3.2. Effect of Chromium Concentration on Seed Germination and Growth
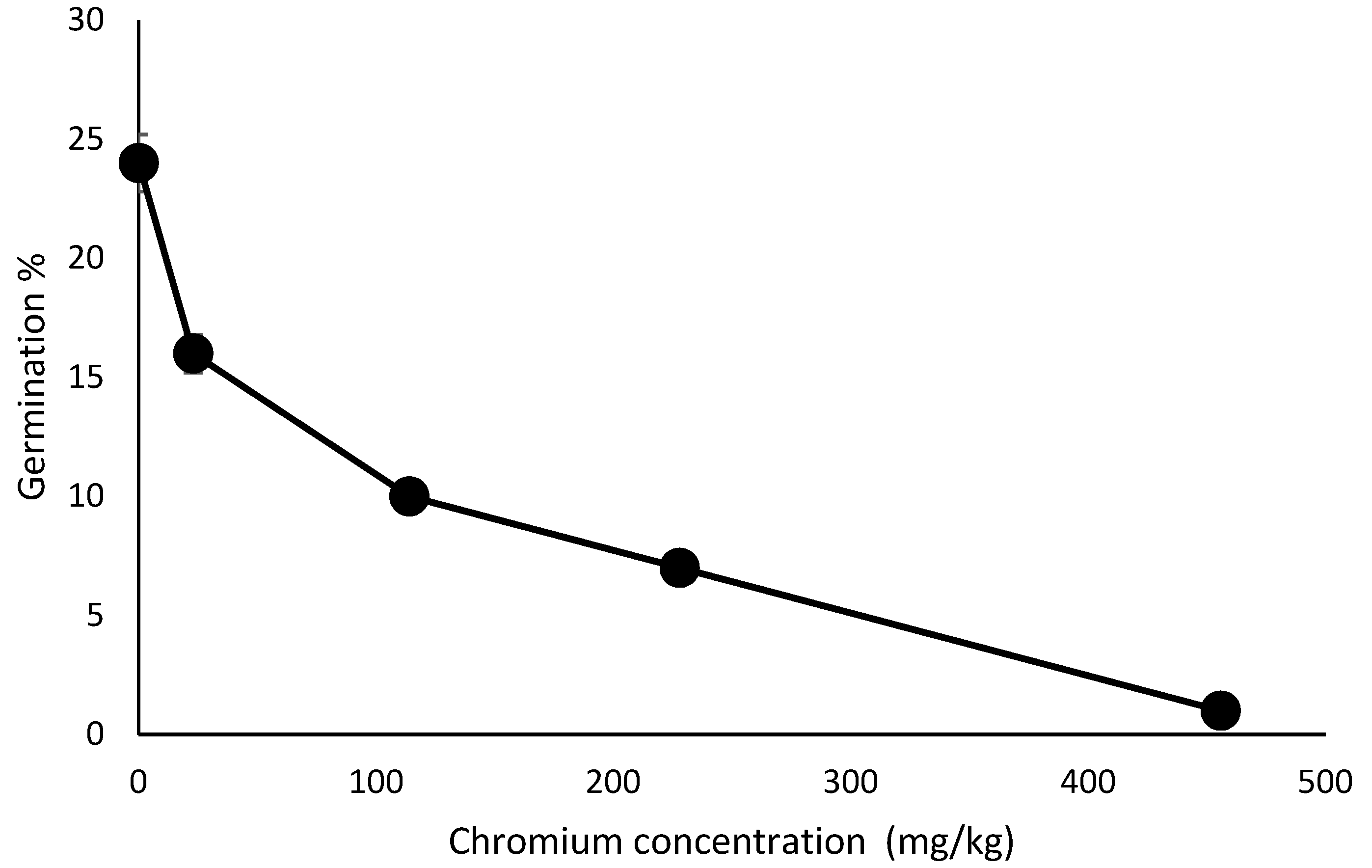
The observed results of the present study show that higher chromium concentration adversely influences the germination process of various vegetable seeds (
Figure 1). Chromium treatment in soils at the level of 23 to 456 mg/kg had different effects. It was observed that 24% germination took place in the control soils but decreased to 16% in soils with concentrations of 23 mg/kg, 10% germination corresponded to soils with 114 mg/kg of chromium concentration, 7% germinated in 228 mg/kg Cr levels and 1% managed to germinate in 456 mg/kg Cr levels. Significant variations in Cr tolerance and sensitivity in terms of seed germination have been recorded in literature, according to Shahid et al. [
24]. This study has established that the germination of different plant seeds is affected differently with different Cr levels (
Figure 1).
The germination time in this study was prolonged to (8 ± 1 days), while the control germination was observed after (6 ± 1 days). The prolonged germination period was observed as the levels of chromium increased from 23 mg/kg to 456 mg/kg. This suggests that the seeds may have undergone secondary induced dormancy in Cr conditions before germination, and this lengthened their germination period. Eze et al. [
28] stated that at a treatment level of 400 mg/kg of chromium in soil, a prolonged germination time (9 ± 1 days) was observed unlike the control (4 ± 1 days) and this implied that germination time increased with increase in chromium dosage.
