
The More Fertile, the More Creative: Changes in Women’s Creative Potential across the Ovulatory Cycle


Abstract
:1. Introduction
1.1. Signaling Theory
1.2. Courtship Displays Based on Parental Investment
1.3. Creativity as a Signal
1.4. Ovulatory Shifts and Creativity
2. Materials and Methods
2.1. Participants
2.2. Materials
2.2.1. Probability of Conception
2.2.2. Arousal
2.2.3. Divergent Thinking
2.3. Procedure
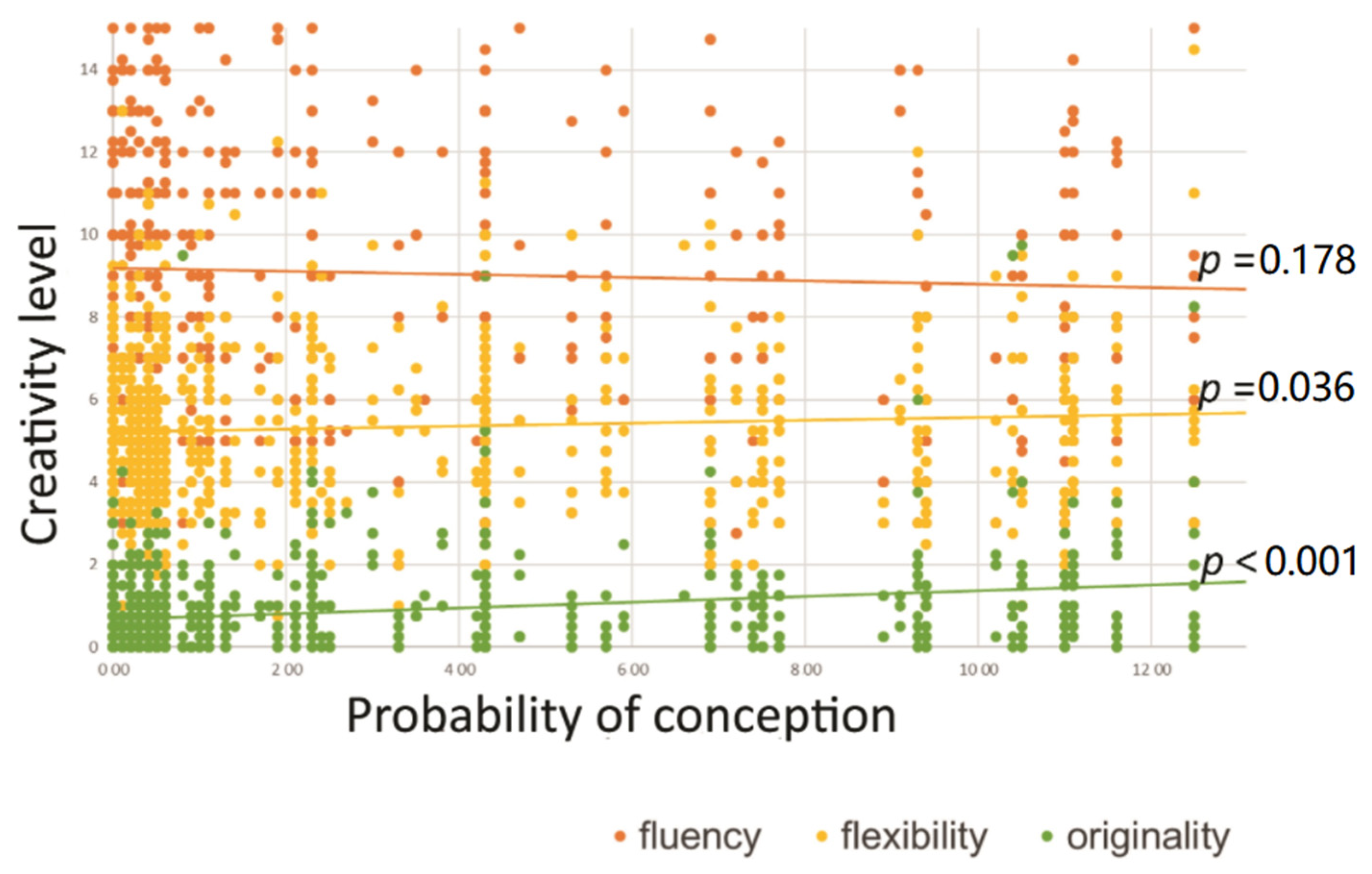
3. Results
3.1. Initial Analyses
3.2. Mediation Analysis: Arousal as a Mediator of the Relationship between Probability of Conception and Creative Originality
3.3. Mediation Analysis: Arousal as a Mediator of the Relationship between Probability of Conception and Creative Flexibility
4. Discussion
5. Limitations of the Study
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Richards, J.S. Ovulation. In Encyclopedia of Reproduction, 2nd ed.; Skinner, M.K., Ed.; Academic Press: Cambridge, MA, USA, 2018; Volume 2, pp. 92–98. [Google Scholar]
- Makhanova, A.; Miller, S.L. Female Fertility and Male Mating: Women’s Ovulatory Cues Influence Men’s Physiology, Cognition, and Behavior. Soc. Personal. Psychol. Compass 2013, 7, 389–400. [Google Scholar] [CrossRef]
- Bianchi, S.; Macchiarelli, G.; Micara, G.; Linari, A.; Boninsegna, C.; Aragona, C.; Rossi, G.; Cecconi, S.; Nottola, S.A. Ultrastructural markers of quality are impaired in human metaphase II aged oocytes: A comparison between reproductive and in vitro aging. J. Assist. Reprod. Genet. 2015, 32, 1343–1358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilcox, A.J.; Weinberg, C.R.; Baird, D.D. Timing of sexual intercourse in relation to ovulation. Effects on the probability of conception, survival of the pregnancy, and sex of the baby. N. Engl. J. Med. 1995, 333, 1517–1521. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, T.A.; Ahmed, S.M.; El-Gammal, Z.; Shouman, S.; Ahmed, A.; Mansour, R.; El-Badri, N. Oocyte Aging: The Role of Cellular and Environmental Factors and Impact on Female Fertility. Adv. Exp. Med. Biol. 2020, 1247, 109–123. [Google Scholar] [CrossRef] [PubMed]
- Conforti, A.; Mascia, M.; Cioffi, G.; De Angelis, C.; Copolla, G.; De Rosa, P.; Pivonello, R.; Alviggi, C.; De Placido, G. Air pollution and female fertility: A systematic review of literature. Reprod. Biol. Endocrinol. 2018, 16. [Google Scholar] [CrossRef] [PubMed]
- Farr, S.L.; Cooper, G.S.; Cai, J.; Savitz, D.A.; Sandler, D.P. Pesticide use and menstrual cycle characteristics among premenopausal women in the Agricultural Health Study. Am. J. Epidemiol. 2004, 160, 1194–1204. [Google Scholar] [CrossRef] [PubMed]
- Farquhar, C.M.; Bhattacharya, S.; Repping, S.; Mastenbroek, S.; Kamath, M.S.; Marjoribanks, J.; Boivin, J. Female subfertility. Nat. Rev. Dis. Primers 2019, 5, 7. [Google Scholar] [CrossRef] [PubMed]
