT binding activity. Int. Arch. Allergy Immunol. 2004, 134, 213–222. [Google Scholar] [CrossRef] [PubMed]
- Kimber, J.; Dearman, R.J. An assessment of the ability of phthalates to influence immune and allergic responses. Toxicology 2010, 127, 73–82. [Google Scholar] [CrossRef] [PubMed]
- Maruyama, T.; Shiba, T.; Iizuka, H.; Matsuda, T.; Kurohane, K.; Imai, Y. Effect of phthalate esters on dendritic cell subsets and interleukin-4 production in fluorescein isothiocyanate-induced contact hypersensitivity. Microbiol. Immun. 2007, 51, 321–326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Badr, M.Z.; Shnyra, A.; Zoubine, M.; Norkin, M.; Herndon, B.; Quinn, T.; Miranda, R.N.; Cunningham, M.L.; Molteni, A. Phthalate-induced liver protection against deleterious effects of the Th1-response: A potentially serious health hazard. PPAR Res. 2008, 2007, 1–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greene, A.D.; Sroga, J.M.; Thomas, M.A.; Burns, K.A. Phthalates may alter macrophage signaling to promote a permissive environment for the initiation of endometriosis. Fertil. Steril. 2017, 108, e326–e327. [Google Scholar] [CrossRef]
- Hansen, J.F.; Bendtzen, K.; Boas, M.; Frederiksen, H.; Nielsen, C.H.; Rasmussen, Å.K.; Feldt, R.U. Influence of phthalates on cytokine production in monocytes and macrophages: A systematic review of experimental trials. PLoS ONE 2015, 10, e0120083. [Google Scholar] [CrossRef] [Green Version]
- Piepenbrink, M.S.; Hussain, I.; Marsh, J.A.; Dietert, R.R. Developmental immunotoxicology of di-(2-ethylhexyl)phthalate (DEHP): Age-based assessment in the female rat. J. Immunotoxicol. 2005, 2, 21–31. [Google Scholar] [CrossRef] [Green Version]
- Yang, Q.; Xie, Y.; Depierre, J.W. Effects of peroxisome proliferators on the thymus and spleen of mice. Clin. Exp. Immunol. 2000, 122, 219–226. [Google Scholar] [CrossRef]
- Dogra, R.; Khanna, S.; Srivastava, S.; Shukla, L.; Chandra, K.; Saxena, G.; Shanker, R. Immunomodulation due to coexposure to styrene and dioctyl phthalate in mice. Immunopharmacol. Immunotoxicol. 1993, 15, 491–514. [Google Scholar] [CrossRef]
- Takano, H.; Yanagisawa, R.; Inoue, K.I.; Ichinose, T.; Sadakane, K.; Yoshikawa, T. Di-(2-ethylhexyl) phthalate enhances atopic dermatitis-like skin lesions in mice. Environ. Health Perspect. 2006, 114, 1266–1269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferguson, K.K.; Loch, C.R.; Meeker, J.D. Urinary phthalate metabolites in relation to biomarkers of inflammation and oxidative stress: NHANES 1999–2006. Environ. Res. 2011, 111, 718–726. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hurst, C.H.; Waxman, D.J. Activation of PPARalpha and PPARgamma by environmental phthalate monoesters. Toxicol. Sci. 2003, 74, 297–308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hong, C.C.; Shimomura, S.M.; Muroi, M.; Tanamoto, K.I. Effect on endocrine disrupting chemicals on lipopolysaccharide-induced tumor necrosis factor- and nitric oxide production by mouse macrophages. Biol. Pharm. Bull. 2004, 27, 1136–1139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohnishi, T.; Yoshida, T.; Igarashi, A.; Muroi, M.; Tanamoto, K.I. Effects of possi- ble endocrine disruptors on MyD88-independent TLR signalling. FEMS Immunol. Med. Microbiol. 2008, 52, 293–295. [Google Scholar] [CrossRef]
- Schlezinger, J.J.; Howard, G.J.; Hurst, C.H.; Emberley, J.K.; Waxman, D.J.; Webster, T.; Sherr, D.H. Environmental and endogenous peroxisome proliferator- activated receptor gamma agonists induce bone marrow B cell growth arrest and apoptosis: Interactions between mono(2-ethylhexyl)phthalate, 9-cis-retinoic acid, and 15-deoxy-Delta12,14-prostaglandin J2. J. Immunol. 2004, 173, 3165–3177. [Google Scholar] [CrossRef]
- Bissonnette, S.L.; Teague, J.E.; Sherr, D.H.; Schlezinger, J.J. An endogenous prostaglandin enhances environmental phthalate-induced apoptosis in bone marrow B cells: Activation of distinct but overlapping pathways. J. Immunol. 2008, 181, 1728–1736. [Google Scholar] [CrossRef] [Green Version]
- Larsen, S.T.; Hansen, J.S.; Thygesen, P.; Begtrup, M.; Poulsen, O.M.; Nielsen, G.D. Adjuvant and immuno-suppressive effect of six mono phthalates in a subcutaneous injection model with BALB/c mice. Toxicology 2001, 169, 37–51. [Google Scholar] [CrossRef]