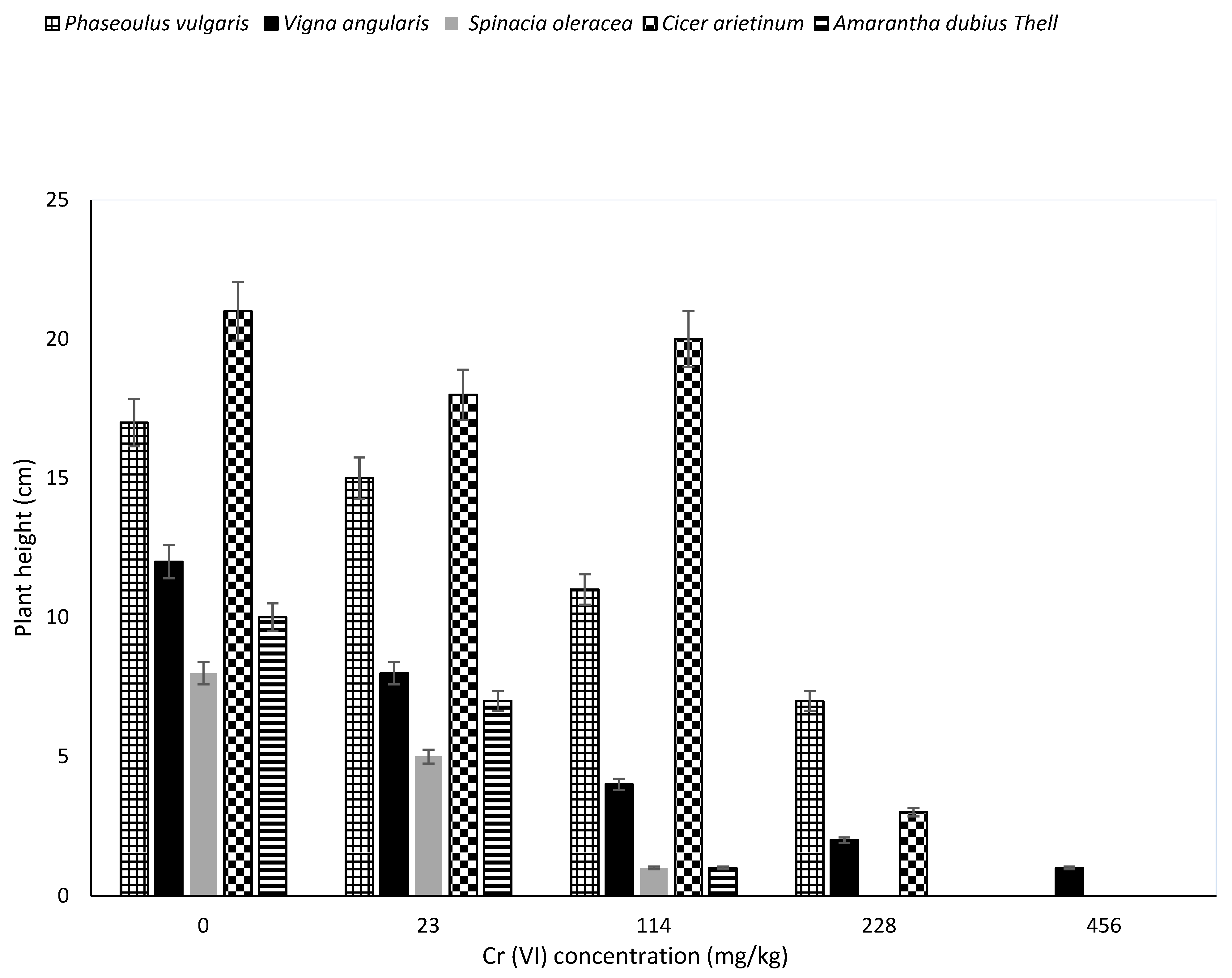
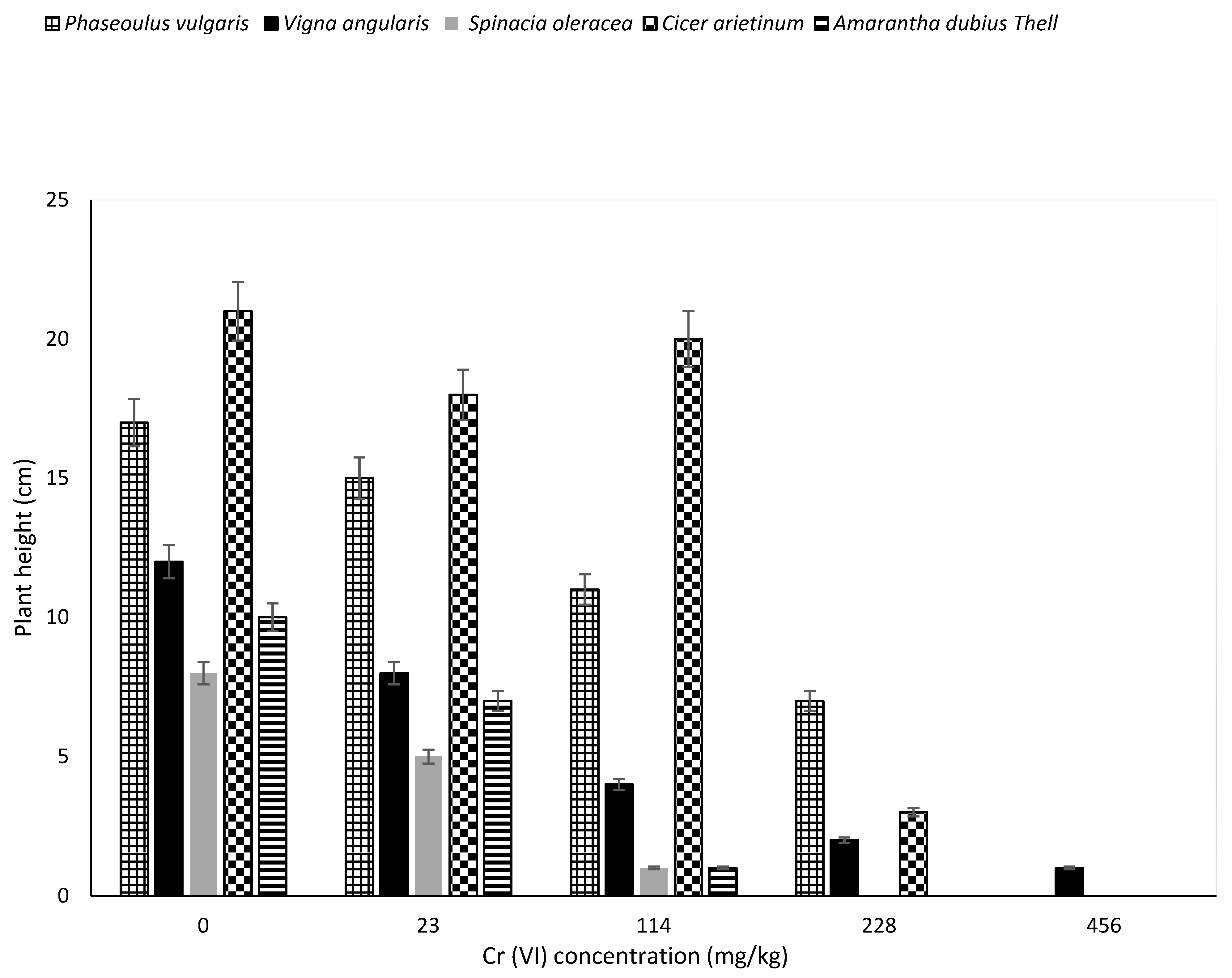
The Cr transported to the aerial parts or retained at the roots might have affected the physiological processes of plant growth as it contributed to their various reductions in height, as shown in
Figure 2. Significant differences (
p < 0.05) was found in plants height with increased concentration of chromium from 0 mg/kg (control) to 456 mg/kg. There was a significant difference between the height of plants in the control and those grown in soils containing varying levels of Cr (23–456 kg).
There was also a significant difference observed between Cicer arietinum and Spinacia oleracea, Phaseoulus vulgaris and Spinacia oleracea, Cicer arietinum and Amarantha dubius Thell, Phaseoulus vulgaris and Amarantha dubius Thell. The significant decrease in height could be attributed to Cr(VI) concentrations. In the 114 mg/kg Cr levels, the highest height recorded was that of Cicer arietinum, while the lowest were Spinacia oleracea and Amarantha dubius Thell. This possibly implies that Spinacia oleracea and Amarantha dubius Thell seeds growth are sensitive to a high increase in Cr(VI) concentrations at 114 mg/kg in comparison to other plants.
In 228 mg/kg levels, the maximum height recorded was that of
Phaseoulus vulgaris and the least was
Vigna angularis. The
Spinacia oleracea and
Amarantha dubius Thell seeds never germinated in 228 mg/kg simulated soil, depicting total inhibition of growth hormones. Lastly,
Vigna angularis was the only plant that germinated and grows (up-to 1 cm) in 456 mg/kg soil. This suggest that
Vigna angularis may be having a unique mechanism that enables it to absorb nutrients in a high chromium contaminated environment.