- Durante, K.M.; Li, N.P.; Haselton, M.G. Changes in women’s choice of dress across the ovulatory cycle: Naturalistic and laboratory task-based evidence. Personal. Soc. Psychol. Bull. 2008, 34, 1451–1460. [Google Scholar] [CrossRef]
- Gangestad, S.W.; Thornhill, R.; Garver-Apgar, C.E. Adaptations to Ovulation: Implications for Sexual and Social Behavior. Curr. Dir. Psychol. Sci. 2005, 14, 312–316. [Google Scholar] [CrossRef]
- Macrae, C.N.; Alnwick, K.A.; Milne, A.B.; Schloerscheidt, A.M. Person perception across the menstrual cycle: Hormonal influences on social-cognitive functioning. Psychol. Sci. 2002, 13, 532–536. [Google Scholar] [CrossRef]
- Krug, R.; Finn, M.; Pietrowsky, R.; Fehm, H.L.; Born, J. Jealousy, general creativity, and coping with social frustration during the menstrual cycle. Arch. Sex. Behav. 1996, 25, 181–199. [Google Scholar] [CrossRef] [PubMed]
- Roney, J.R.; Simmons, Z.L. Men smelling women: Null effects of exposure to ovulatory sweat on men’s testosterone. Evol. Psychol. 2012, 10, 703–713. [Google Scholar] [CrossRef] [PubMed]
- Bullivant, S.B.; Sellergren, S.A.; Stern, K.; Spencer, N.A.; Jacob, S.; Mennella, J.A.; McClintock, M.K. Women’s sexual experience during the menstrual cycle: Identification of the sexual phase by noninvasive measurement of luteinizing hormone. J. Sex. Res. 2004, 41, 82–93. [Google Scholar] [CrossRef] [PubMed]
- Van Stein, K.R.; Strauß, B.; Brenk-Franz, K. Ovulatory Shifts in Sexual Desire but not Mate Preferences: An LH-Test-Confirmed, Longitudinal Study. Evol. Psychol. 2019, 17, 1474704919848116. [Google Scholar] [CrossRef] [PubMed]
- Marcinkowska, U.M.; Mijas, M.; Koziara, K.; Grebe, N.M.; Jasieńska, G. Variation in sociosexuality across natural menstrual cycles: Associations with ovarian hormones and cycle phase. Evol. Hum. Behav. 2020, 42, 35–42. [Google Scholar] [CrossRef]
- Little, A.C.; Jones, B.C.; Burriss, R.P. Preferences for masculinity in male bodies change across the menstrual cycle. Horm. Behav. 2007, 51, 633–639. [Google Scholar] [CrossRef] [Green Version]
- Thomas, A.G.; Armstrong, S.L.; Stewart-Williams, S.; Jones, B.C. Current Fertility Status Does Not Predict Sociosexual Attitudes and Desires in Normally Ovulating Women. Evol. Psychol. 2021, 19, 1474704920976318. [Google Scholar] [CrossRef] [PubMed]
- Stern, J.; Gerlach, T.M.; Penke, L. Probing Ovulatory-Cycle Shifts in Women’s Preferences for Men’s Behaviors. Psychol. Sci. 2020, 31, 424–436. [Google Scholar] [CrossRef] [Green Version]
- Jünger, J.; Motta-Mena, N.V.; Cardenas, R.; Bailey, D.; Rosenfield, K.A.; Schild, C.; Penke, L.; Puts, D.A. Do women’s preferences for masculine voices shift across the ovulatory cycle? Horm. Behav. 2018, 106, 122–134. [Google Scholar] [CrossRef] [Green Version]
- Wood, W.; Kressel, L.; Joshi, P.D.; Louie, B. Meta-Analysis of Menstrual Cycle Effects on Women’s Mate Preferences. Emot. Rev. 2014, 6, 229–249. [Google Scholar] [CrossRef] [Green Version]