- Jepsen, K.; Abildtrup, A.; Larsen, S. Monophthalates promote IL-6 and IL-8 production in the human epithelial cell line A549. Toxicol In Vitro. 2004, 18, 265–269. [Google Scholar] [CrossRef]
- Larsen, S.T.; Lund, R.M.; Nielsen, G.D.; Thygesen, P.; Poulsen, O.M. Adjuvant effect of di-n-butyl-, di-n-octyl-, di-iso-nonyl-and di-iso-decyl phthalate in a subcutaneous injection model using BALB/c mice. Pharmcl. Toxic. 2002, 91, 264–272. [Google Scholar] [CrossRef]
- Glue, C.; Platzer, M.H.; Larsen, S.T.; Nielsen, G.D.; Skov, P.S.; Poulsen, L.K. Phthalates potentiate the response of allergic effector cells. Basic Clin. Pharmacol. Toxicol. 2005, 96, 140–142. [Google Scholar] [CrossRef] [PubMed]
- Larsen, S.T.; Hansen, J.S.; Hansen, E.W.; Clausen, P.A.; Nielsen, G.D. Airway inflammation and adjuvant effect after repeated airborne exposures to di-(2-ethylhexyl) phthalate and ovalbumin in BALB/c mice. Toxicology 2007, 235, 119–129. [Google Scholar] [CrossRef] [PubMed]
- Hansen, J.S.; Larsen, S.T.; Poulsen, L.K.; Nielsen, G.D. Adjuvant effects of inhaled mono-2-ethylhexyl phthalate in BALB/cJ mice. Toxicology 2007, 232, 79–88. [Google Scholar] [CrossRef] [PubMed]
- Costantini, E.; D’Angelo, C.; Reale, M. The Role of Immunosenescence in Neurodegenerative Diseases. Mediat. Inflamm. 2018, 6039171. [Google Scholar] [CrossRef] [PubMed]
- Couleau, N.; Falla, J.; Beillerot, A.; Battaglia, E.; d’Innocenzo, M.; Plançon, S.; Laval-Gilly, P.; Bennasroune, A. Effects of endocrine disruptor compounds, alone or in combination, on human macrophage-like THP-1 cell response. PLoS ONE 2015, 10, e0131428. [Google Scholar] [CrossRef]
- Bennasroune, A.; Rojas, L.; Foucaud, L.; Goulaouic, S.; Laval-Gilly, P.; Fickova, M.; Couleau, N.; Durandet, C.; Henry, S.; Falla, J. Effects of 4-nonylphenol and/or diisononylphthalate on THP-1 cells: Impact of endocrine disruptors on human immune system parameters. Int. J. Immunopath. Pharmacol. 2012, 25, 365–376. [Google Scholar] [CrossRef]
- Miao, S.; Gao, Z.; Kou, Z.; Xu, G.; Su, C.; Liu, N. Influence of bisphenol on developing rat estrogen receptors and some cytokines in rats: A two-generational study. J. Toxic.Environ. Health Part A 2008, 71, 1000–1008. [Google Scholar] [CrossRef]
- Bansal, A.; Rashid, C.; Xin, F.; Li, C.; Polyak, E.; Duemler, A.; Van, D.M.T.; Stefaniak, M.; Wajid, S.; Doliba, N. Sex-and dose-specific effects of maternal bisphenol A exposure on pancreatic islets of first-and second-generation adult mice offspring. Environ. Health Perspect. 2017, 125, 097022. [Google Scholar] [CrossRef]
- Ghisletti, S.; Meda, C.; Maggi, A.; Vegeto, E. 17β-estradiol inhibits inflammatory gene expression by controlling NF-κB intracellular localization. Mol. Cell. Biol. 2005, 25, 2957–2968. [Google Scholar] [CrossRef] [Green Version]
- Dogra, R.; Khanna, S.; Nagale, S.; Shukla, L.; Srivastava, S.; Bhatnagar, M.; Gupta, P.; Shanker, R. Effect of dioctyl phthalate on immune system of rat. Indian J. Expt. Biol. 1985, 23, 315–319. [Google Scholar]
- Dogra, R.; Khanna, S.; Shukla, L.; Srivastava, S.; Gupta, S.; Katiyar, J.C.; Shanker, R. Modification of the immune response in rats by di-octyl phthalate. Indst. Health 1987, 25, 97–101. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Lei, P.; Liu, X.; Li, X.; Walker, K.; Kotha, L.; Rowlands, C.; Safe, S. The aryl hydrocarbon receptor as a target for estrogen receptor-negative breast cancer chemotherapy. Endocr. Relat. Cancer 2009, 16, 835–844. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, M.; Holloway, A.; Foster, W. Benzo-[a]-pyrene increases invasion in MDA-MB-231 breast cancer cells via increased COX-II expression and prostaglandin E 2 (PGE 2) output. Clin. Expt. Metastasis 2005, 22, 149–156. [Google Scholar] [CrossRef]
- Zhang, X.; Odom, D.T.; Koo, S.H.; Conkright, M.D.; Canettieri, G.; Best, J.; Chen, H.; Jenner, R.; Herbolsheimer, E.; Jacobsen, E. Genome-wide analysis of cAMP-response element binding protein occupancy, phosphorylation, and target gene activation in human tissues. Proc. Natl. Acad. Sci. USA 2005, 102, 4459–4464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, Y.S.; Lim, K.H.; Guo, X.; Kawaguchi, Y.; Gao, Y.; Barrientos, T.; Ordentlich, P.; Wang, X.F.; Counter, C.M.; Yao, T.P. The cytoplasmic deacetylase HDAC6 is required for efficient oncogenic tumorigenesis. Cancer Res. 2008, 68, 7561–7569. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martinez-Arguelles, D.B.; Guichard, T.; Culty, M.; Zirkin, B.R.; Papadopoulos, V. In utero exposure to the antiandrogen di-(2-ethylhexyl) phthalate decreases adrenal aldosterone production in the adult rat. Biol. Reprod. 2011, 85, 51–61. [Google Scholar] [CrossRef] [Green Version]