Vigna angularis and other plants in this study may have used a sulphate transporter mechanism to actively transport Cr(VI) into their body cells because it is an anion which is known to be very mobile because of its negative ions, and competes with sulphate ions in the uptake due to their similarities. In addition to that, these plants may have also used the passive sorption mechanism after reducing Cr(VI) absorbed in their tissues to Cr(III). This mechanism may have involved the diffusion of Cr(III) ions across the cell wall and plasma membranes into the plant’s body for translocation to other parts. These uptake mechanisms, as suggested in this study, are also supported by literature reports [
29,
30,
31,
32].
Cr transported to the aerial parts of these plants directly impacted cellular metabolism of shoots contributing to the reduction in their height, as seen in
Figure 2 of this study. This is in agreement with Sundaramoorthy and Sankarganesh [
14] and Bahira et al. [
17], who reported that high Cr(VI) concentrations (500 mg/kg) in soil affected shoot growth of wheat and oat. It also led to a decrease in plant height as reduced root growth was observed and, consequently, decreased nutrients and water transport to the higher parts of the plant.
The current study observed a significant variation in the germination and growth of edible vegetable seeds in Cr polluted soils (
Figure 3c). Simulated studies using Cr(VI) as the spiking agent to investigate different crop reactions and growth have been reported by several authors [
15,
33,
34]. Cr is considered strongly toxic because Cr(III) compounds in the soil are more or less insoluble and their ions are tightly bound to humus and clay particles while Cr(VI) is very soluble and easily passes through the plant cells into vacuoles where they combine with cations and form stable compounds which either accelerate or retard plants growth [
34].
Amarantha dubuis Thell and
Spinacia oleracea germinated in low concentration only while
Cicer arietinum, Phaseoulus vulgaris and
Vigna angularis germinated in both low and high concentration of Cr(VI). It may be possible to state here that seed coats of different plants’ impermeability and embryos’ selectivity could have affected the tolerance of chromium impacts. Those events naturally could have helped in the selection of high and low tolerant species or chromium varieties during the germination, early seedling stages and growth height in this study. These findings are in agreement with those reported by Kidd and Mart [
35], who reported that higher concentrations of heavy metal significantly reduced the strength of germination as compared to the lower concentrations, which had the least harmful effect on germination. Peralta et al. [
36] also found that there was no germination of spinach with applied Cr level at 320 mg/kg in the soil.
The consistent germination and significant growth of
Phaseoulus vulgaris,
Cicer arietinum and
Vigna angularis in chromium contaminated soil observed in this study seem to suggest that these plants might be tolerant or have mechanisms that allow them to germinate and grow in Cr toxic environment. It could be observed that at the roots of
Vigna angularis at the highest concentration was modified with fewer hair roots (Figure not shown) as compared to those from low concentrations and control. This could be the part of mechanism this plant applied in the roots to exclude excess Cr(VI) to its aerial parts, which made it possible for it to grow in such high concentration. Sharma et al. [
37] stated that toxic properties of Cr(VI) could be reduced by oxidising agents as well as from the formation of free radicals during the reduction of Cr(VI) to Cr(III) inside the root cells of plants.
To understand practically how these plants may have applied these strategies and mechanisms to survive in Cr polluted soil, the sampled plants in this study were divided into root, stem and leaf. Then measurement involving the assessment of different levels of Cr oxidation states; Ch
T, Cr(VI) and Cr(III) in the vegetables grown in the polluted soil was undertaken, and their occurrence is given in
Table 3. Different vegetable cultivars were found to differ in their ability to take up Cr oxidation states as the occurrence varied in this study, as depicted in
Table 3. This is in agreement with Fitz and Wenzel [
38], who reported that different plants exhibit diverse strategies to high concentrations of Cr, such as indicators, excluders and accumulators.
The mean values of chromium oxidation states in the sampled parts of edible vegetables and their controls were then subjected to Tukey’s test at confidence level (p ˂ 0.05). The values were found to have statistically significant differences in their control and parts (p = 0.000), indicating strong significant difference between the measured variables. This could be attributed to the Cr(VI) spiked in the experimental soils which could have been accumulated by these plants differentially through their tolerance mechanisms. In the roots, Cicer arietinum had the highest ChT while Spinacia oleracea had the lowest level. Vigna angularis had high Cr(VI) and Spinacia oleracea registered the lowest. The Cicer arietinum had highest Cr(III) while the lowest was in Amarantha dubuis Thell. This depicted that Cicer arietinum and Vigna angularis tolerance mechanisms for different Cr oxidation states were superior to Spinacia oleracea and Amarantha dubuis Thell in their roots, probably from the modification of their root hairs.
The level of total Cr translocated to the stem was high in Cicer arietinum and Vigna angularis and least in Spinacia oleracea. Vigna angularis accumulated high Cr(VI) in the stem as Spinacia oleracea registered nil in the stem. Cicer arietinum and Vigna angularis had a high concentration of Cr(III) in the stem, and the lowest level was recorded in Spinacia oleracea. Spinacia oleracea and Amarantha dubuis Thell maintained their low uptake implying either efficient exclusion or poor translocation. Vigna angularis seems efficient in the uptake of Cr(VI), suggesting that it has a unique mechanism of transporting it in the root and stem.