- Gangestad, S.W.; Thornhill, R. Human oestrus. Proc. R. Soc. B Biol. Sci. 2008, 275, 991–1000. [Google Scholar] [CrossRef]
- Haselton, M.G.; Gildersleeve, K. Can Men Detect Ovulation? Curr. Dir. Psychol. Sci. 2011, 20, 87–92. [Google Scholar] [CrossRef]
- Bobst, C.; Lobmaier, J.S. Men’s preference for the ovulating female is triggered by subtle face shape differences. Horm. Behav. 2012, 62, 413–417. [Google Scholar] [CrossRef]
- Catena, T.M.; Simmons, Z.L.; Roney, J.R. Do women’s faces become more attractive near ovulation? Horm. Behav. 2019, 115, 104560. [Google Scholar] [CrossRef] [PubMed]
- Kirchengast, S.; Gartner, M. Changes in fat distribution (WHR) and body weight across the menstrual cycle. Coll. Antropol. 2002, 26, 47–57. [Google Scholar]
- Miller, G.; Tybur, J.M.; Jordan, B.D. Ovulatory cycle effects on tip earnings by lap dancers: Economic evidence for human estrus? Evol. Hum. Behav. 2007, 28, 375–381. [Google Scholar] [CrossRef] [Green Version]
- Puts, D.A.; Bailey, D.H.; Cardenas, R.A.; Burriss, R.P.; Welling, L.L.; Wheatley, J.R.; Dawood, K. Women’s attractiveness changes with estradiol and pro-gesterone across the ovulatory cycle. Horm. Behav. 2013, 63, 13–19. [Google Scholar] [CrossRef]
- Cobey, K.D.; Buunk, A.P.; Pollet, T.; Klipping, C.; Roberts, S.C. Men perceive their female partners, and themselves, as more attractive around ovulation. Biol. Psychol. 2013, 94, 513–516. [Google Scholar] [CrossRef]
- Fink, B.; Hugill, N.; Lange, B.P. Women’s body movements are a potential cue to ovulation. Pers. Individ. Differ. 2012, 53, 759–763. [Google Scholar] [CrossRef]
- Gildersleeve, K.A.; Haselton, M.G.; Larson, C.M.; Pillsworth, E.G. Body odor attractiveness as a cue of impending ovulation in women: Evidence from a study using hormone-confirmed ovulation. Horm. Behav. 2012, 61, 157–166. [Google Scholar] [CrossRef]
- Bryant, G.A.; Haselton, M.G. Vocal cues of ovulation in human females. Biol. Lett. 2009, 5, 12–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haselton, M.G.; Mortezaie, M.; Pillsworth, E.G.; Bleske-Rechek, A.; Frederick, D.A. Ovulatory shifts in human female ornamentation: Near ovulation, women dress to impress. Horm. Behav. 2007, 51, 40–45. [Google Scholar] [CrossRef]
- Haselton, M.G.; Miller, G.F. Women’s fertility across the cycle increases the short-term attractiveness of creative intelligence. Hum. Nat. 2006, 17, 50–73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Udry, J.R.; Morris, N.M. Variations in pedometer activity during menstrual cycle. Obstet. Gynecol. 1970, 35, 199–201. [Google Scholar]
- Fessler, D. No Time to Eat: An Adaptationist Account of Periovulatory Behavioral Changes. Q. Rev. Biol. 2003, 78, 3–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Šukolová, D.; Sarmány-Schuller, I. Fluctuating perception of selected risk situations with respect to hormonal changes during menstrual cycle. Studia Psychol. 2011, 53, 3–12. [Google Scholar]