- Kuo, C.H.; Hsieh, C.C.; Kuo, H.F.; Huang, M.Y.; Yang, S.N.; Chen, L.C.; Huang, S.K.; Hung, C.H. Phthalates suppress type I interferon in human plasmacytoid dendritic cells via epigenetic regulation. Allergy 2013, 68, 870–879. [Google Scholar] [CrossRef]
- Fonteneau, J.F.; Gilliet, M.; Larsson, M.; Dasilva, I.; Münz, C.; Liu, Y.J.; Bhardwaj, N. Activation of influenza virus–specific CD4+ and CD8+ T cells: A new role for plasmacytoid dendritic cells in adaptive immunity. Blood 2003, 101, 3520–3526. [Google Scholar] [CrossRef]
- Arvanitoyannis, I.S.; Bosnea, L. Migration of substances from food packaging materials to foods. Critical Rev. Food Sci. Nutr. 2004, 44, 63–76. [Google Scholar] [CrossRef]
- Bhunia, K.; Sablani, S.S.; Tang, J.; Rasco, B. Migration of chemical compounds from packaging polymers during microwave, conventional heat treatment, and storage. Compr. Rev. Food Sci. Food Safety 2013, 12, 523–545. [Google Scholar] [CrossRef]
- Gallart-Ayala, H.; Núñez, O.; Lucci, P. Recent advances in LC-MS analysis of food-packaging contaminants. TrAC Trends Anal. Chem. 2013, 42, 99–124. [Google Scholar] [CrossRef] [Green Version]
- EFSA. Report on the development of a Food Classification and Description System for exposure assessment and guidance on its implementation and use. EFSA J. 2011, 9, 2489. [Google Scholar] [CrossRef]
- EFSA. Commission directive 2006/141/EC of 22 December 2006 on infant formulae and follow-on formulae and amending Directive 1999/21/EC. EFSA J. 2006, 4, 4544. [Google Scholar]
- Blank, R.; Burau, V.; Kuhlmann, E. Comparative Health Policy, 5th ed.; Macmillan International Higher Education: New York, NY, USA, 2017. [Google Scholar]
- Cao, X.L. Phthalate esters in foods: Sources, occurrence, and analytical methods. Compr. Rev. Food Sci. Food Saf. 2010, 9, 21–43. [Google Scholar] [CrossRef]
- Del Carlo, M.; Pepe, A.; Sacchetti, G.; Compagnone, D.; Mastrocola, D.; Cichelli, A. Determination of phthalate esters in wine using solid-phase extraction and gas chromatography–mass spectrometry. Food Chem. 2008, 111, 771–777. [Google Scholar] [CrossRef]
- Carrillo, J.D.; Martínez, M.P.; Tena, M.T. Determination of Phthalates in Wine by Headspace Solid-Phase Microextraction Followed by Gas Chromatography-Mass Spectrometry. Use of Deuterated Phthalates as Internal Standards. J. Chromatogr. A 2008, 1181, 125–130. [Google Scholar] [CrossRef]
- Russo, P.; Nastrucci, C.; Alzetta, G.; Szalai, C. Tobacco habit: Historical, cultural, neurobiological, and genetic features of people’s relationship with an addictive drug. Perspc. Biol. Med. 2011, 54, 557–577. [Google Scholar] [CrossRef]
- Hayasaka, Y. Analysis of phthalates in wine using liquid chromatography tandem mass spectrometry combined with a hold-back column: Chromatographic strategy to avoid the influence of pre-existing phthalate contamination in a liquid chromatography system. J. Chromatogr. A 2014, 1372, 120–127. [Google Scholar] [CrossRef]
- Chatonnet, P.; Boutou, S.; Plana, A. Contamination of wines and spirits by phthalates: Types of contaminants present, contamination sources and means of prevention. Food Addit. Contam. Part A 2014, 31, 1605–1615. [Google Scholar] [CrossRef]
- Jurica, K.; Brcic, K.I.; Lasić, D.; Vukic, L.D.; Anic, J.S.; Lusic, D. Determination of phthalates in plum spirit and their occurrence during plum spirit production. Acta Aliment. 2016, 45, 141–148. [Google Scholar] [CrossRef] [Green Version]
- Pellegrino Vidal, R.; Ibañez, G.; Escandar, G. A green method for the quantification of plastics-derived endocrine disruptors in beverages by chemometrics-assisted liquid chromatography with simultaneous diode array and fluorescent detection. Talanta 2016, 159. [Google Scholar] [CrossRef]
- Montevecchi, G.; Masino, F.; Zanasi, L.; Antonelli, A. Determination of phthalate esters in distillates by ultrasound-vortex-assisted dispersive liquid-liquid micro-extraction (USVADLLME) coupled with gas chromatography/mass spectrometry. Food Chem. 2017, 221, 1354–1360. [Google Scholar] [CrossRef]
- Plank, C.M.; Trela, B.C. A Review of Plastics Use in Winemaking: HACCP Considerations. Am. J. Enol. Vitic. 2018, 69, 307–320. [Google Scholar] [CrossRef]