The level of ChT translocated to the leaf parts was highest in Cicer arietinum and Spinacia oleracea but least in Phaseoulus vulgaris. Cicer arietinum accumulated maximum Cr(VI) in the leaf and a very low concentration was observed in Spinacia oleracea. However, Spinacia oleracea had high levels of Cr(III) in the leaf, while the least concentration was found in Amarantha dubuis Thell. Cicer arietinum and Spinacia oleracea transported more Cr oxidation states to the leaf, which may be due to their leaf physiology, which encouraged phytovolarisation of the Cr pollutants from the plants.
Residual chromium concentrations (Ch
T, Cr(VI) and Cr(III)) were still detected in simulated soil at the end of the experiment, despite the effects of soil natural matrices and uptake by plants. This indicated that Cr is persistent in soil environments, and this is a source of concern. de Andrade et al. [
1] stated that the risk that could be associated with Cr(VI) amendment in soils may involve the chance of Cr persistence in soil and accumulation in plants that may eventually enter into the food chain. The concentrations derived were used to assess the transfer of Cr pollutants from soil to plants’ portion using pollution indices, such as BF and TF, as monitoring tools for pollution effect on edible vegetables grown in Cr contaminated sites.
3.3. Bioaccumulation/Bioconcentration Factor (BF/BCF)
In this study, Cr oxidation states levels revealed significant variations in the five studied vegetables, as shown in
Table 4. Significant differences (
p < 0.05) was found in Ch
T, Cr(VI) and Cr(III) levels between BF in the plant’s roots. Ch
T concentrations between
Amarantha dubuis Thell and
Phaseoulus vulgaris,
Spinacia oleracea and
Phaseoulus vulgaris,
Spinacia oleracea and
Cicer arietinum were significant while Cr(VI) significant levels were found between
Spinacia oleracea and
Phaseoulus vulgaris and
Spinacia oleracea and
Cicer arietinum. This suggested that these edible vegetables may have the potential to bioaccumulate and transfer chromium into their tissues. BF/BCF is an index that measures the ability of the plant to accumulate a particular metal with respect to its concentration in the soil and the root of the plant [
39]. Under normal conditions, the concentration of Cr in plants is supposed to be less than 0.001 mg/kg [
40]. Thus in terms of accumulation,
Cicer arietinum could be grouped as Cr moderate accumulator plants while
Vigna angularis,
Spinacia oleracea,
Phaseoulus vulgaris and
Amarantha dubuis Thell were low Cr accumulator plants based on the Malayeri et al. [
41] categorisation system. However, there were no non-accumulator and hyper-accumulator plants in this study. Instead, these plants could be grouped as phytoextractants or phytostabilisers. Since they are edible crops, this disqualifies them for phytoremediation of Cr contaminated sites.
The high values of BF/BCF confirmed that the roots are the main bio-accumulators of Cr in all its oxidation states. The highest Cr accumulation in the roots occurred due to their direct contact with Cr oxidation states in the soil. This Cr accumulation in the roots could be apportioned to that fraction of Cr ions physically adsorbed to the cell walls of the root and another fraction absorbed by the cells that were possibly immobilised in the root vacuoles. Since there was an increase in the shoot biomass, these mechanisms were suspected of having played a key role in the BF/BCF of the plants under toxic conditions in the soil.
Amaranthus and
Spinacia vegetables were found to have limited potential for the bioaccumulation of higher Cr concentration in their roots. That is why they were unable to germinate and grow in 228 and 456 mg/kg levels in this study. Oliveira [
40] and Chandra et al. [
42] explained that enhanced accumulation of chromium in the roots of
angularis and
arietinum species may have been due to the presence of organic acids (carboxylic acid and amino acids) in the root exudates which form complexes with chromium, thereby making them available for the uptake by roots while
Amaranthus and
Spinacia had low potential for accumulation of Cr. Rashed [
43] reiterated that the concentration of Cr is always higher in the roots than in the shoots. These differences in Cr accumulation in different parts of the plants suggested that different cellular mechanisms of bioaccumulation of Cr took place, and this influenced Cr bioaccumulation and partitioning in these plants, thus the need to find out the quantity that was translocated from the root to the shoot.