- Piccoli, V.; Foroni, F.; Carnaghi, A. Comparing group dehumanization and intra-sexual competition among normally ovulating women and hormonal contraceptive users. Personal. Soc. Psychol. Bull. 2013, 39, 1600–1609. [Google Scholar] [CrossRef]
- Zhuang, J.Y.; Wang, J.X. Women ornament themselves for intrasexual competition near ovulation, but for intersexual attraction in luteal phase. PLoS ONE 2014, 9, e106407. [Google Scholar] [CrossRef] [PubMed]
- Lobmaier, J.S.; Bobst, C.; Probst, F. Can women detect cues to ovulation in other women’s faces? Biol. Lett. 2016, 12, 20150638. [Google Scholar] [CrossRef] [PubMed]
- Maner, J.K.; McNulty, J.K. Attunement to the fertility status of same-sex rivals: Women’s testosterone responses to olfactory ovulation cues. Evol. Hum. Behav. 2013, 34, 412–418. [Google Scholar] [CrossRef]
- Krug, R.; Stamm, U.; Pietrowsky, R.; Fehm, H.L.; Born, J. Effects of menstrual cycle on creativity. Psychoneuroendocrinology 1994, 19, 21–31. [Google Scholar] [CrossRef]
- Krug, R.; Mölle, M.; Fehm, H.L.; Born, J. Variations across the menstrual cycle in EEG activity during thinking and mental relaxation. J. Psychophysiol. 1999, 13, 163–172. [Google Scholar] [CrossRef]
- Miller, G.F. The Mating Mind: How Sexual Choice Shaped the Evolution of Human Nature; Doubleday: New York, NY, USA, 2000. [Google Scholar]
- Darwin, C. The Descent of Man and Selection in Relation to Sex; Murray: London, UK, 1871. [Google Scholar]
- Loyau, A.; Jalme, M.S.; Cagniant, C.; Sorci, G. Multiple sexual advertisements honestly reflect health status in peacocks (Pavo cristatus). Behav. Ecol. Sociobiol. 2005, 58, 552–557. [Google Scholar] [CrossRef]
- Zahavi, A. Mate selection—A selection for a handicap. J. Theor. Biol. 1975, 53, 205–213. [Google Scholar] [CrossRef]
- Grafen, A. Biological signals as handicaps. J. Theor. Biol. 1990, 144, 517–546. [Google Scholar] [CrossRef]
- Folstad, I.; Karter, A.J. Parasites, bright males, and the immunocompetence handicap. Am. Nat. 1992, 139, 603–622. [Google Scholar] [CrossRef]
- Andersson, M. Sexual Selection; Princeton University Press: Princeton, UK, 1994. [Google Scholar]
- Boogert, N.J.; Fawcett, T.W.; Lefebvre, L. Mate choice for cognitive traits: A review of the evidence in nonhuman vertebrates. Behav. Ecol. 2011, 22, 447–459. [Google Scholar] [CrossRef]
- Keagy, J.; Savard, J.-F.; Borgia, G. Male satin bowerbird problem-solving ability predicts mating success. Anim. Behav. 2009, 78, 809–817. [Google Scholar] [CrossRef]
- Wiggins, G.A.; Tyack, P.; Scharff, C.; Rohrmeier, M. The evolutionary roots of creativity: Mechanisms and motivations. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2015, 370, 20140099. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nowicki, S.; Peters, S.; Podos, J. Song Learning, Early Nutrition and Sexual Selection in Songbirds. Am. Zool. 1998, 38, 179–190. [Google Scholar] [CrossRef] [Green Version]