- Grinbaum, M.; Camponovo, A.; Jean-Michel, D.; Poupault, P.; Meisterman, E.; Chatelet, B.; Davaux, F.; Lempereur, V. Phthalates: Potential sources and control measures. In Proceedings of the 41st World Congress of Vine and Wine, Punta del Este, Uruguay, 19–23 November 2018. [Google Scholar]
- Cinelli, M.; Coles, R.; Kirwan, K. Use of multi criteria decision analysis to support life cycle sustainability assessment: An analysis of the appropriateness of the available methods. In Proceedings of the 6th International Conference on Life Cycle Management, Gothenburg, Sweden, 25–28 August 2013. [Google Scholar]
- Guo, J.; Luo, K.; Chen, D.; Tan, X.; Song, Z. A rapid and sensitive method for the determination of dibutyl phthalate in wine by flow-injection chemiluminescence analysis. J. Food Compos. Anal. 2013, 31, 226–231. [Google Scholar] [CrossRef]
- Fan, Y.; Liu, S.; Xie, Q. Rapid determination of phthalate esters in alcoholic beverages by conventional ionic liquid dispersive liquid–liquid microextraction coupled with high performance liquid chromatography. Talanta 2014, 119, 291–298. [Google Scholar] [CrossRef] [PubMed]
- March, J.G.; Cerdà, V. An innovative arrangement for in-vial membrane-assisted liquid-liquid microextraction: Application to the determination of esters of phthalic acid in alcoholic beverages by gas chromatography-mass spectrometry. Anal. Bioanal. Chem. 2015, 407, 4213–4217. [Google Scholar] [CrossRef] [PubMed]
- Amiridou, D.; Voutsa, D. Alkylphenols and phthalates in bottled waters. J. Hazard. Mater. 2011, 185, 281–286. [Google Scholar] [CrossRef]
- Plotan, M.; Frizzell, C.; Robinson, V.; Elliott, C.T.; Connolly, L. Endocrine disruptor activity in bottled mineral and flavoured water. Food Chem. 2013, 136, 1590–1596. [Google Scholar] [CrossRef]
- Bach, C.; Dauchy, X.; Chagnon, M.C.; Etienne, S. Chemical compounds and toxicological assessments of drinking water stored in polyethylene terephthalate (PET) bottles: A source of controversy reviewed. Water Res. 2012, 46, 571–583. [Google Scholar]
- Baram, G.; Azarova, I.; Gorshkov, A.; Vereshchagin, A.; Lang, B.; Kiryukhina, E. Determination of bis (2-ethylhexyl) phthalate in water by high-performance liquid chromatography with direct on-column preconcentration. J. Anal. Chem. 2000, 55, 750–754. [Google Scholar] [CrossRef]
- Biscardi, D.; Monarca, S.; De Fusco, R.; Senatore, F.; Poli, P.; Buschini, A.; Rossi, C.; Zani, C. Evaluation of the migration of mutagens/carcinogens from PET bottles into mineral water by Tradescantia/micronuclei test, Comet assay on leukocytes and GC/MS. Sci. Total Environ. 2003, 302, 101–108. [Google Scholar] [CrossRef]
- Sugita, T.; Hirayama, K.; Nino, R.; Ishibashi, T.; Yamada, T. Contents of phthalate in polyvinyl chloride toys. Shokuhin Eiseigaku Zasshi. J. Food Hygienic Soc. Jpn. 2001, 42, 48–55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bono-Blay, F.; Guart, A.; De, L.F.B.; Pedemonte, M.; Pastor, M.C.; Borrell, A.; Lacorte, S. Survey of phthalates, alkylphenols, bisphenol A and herbicides in Spanish source waters intended for bottling. Environ. Sci. Pollution Res. 2012, 19, 3339–3349. [Google Scholar] [CrossRef] [PubMed]
- Keresztes, S.; Tatar, E.; Czegény, Z.; Zaray, G.; Mihucz, V.G. Study on the leaching of phthalates from polyethylene terephthalate bottles into mineral water. Sci. Total Envt. 2013, 458, 451–458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Saleh, I.; Shinwari, N.; Alsabbaheen, A. Phthalates residues in plastic bottled waters. J. Toxicol. Sci. 2011, 36, 469–478. [Google Scholar] [CrossRef] [Green Version]
- Montuori, P.; Jover, E.; Morgantini, M.; Bayona, J.M.; Triassi, M. Assessing human exposure to phthalic acid and phthalate esters from mineral water stored in polyethylene terephthalate and glass bottles. Food Addit. Contam. 2008, 25, 511–518. [Google Scholar] [CrossRef] [Green Version]
- Casajuana, N.; Lacorte, S. Presence and release of phthalic esters and other endocrine disrupting compounds in drinking water. Chromatographia 2003, 57, 649–655. [Google Scholar] [CrossRef]
- Schmid, P.; Kohler, M.; Meierhofer, R.; Luzi, S.; Wegelin, M. Does the reuse of PET bottles during solar water disinfection pose a health risk due to the migration of plasticisers and other chemicals into the water? Water Res. 2008, 42, 5054–5060. [Google Scholar] [CrossRef]