3.4. Translocation Factors
In this study, significant differences (
p < 0.05) were found in Ch
T, Cr(VI) and Cr(III) levels between TF in the plants stems and leaves. The significant differences trend between Ch
T and Cr(VI) concentrations shown in BF were also observed in TF. To confirm the distance moved by Ch
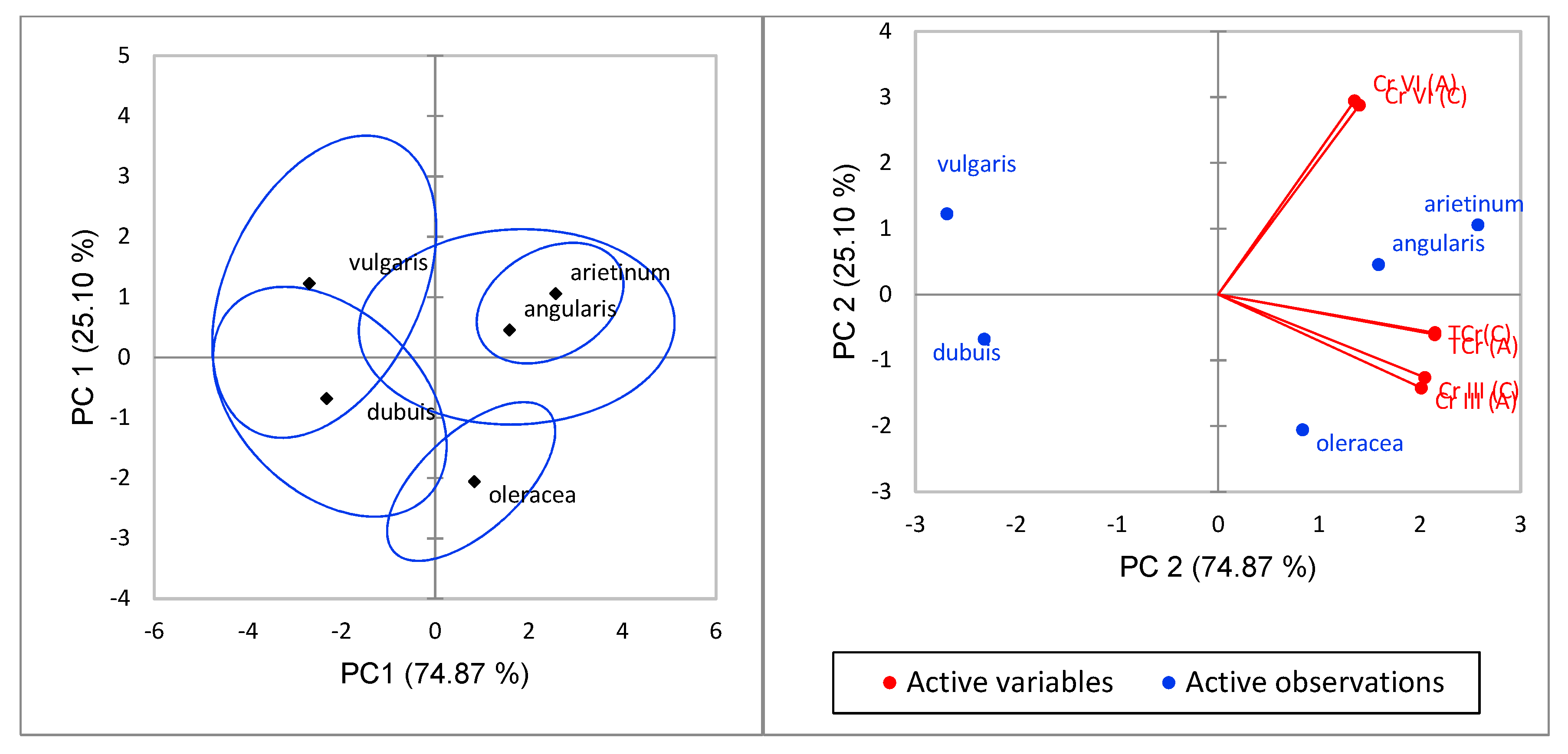
T, Cr(VI) and Cr(III) between these plant parts, further statistical analysis was done using principal component analysis (PCA). The relationship between the concentration of Ch
T, Cr(VI) and Cr(III) (active variables) in the stems (A) and leaves (C) of the edible vegetables species, such as
P. vulgaris A. dubuis,
S. oleracea,
V. angularis and
C. arietinum (active observations), for the first two principal components obtained (PC1 and PC2, which account for the 74.87% and the 25.10% of the total variance, respectively) are shown in
Figure 4. The active observations show that
Cicer arietinum and
Vigna angularis had a closer positive upper relationship than
Spinacia oleracea that had a lower positive relationship with far distant from the two.
Amarantha dubuis Thell and
Phaseoulus vulgaris had a negative relationship with
Phaseoulus vulgaris in the upper region and
Amarantha dubuis Thell in the lower area. The active variables showed that Cr(VI) was active in the upper positive region, while Ch
T and Cr(III) were active in the lower positive region. This depicted Cr(VI) to be more mobile than Ch
T and Cr(III).
Cicer arietinum and
Vigna angularis were observed to be likely related in their accumulation and transfer of chromium species. Other reported studies found out that Cr is absorbed by roots from nutrient solution as Cr(III) or Cr(VI) and translocated to aerial portions and roots, but it is largely retained in the roots [
44].