- Petrie, M. Improved growth and survival of offspring of peacocks with more elaborate trains. Nature 1994, 371, 598–599. [Google Scholar] [CrossRef]
- Clegg, N.; Nettle, D.; Miell, D. Status and mating success amongst visual artists. Front. Psychol. 2011, 2, 310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beaussart, M.L.; Kaufman, S.B.; Kaufman, J.C. Creative Activity, Personality, Mental Illness, and Short-Term Mating Success. J. Creat. Behav. 2012, 46, 151–167. [Google Scholar] [CrossRef]
- Griskevicius, V.; Cialdini, R.B.; Kenrick, D.T. Peacocks, Picasso, and parental investment: The effects of romantic motives on creativity. J. Personal. Soc. Psychol. 2006, 91, 63–76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harrison, A.; Moore, M.; Rucker, M. Further evidence on career and family compatibility among eminent women and men. Arch. Psicol. Neurol. Psichiatr. 1985, 46, 140–155. [Google Scholar]
- Feist, G. Natural and sexual selection in the evolutionary of creativity. Bull. Psychol. Arts 2001, 2, 11–16. [Google Scholar]
- Buss, D.M.; Schmitt, D.P. Sexual Strategies Theory: An evolutionary perspective on human mating. Psychol. Rev. 1993, 100, 204–232. [Google Scholar] [CrossRef] [PubMed]
- Trivers, R.L. Parental investment and sexual selection. In Sexual Selection and the Descent of Man; Campbell, B., Ed.; Aldine: Chicago, IL, USA, 1972; pp. 1871–1971. [Google Scholar]
- Stewart-Williams, S.; Thomas, A.G. The ape that thought it was a peacock: Does evolutionary psychology exaggerate human sex differences? Psychol. Inq. 2013, 24, 137–168. [Google Scholar] [CrossRef]
- Eens, M.; Pinxten, R. Sex-role reversal in vertebrates: Behavioral and endocrinological accounts. Behav. Process. 2000, 51, 135–147. [Google Scholar] [CrossRef]
- Lack, D. Ecological Adaptations for Breeding in Birds; Methuen: London, UK, 1968. [Google Scholar]
- Lassek, W.D.; Gaulin, S.J.C. Costs and benefits of fat-free muscle mass in men: Relationship to mating success, dietary requirements, and native immunity. Evol. Hum. Behav. 2009, 30, 322–328. [Google Scholar] [CrossRef]
- Hyde, J.S. The gender similarities hypothesis. Am. Psychol. 2005, 60, 581–592. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buss, D. Sex differences in human preferences: Evolutionary hypotheses tested in 37 cultures. Behav. Brain Sci. 1989, 12, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Lippa, R.A. The preferred traits of mates in a cross-national study of heterosexual and homosexual men and women: An examination of biological and cultural influences. Arch. Sex. Behav. 2007, 36, 193–208. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, W.C.; Putcha-Bhagavatula, A.; Miller, L.C. Are Men and Women Really That Different? Examining Some of Sexual Strategies Theory (SST)’s Key Assumptions about Sex-Distinct Mating Mechanisms. Sex Roles 2011, 64, 629–643. [Google Scholar] [CrossRef]