- Luo, Q.; Liu, Z.H.; Yin, H.; Dang, Z.; Wu, P.X.; Zhu, N.W.; Lin, Z.; Liu, Y. Migration and potential risk of trace phthalates in bottled water: A global situation. Water Res. 2018, 147, 362–372. [Google Scholar] [CrossRef]
- Abtahi, M.; Dobaradaran, S.; Torabbeigi, M.; Jorfi, S.; Gholamnia, R.; Koolivand, A.; Darabi, H.; Kavousi, A.; iSaeedi, R. Health risk of phthalates in water environment: Occurrence in water resources, bottled water, and tap water, and burden of disease from exposure through drinking water in Tehran, Iran. Environ. Res. 2019, 173, 469–479. [Google Scholar] [CrossRef]
- Bošnir, J.; Puntarić, D.; Galić, A.; Škes, I.; Dijanić, T.; Klarić, M.; Grgić, M.; Čurković, M.; Šmit, Z. Migration of phthalates from plastic containers into soft drinks and mineral water. Food Technol. Biotechnol. 2007, 45, 91–95. [Google Scholar]
- Ustun, I.; Sungur, S.; Okur, R.; Sumbul, A.; Oktar, S.; Yilmaz, N.; Gokce, C. Determination of Phthalates Migrating from Plastic Containers into Beverages. Food Anal. Methods 2015, 8, 222–228. [Google Scholar] [CrossRef]
- Wu, C.F.; Chang-Chien, G.P.; Su, S.W.; Chen, B.H.; Wu, M.T. Findings of 2731 suspected phthalate-tainted foodstuffs during the 2011 phthalates incident in Taiwan. J. Formos. Med. Assoc. 2014, 113, 600–605. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Troisi, J.; Richards, S.; Symes, S.; Ferretti, V.; Di Maio, A.; Amoresano, A.; Daniele, B.; Aliberti, F.; Guida, M.; Trifuoggi, M. A comparative assessment of metals and phthalates in commercial tea infusions: A starting point to evaluate their tolerance limits. Food Chem. 2019, 288, 193–200. [Google Scholar] [CrossRef] [PubMed]
- Fankhauser-Noti, A.; Grob, K. Migration of plasticizers from PVC gaskets of lids for glass jars into oily foods: Amount of gasket material in food contact, proportion of plasticizer migrating into food and compliance testing by simulation. Trends Food Sci. Technol. 2006, 17, 105–112. [Google Scholar] [CrossRef]
- Nanni, N.; Fiselier, K.; Grob, K.; Di Pasquale, M.; Fabrizi, L.; Aureli, P.; Coni, E. Contamination of vegetable oils marketed in Italy by phthalic acid esters. Food Control. 2011, 22, 209–214. [Google Scholar] [CrossRef]
- Long-Kai, S.; Ming-Ming, Z.; Yu-Lan, L. Concentration and survey of phthalic acid esters in edible vegetable oils and oilseeds by gas chromatography-mass spectrometry in China. Food Control. 2016, 68, 118–123. [Google Scholar]
- Marega, M.; Grob, K.; Moret, S.; Purcaro, G.; Conte, L.S. Phtalate contamination in olive oil production chain. In Proceedings of the Workshop Contaminants of Edible Fats and Oils: Analytical, normative issues and prevention, Udine, Italy, 12–13 November 2009. [Google Scholar]
- Bi, X.; Pan, X.; Yuan, S.; Wang, Q. Plasticizer contamination in edible vegetable oil in a U.S. retail market. J. Agric. Food Chem. 2013, 61, 9502–9509. [Google Scholar] [CrossRef]
- Oh, M.S.; Lee, S.W.; Moon, M.; Lee, D.; Park, H.M. Simultaneous analysis of phthalates, adipate and polycyclic aromatic hydrocarbons in edible oils using isotope dilution-gas chromatography-mass spectrometry. Food Addit. Contam. Part B Surveill. 2014, 7, 168–175. [Google Scholar] [CrossRef]
- Lacoste, F. Undesirable substances in vegetable oils: Anything to declare? OCL 2014, 21, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Sungur, S.; Okur, R.; Turgut, F.; Ustun, I.; Gokce, C. Migrated phthalate levels into edible oils. Food Addit. Contam. Part B Surveill. 2015, 8, 190–194. [Google Scholar] [CrossRef]
- Xiang, W.; Gong, Q.; Xu, J.; Li, K.; Yu, F.; Chen, T.; Qin, S.; Li, C.; Wang, F. Cumulative risk assessment of phthalates in edible vegetable oil consumed by Chinese residents. J. Sci. Food Agric. 2020, 100, 1124–1131. [Google Scholar] [CrossRef] [PubMed]
- Luo, Q.; Liu, Z.H.; Yin, H.; Dang, Z.; Wu, P.X.; Zhu, N.W.; Lin, Z.; Liu, Y. Global review of phthalates in edible oil: An emerging and nonnegligible exposure source to human. Sci. Total Environ. 2020, 704, 135369. [Google Scholar] [CrossRef] [PubMed]
- Petersen, J.H. Survey of di-(2-ethylhexyl)phthalate plasticizer contamination of retail Danish milks. Food Addit. Contam. 1991, 8, 701–705. [Google Scholar] [CrossRef] [PubMed]
- Fierens, T.; Van Holderbeke, M.; Willems, H.; De Henauw, S.; Sioen, I. Phthalates in Belgian cow’s milk and the role of feed and other contamination pathways at farm level. Food. Chem. Toxicol. 2012, 50, 2945–2953. [Google Scholar] [CrossRef]