Given that these plants have shown bioaccumulation and translocation potential for Cr states in edible parts, they are of major health risk concern to the human population in Kenya and South Africa. Some of these plants have also been surveyed at tannery chromium dumpsites either growing wildly or grown near/at abandoned dumpsites by tannery workers in both countries, as shown in
Figure 3a,b. The tendency to retain more Cr in the root > leaf > stem was seen to be common in most of the plant species, but there was also quantitative translocation among the studied plant species. This may also mean that plants with high levels of Cr in their leaves may be trying to phytovolarise the metal into the atmosphere through their leaves, as seen by yellowing from the edge towards the petal and eventually dropping off the plant after possibly translocating it and starting a new leaf growth. This is in agreement with Khan et al. [
45], who reported that the maximum amount of Cr is accumulated in the roots followed by leaves and then fruits, which is close to findings in this study. According to Rashed [
43] dicotyledon species, such as
Vigna angularis, uptake and transport more Cr to shoots than monocotyledons plants, such as maize, due to differences in the rooting patterns, transpiration rates and metabolism between these two groups of plants while Cui et al. [
46] oppositely found that
Amaranthus dubius tolerated high Cr(VI) concentrations by accumulating and transferring them to aerial parts. The outcome of this study also compares to the reported findings by Oliveira et al. [
47], which found that an increase in Cr concentration in the leaves may be related to higher soil Cr concentration from the spiked soil and therefore, the metal was bioaccumulated from the roots to the leaves. Lower accumulation of Cr in leaves than in roots can be related to the conservation of photosynthesis processes from toxic levels of trace elements as well, according to Zojaji et al. [
48]. Thus, the levels of Cr(VI) transferred and detected in
Cicer arietinum,
Phaseoulus vulgaris,
Amarantha dubuis Thell and
Vigna angularis in the leaf portions of the plants signify high health risk to consumers.
3.5. Daily Intake of Chromium (ChT, Cr(VI), Cr(III)) through Edible Vegetables Grown on Cr(VI) Spiked Soils
This study focused on the cultivation of vegetables on Cr(VI) simulated soils and then estimated the average daily intake of Cr by adult and child consumers of these vegetables. A significant difference (
p < 0.05) was observed between child and adult consuming edible vegetables with Ch
T and Cr(III). This shows that edible vegetables in this study may have accumulated more Cr(VI) by converting it either to Ch
T or Cr(III) in various concentrations, but the uptake became greater than the maximum permissible limits of World Health Organisation (WHO)/Food Agriculture Organisation (FAO) [
49] for plant (0.10 mg/kg) in the spiked soil. In principle, the required amount of vegetables in people’s daily diet is supposed to be between 300 and 350 g per person’s (adult) and 200–230 g per child, as suggested by guidelines of WHO [
50].
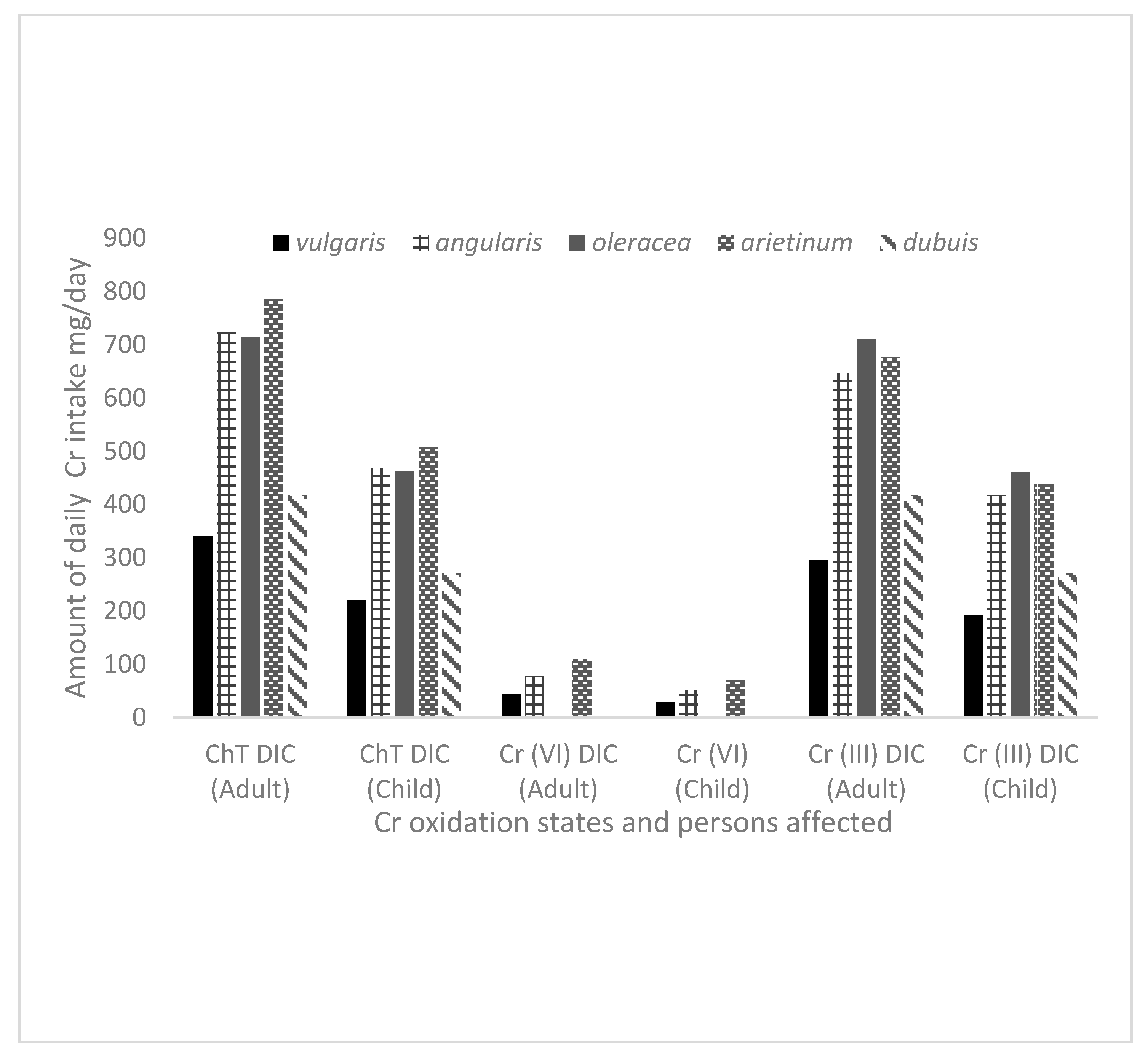
Figure 5 depicts the possible daily intake of high Ch
T, Cr(VI) and Cr(III) by humans from the edible vegetables grown in spiked soil in this study. The intake values were calculated by taking the average value of Cr oxidation states to assess the level exposure in all the five varieties of the vegetables (