- Fierens, T.; Van Holderbeke, M.; Willems, H.; De Henauw, S.; Sioen, I. Transfer of eight phthalates through the milk chain-a case study. Environ. Int. 2013, 51, 1–7. [Google Scholar] [CrossRef]
- Sharman, M.; Read, W.A.; Castle, L.; Gilbert, J. Levels of di-(2-ethylhexyl) phthalate and total phthalate esters in milk, cream, butter and cheese. Food Addit. Contam. 1994, 11, 375–385. [Google Scholar] [CrossRef]
- MeeKyung, K.; Seon, J.Y.; Gab-Soo, C. Determination of phthalates in raw bovine milk by gas chromatography/time-of-flight mass spectrometry (GC/TOF-MS) and dietary intakes. Food Addit. Contam. Part A Chem. Anal. Control. Expo. Risk Assess. 2009, 26, 134–138. [Google Scholar]
- Jarošová, A.; Bogdanovičová, S. Phthalates in meat products in dependence on the fat content. Potravin. Slovak. J. Food Sci. 2016, 10, 378–383. [Google Scholar]
- Tsai, M.Y.; Ho, C.H.; Chang, H.Y.; Yang, W.; Lin, C.F.; Lin, C.T.; Xue, Y.J.; Lai, J.; Wang, J.; Chang, G.R. Analysis of Pollution of Phthalates in Pork and Chicken in Taiwan Using Liquid Chromatography–Tandem Mass Spectrometry and Assessment of Health Risk. Molecules 2019, 24, 3817. [Google Scholar] [CrossRef] [Green Version]
- Lü, H.; Mo, C.H.; Zhao, H.M.; Xiang, L.; Katsoyiannis, A.; Li, Y.W.; Cai, Q.Y.; Wong, M. Soil contamination and sources of phthalates and its health risk in China: A review. Environ. Res. 2018, 164, 417–429. [Google Scholar] [CrossRef]
- Sun, J.; Wu, X.; Gan, J. Uptake and Metabolism of Phthalate Esters by Edible Plants. Environ. Sci. Tech. 2015, 49, 8471–8478. [Google Scholar] [CrossRef]
- Chen, N.; Shuai, W.; Hao, X.; Zhang, H.; Zhou, D.; Gao, J. Contamination of Phthalate Esters in Vegetable Agriculture and Human Cumulative Risk Assessment. Pedosphere 2017, 27, 439–451. [Google Scholar] [CrossRef]
- Benjamin, S.; Masai, E.; Kamimura, N.; Takahashi, K.; Anderson, R.C.; Faisal, P.A. Phthalates impact human health: Epidemiological evidences and plausible mechanism of action. J. Hazard. Mater. 2017, 340, 360–383. [Google Scholar] [CrossRef]
- Pan, Y.; Jing, J.; Dong, F.; Yao, Q.; Zhang, W.; Zhang, H.; Yao, B.; Dai, J. Association between phthalate metabolites and bio- markers of reproductive function in 1066 Chinese men of reproductive age. J. Hazard. Mater. 2015, 300, 729–736. [Google Scholar] [CrossRef]
- Bloom, M.S.; Whitcomb, B.W.; Chen, Z.; Ye, A.; Kannan, K.; Buck Louis, G.M. Associations between urinary phthalate concentrations and semen quality parameters in a general population. Hum. Reprod 2015, 30, 2645–2657. [Google Scholar] [CrossRef]
- Thurston, S.W.; Mendiola, J.; Bellamy, A.R.; Levine, H.; Wang, C.; Sparks, A.; Redmon, J.B.; Drobnis, E.Z.; Swan, S.H. Phthalate exposure and semen quality in fertile US men. Andrology 2016, 4, 632–638. [Google Scholar] [CrossRef] [Green Version]
- Lenters, V.; Portengen, L.; Smit, L.A.; Jönsson, B.A.G.; Giwercman, A.; Rylander, L.; Lindh, C.; Spanò, M.; Pedersen, H.S.; Ludwicki, J.K.; et al. Phthalates, perfluoroalkyl acids, metals and organochlorines and reproductive function: A multipollutant assessment in Greenlandic, Polish and Ukrainian men. Occup. Environ. Med. 2015, 72, 385–393. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.; Wang, H.; Tian, M.; Zhang, J.; Panuwet, P.; D’Souza, P.E.; Barr, D.B.; Huang, Q.; Xia, Y.; Shen, H. Phthalate metabolites related to infertile biomarkers and infertility in Chinese men. Environ. Pollut. 2017, 231, 291–300. [Google Scholar] [CrossRef]
- Tian, M.; Liu, L.; Wang, H.; Wang, X.; Martin, F.L.; Zhang, J.; Huang, Q.; Shen, H. Phthalates induce androgenic effects at exposure levels that can be environmentally relevant in humans. Environ. Sci. Technol. Lett. 2018, 5, 232–236. [Google Scholar] [CrossRef]
- Høyer, B.B.; Lenters, V.; Giwercman, A.; Jönsson, B.A.G.; Toft, G.; Hougaard, K.S.; Bonde, J.P.E.; Specht, I.O. Impact of di-2-ethylhexyl phthalate metabolites on male reproductive function: A systematic review of human evidence. Curr. Environ. Health Rep. 2018, 5, 20–33. [Google Scholar] [CrossRef] [PubMed]
- Tsatsakis, A.; Docea, A.O.; Tsitsimpikou, C. New challenges in risk assessment of chemicals when simulating real exposure scenarios; Simultaneous multi-chemicals’ low dose exposure. Food Chem. Toxicol. 2016, 96. [Google Scholar] [CrossRef] [PubMed]
- Margina, D.; Nițulescu, G.M.; Ungurianu, A.; Mesnage, R.; Goumenou, M.; Sarigiannis, D.A.; Tsatsakis, A. Overview of the effects of chemical mixtures with endocrine disrupting activity in the context of real-life risk simulation (RLRS): An integrative approach. World Acad. Sci. J. 2019, 1, 157–164. [Google Scholar] [CrossRef] [PubMed]

| Chemical Name | Structure | Use |
|---|---|---|
| Diethyl phthalate (DEP) |  | Personal care products (e.g., fragrances), coatings (e.g., pharmaceuticals), dyes, pesticides |
| Di-(2-ethylhexyl) phthalate (DEHP) |  | Perfumes, PVC plastics used in household products (e.g., toys, floor tiles and furniture upholstery, cables, garden hoses, wall coverings, and gloves), food packaging, blood storage bags, and medical devices |
| Di-isononyl phthalate (DINP) |  | Plasticizer, remaining in rubbers, inks, adhesives and sealants, paints, and lacquers |
| Benzyl Butylphthalate (BBP) |  | Vinyl flooring, adhesives and sealants, car-care products, toys, food packaging, synthetic leather, industrial solvents, glues, personal care products, and automotive products |
| Di-n-butyl phthalate (DBP) |  | Cellulose acetate plastics, solvent for oil-soluble dyes, pesticides, personal care products (e.g., nail polish and cosmetics), lacquers, varnishes, and coatings (e.g., pharmaceuticals) |
| Phthalate | Abbreviation | Formula |
|---|---|---|
| Benzyl-butyl Phthalate | BBP | C19H20O4 |
| Di-butyl Phthalate | DBP | C16H22O4 |
| Di-cyclohexyl Phthalate | DCHP | C20H26O4 |
| Di-ethyl Phthalate | DEP | C12H14O4 |
| Di-etylhexyl Phthalate | DEHP | C24H38O4 |
| Di-isobutyl Phthalate | DIBP | C16H22O4 |
| Di-isodecyl Phthalate | DIDP | C28H46O4 |
| Di-isononyl Phthalate | DINP | C26H42O4 |
| Di-methyl Phthalate | DMP | C10H10O4 |
| Di-n-octyl Phthalate | DnOP | C24H38O4 |
| Mono-n-butyl phthalate | MnBP | C12H14O4 |
| Monobenzyl phthalate | MBzP | C15H12O4 |
| Monocarboxy-isononly phthalate | MCNP | C18H24O6 |
| Monocarboxyoctyl phthalate | MCOP | C₁₇H₂₂O₆ |
| Mono-(3-carboxypropyl) phthalate | MCPP | C12H12O6 |
| Mono(2-ethyl-5-carboxypentyl) phthalate | MECPP | C16H20O6 |
| Mono(2-ethyl-5-hydroxyhexyl) Phthalate | MEHHP | C16H22O5 |
| Mono(2-ethylhexyl) Phthalate | MEHP | C16H22O4 |
| Mono(2-ethyl-5-oxohexyl) Phthalate | MEOHP | C16H20O5 |
| Mono-ethyl phthalate | MEP | C10H10O4 |
| Monoisobutyl Phthalate | MiBP | C12H14O4 |
| Monoisononyl Phthalate | MINP | C17H24O4 |
| Mono-methyl Phthalate | MMP | C9H8O4 |
| Mono-methyl Phthalate | MNOP | C9H8O4 |
| Matrix | Phthalate | Average Concentration | Reference |
|---|---|---|---|
| Wines in glass bottle | DIBP | 0.099 | [189] |
| DBP | 0.053 | ||
| DEPH | 0.076 | ||
| BBP | 0.040 | ||
| Wines in polyethylene coupled film brick | DIBP | 0.076 | |
| DBP | 0.115 | ||
| DEHP | 0.078 | ||
| Wine in glass bottles with one-piece cork | DEP | 4.22 | [190] |
| DBP | 2.21 | ||
| BBP | 4.29 | ||
| Wine in glass bottles with synthetic stoppers | DEP | 2.95 | |
| DBP | 1.02 | ||
| DEHP | 5.22 | ||
| Wines in cartons | DBP | 2.22 | |
| DEHP | 3.90 | ||
| Wines in bag-in-box | DMP | 0.61 | |
| DEP | 1.78 | ||
| DBP | 0.30 | ||
| Commercial white wines in tetrapak box | DBP | 10.0 | [191] |
| BBP | 1.0 | ||
| DEHP | 16.0 | ||
| Commercial red wines in glass bottles | DBP | 7.3–23 | |
| BBP | 0.1–5.2 | ||
| DEHP | 3.1–15.8 | ||
| Commercial white wines in glass bottles | DBP | 19.3–21.3 | |
| BBP | 0.4–7.0 | ||
| DEHP | 9.2–15.1 | ||
| Home-prepared red wines | DBP | 22.8 | |
| BBP | ND | ||
| DEHP | 2.4 | ||
| Commercial red wine samples (Australia) | DIBP | 5.3–10.7 | [192] |
| DBP | 3.4–9.3 | ||
| DEHP | 1.7–4 | ||
| BBP | 3.5–6.3 | ||
| Commercial white wine samples (Australia) | DIBP | 4.6–9.1 | |
| DBP | 2.9–3.7 | ||
| DEHP | 2.3–4 | ||
| BBP | 0.3–1.1 | ||
| Commercial wines (France) | DBP | 0.273 | [193] |
| BBP | 0.008 | ||
| DEHP | 0.134 | ||
| Commercial spirits (France) | DBP | 0.314 | |
| BBP | 0.026 | ||
| DEHP | 0.513 | ||
| DIBP | 0.103 | ||
| Glass bottled plum spirits | DBP | 414.5 | [194] |
| DEHP | 423.8 | ||
| BBP | 79.0 | ||
| DIBP | 38.8 | ||
| Red wine samples | DBP | 334.0 | [195] |
| DEHP | 80.3 | ||
| DEP | 56 | ||
| Stout beer | DBP | 74.7 | |
| DEHP | 16.6 | ||
| DEP | 4.7 | ||
| Lager beer | DBP | 1.1 | |
| DEHP | 18.2 | ||
| DEP | ND | ||
| Schnapps | DBP | 76.6 | |
| DEHP | 28.0 | ||
| DEP | 4.7 | ||
| Cachaca | DBP | 40.5 | |