Table 4) and taking into account that each adult and child in Kenya and South Africa (assuming 60 and 25 kg of body weight for adult and child, respectively) consumes approximately 340 and 220 g, respectively of vegetables per day according to WHO [
50]. However, the amounts in
Figure 5 give a picture showing that daily intake of vegetable species was far above recommended intake levels.
In the consumption habits of local residents of Kenya and South Africa,
Spinacia oleracea,
Cicer arietinum,
Phaseoulus vulgaris,
Amarantha dubuis Thell, and
Vigna angularis are consumed as leafy vegetables which accounts for 90% of total consumption of vegetables within the region with the remaining percentage being taken as seeds or pods [
4,
51]. This means that a very large population from these two countries are potentially at health risk due to exposure to these Ch
T, Cr(VI) and Cr(III). Therefore daily intake of chromium by human consumers of these vegetables is likely to expose them to these types of clinical disorders, i.e., respiratory, carcinogenic, renal, hepatic, gastrointestinal, cardiovascular, haematological, reproductive and developmental, genotoxic and mutagenic effects [
52].
3.7. Hazard Index
The HI method was used to assess the total of all potential health risks of Ch
T, Cr(VI) and Cr(III) accumulation through leafy vegetable consumption for adults and children. The risk is considered unacceptable at HI > 1. The results of the five leaf vegetables were all found to be above one, which may present a risk to adults and children in terms of Ch
T, Cr(VI) and Cr(III) exposure. HI values were observed in this decreasing order for adults and children as
Cicer arietinum > Vigna angularis > Spinacia oleracea > Amarantha dubuis Thell > Phaseoulus vulgaris, as shown in
Table 6. This suggested that the potential HI of Cr oxidation states through vegetable consumption were higher in
Cicer arietinum, being as high as 17.2 and 27.2 for adults and children, respectively. These results implied that the potential health risk of Cr oxidation states through the consumption of leafy vegetables was higher for all vegetable types. The estimation of HI, which takes care of the chemical mixtures, is very important in assessing multiple effects of heavy metals, such as Cr. In nature, chronic low-level intake of toxic metal elements can have a negative effect on human health, and the detrimental impact only becomes known after several years of exposure [
53].
The HI values of Cr oxidation states through vegetable consumption for children were higher than the values for adults in all vegetables. Therefore, Cr oxidation states are likely to contribute to the potential health risks of vegetable consumption for residents living and accessing sites contaminated with tannery Cr wastes. The HI findings in this study were lower than that of Chaturvedi et al. [
23], who found HI for Cr in children’s toys at 91.9. Huang et al. [
39] stated in their work that Cr speciation was of concern. This was because, according to them, health risk from Cr exposure may be overestimated if Cr(III) co-exists with Cr(VI). Cr(III) is considered essential in the metabolism of carbohydrates in animals, but high levels are equally risky under HI assessment, as seen in this study. Therefore, this study considered Cr speciation in vegetables from polluted sites and found out that their HI is a potential risk to both adults and children consuming them.

 ,
, 





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}