| DEHP | 140.0 | ||
| DEP | 25.8 | ||
| Base wine samples | DBP | 12.0–79.0 | [196] |
| DEHP | 5.0–41.0 | ||
| Brandy samples | DBP | 620.0 | |
| DEHP | 470.0 |
| Oil sources | Phthalate | Average Concentration | Reference |
|---|---|---|---|
| Extra virgin olive | DEHP | 1134 | [222] |
| DINP | 1722 | ||
| DBP | 90 | ||
| Olive | DEHP | 1262 | |
| DINP | 2884 | ||
| DBP | 360 | ||
| Sunflower | DEHP | 134 | |
| DINP | 971 | ||
| DBP | 35 | ||
| Various seed | DEHP | 132 | |
| DINP | 1361 | ||
| DBP | 30 | ||
| Corn | DEHP | 81 | |
| DINP | 2982 | ||
| DBP | 23 | ||
| Peanut | DEHP | 334 | |
| DINP | 1518 | ||
| DBP | 41 | ||
| Soybean | DEHP | 77 | |
| DINP | 1017 | ||
| DBP | 22 | ||
| Olive pomace | DEHP | 1643 | |
| DINP | 6480 | ||
| DBP | 224 | ||
| Peanut oil | DEHP | 1250 | [223] |
| DBP | 250 | ||
| Teaseed oil | DEHP | 1250 | |
| DBP | 1610 | ||
| Rice bran oil | DEHP | 650 | |
| DBP | 1060 | ||
| Sunflower oil | DEHP | 260 | |
| DBP | 140 | ||
| Soybean oil | DEHP | 140 | |
| DBP | 60 | ||
| Corn oil | DEHP | 100 | |
| DBP | 20 | ||
| Rapeseed oil | DEHP | 160 | |
| DBP | 470 | ||
| Cottonseed oil | DEHP | 350 | |
| DBP | 270 | ||
| Olive oil | DEHP | 850 | |
| DBP | 110 | ||
| Wheat germ oil | DEHP | 1110 | |
| DBP | 21,290 | ||
| Grape seed oil | DEHP | 930 | |
| DBP | 1690 | ||
| Walnut oil | DEHP | 1590 | |
| DBP | 1206 | ||
| Sesame | DEHP | 290 | [223] |
| DBP | 80 | ||
| Corn | DEHP | 150 | |
| DBP | 170 | ||
| Rapeseed | DEHP | 400 | |
| DBP | 90 | ||
| Teaseed | DEHP | 140 | |
| DBP | 20 | ||
| Soybean | DEHP | 190 | |
| DBP | 120 |
| Matrix | Phthalate | Average Concentration | Reference |
|---|---|---|---|
| Milk | DEHP | 20–480 | [231] Milk samples from Scandinavian countries |
| Milk in silo and tanker | DEHP | 60–140 | |
| Cream 35% fat | DEHP | 1060–1670 | |
| Milk <1% fat | DEHP | 20–40 | |
| Milk | DEHP | 10–40 | Milk samples from Spain |
| Cream | DEHP | 480–550 | |
| Milk | DEHP | 10–90 | Milk from UK |
| Cheese | DEHP | 600–3000 | |
| Cream | DEHP | 200–2700 | |
| Summer milk | DEHP | ND–787.6 | [232] |
| DIBP | ND–15 | ||
| DBP | ND–15.3 | ||
| BBP | ND–15.5 | ||
| Winter milk | DEHP | 201.3–499.7 | |
| DIBP | 17.2–51.5 | ||
| DBP | ND–15 | ||
| BBP | 10–20.5 | ||
| Milked by hand | DEHP | <60 | |
| DIBP | 29 | ||
| DBP | <15 | ||
| BBP | <10 | ||
| Milked by machine | DEHP | 123.5 | |
| DIBP | 15.1 | ||
| DBP | ND | ||
| BBP | 14.3 | ||
| Raw milk cooling tank | DEHP | 364 | [233] |
| DIBP | <15 | ||
| DBP | ND | ||
| BBP | ND | ||
| Pasteurized milk cooling tank | DEHP | 426 | |
| DIBP | ND | ||
| DBP | ND | ||
| BBP | <15 | ||
| Milk powder before filling | DEHP | 478 | |
| DIBP | 32 | ||
| DBP | 28 | ||
| BBP | ND | ||
| Milk after filling (can) | DEHP | 630 | |
| DIBP | 56 | ||
| DBP | 52 | ||
| BBP | 12 | ||
| Milk after filling (pouch) | DEHP | 523 | |
| DIBP | 31 | ||
| DBP | 60 | ||
| BBP | 53 | ||
| Milk powder at retail (can) | DEHP | 566 | |
| DBP | 75 | ||
| DBP | 53 | ||
| BBP | 12 | ||
| Milk powder at retail (pouch) | DEHP | 526 | |
| DIBP | 75 | ||
| DBP | 53 | ||
| BBP | 12 | ||
| DBP | 80 | ||
| DBP | 170 | ||
| Milk samples | DEHP | 57 | [234] |
| DBP | 30 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Giuliani, A.; Zuccarini, M.; Cichelli, A.; Khan, H.; Reale, M. Critical Review on the Presence of Phthalates in Food and Evidence of Their Biological Impact. Int. J. Environ. Res. Public Health 2020, 17, 5655. https://doi.org/10.3390/ijerph17165655
Giuliani A, Zuccarini M, Cichelli A, Khan H, Reale M. Critical Review on the Presence of Phthalates in Food and Evidence of Their Biological Impact. International Journal of Environmental Research and Public Health. 2020; 17(16):5655. https://doi.org/10.3390/ijerph17165655
Chicago/Turabian StyleGiuliani, Angela, Mariachiara Zuccarini, Angelo Cichelli, Haroon Khan, and Marcella Reale. 2020. "Critical Review on the Presence of Phthalates in Food and Evidence of Their Biological Impact" International Journal of Environmental Research and Public Health 17, no. 16: 5655. https://doi.org/10.3390/ijerph17165655
APA StyleGiuliani, A., Zuccarini, M., Cichelli, A., Khan, H., & Reale, M. (2020). Critical Review on the Presence of Phthalates in Food and Evidence of Their Biological Impact. International Journal of Environmental Research and Public Health, 17(16), 5655. https://doi.org/10.3390/ijerph17165655