- Takeuchi, H.; Taki, Y.; Sassa, Y.; Hashizume, H.; Sekiguchi, S.; Fukushima, A.; Kawashima, R. White matter structures associated with creativity: Evidence from diffusion tensor imaging. Neuroimage 2010, 51, 11–18. [Google Scholar] [CrossRef]
- Koestler, A. The Act of Creation; Macmillan: New York, NY, USA, 1964. [Google Scholar]
- Prum, R.O. The Evolution of Beauty. How Darwin’s Forgotten Theory of Mate Choice Shapes the Animal World—and Us; Doubleday: New York, NY, USA, 2017. [Google Scholar]
- Sternberg, R.J.; Lubart, T.I. The Concept of Creativity: Prospects and Paradigms. In Handbook of Creativity; Sternberg, R.J., Ed.; Cambridge University Press: Cambridge, UK, 1999; pp. 3–15. [Google Scholar]
- Heilman, K.M.; Nadeau, S.E.; Beversdorf, D.O. Creative innovation: Possible brain mechanisms. Neurocase 2003, 9, 369–379. [Google Scholar] [CrossRef]
- Kaufman, S.B.; Kozbelt, A.; Bromley, M.L.; Miller, G.R. The Role of Creativity and Humor in Human Mate Selection. In Mating Intelligence: Sex, Relationships, and the Mind’s Reproductive System; Geher, G., Miller, G., Eds.; Lawrence Erlbaum Associates Publishers: Mahwah, NJ, USA, 2008; pp. 227–262. [Google Scholar]
- Runco, M.A.; Acar, S. Divergent Thinking as an Indicator of Creative Potential. Creat. Res. J. 2012, 24, 66–75. [Google Scholar] [CrossRef]
- Kaufman, J.C.; Beghetto, R.A. Beyond big and little: The four c model of creativity. Rev. Gen. Psychol. 2009, 13, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Acar, S.; Burnet, C.; Cabra, J.F. Ingredients of creativity: Originality and more. Creat. Res. J. 2017, 29, 133–144. [Google Scholar] [CrossRef]
- Runco, M.A. Creativity: Theories and Themes: Research, Development, and Practice; Elsevier Academic Press: Burlington, VT, USA, 2007. [Google Scholar]
- Jauk, E.; Benedek, M.; Dunst, B.; Neubauer, A.C. The relationship between intelligence and creativity: New support for the threshold hypothesis by means of empirical breakpoint detection. Intelligence 2013, 41, 212–221. [Google Scholar] [CrossRef] [Green Version]
- Gabora, L.; Kaufman, S.B. Evolutionary perspectives on creativity. In The Cambridge Handbook of Creativity; Kaufman, J., Sternberg, R., Eds.; Cambridge University Press: Cambridge, UK, 2010; pp. 279–300. [Google Scholar]
- Stojilovic, I. Effects of creativity on aesthetic experience. Psihologija 2017, 50, 319–339. [Google Scholar] [CrossRef] [Green Version]
- Miller, G.F. Aesthetic fitness: How sexual selection shaped artistic virtuosity as Evolutionary Approaches to Creativity 26 a fitness indicator and aesthetic preferences as mate choice criteria. Bull. Psychol. Arts 2001, 2, 20–25. [Google Scholar]
- Guilford, J.P. Creativity. Am. Psychol. 1950, 5, 444–454. [Google Scholar] [CrossRef] [PubMed]
- Kaufman, J.C.; Sternberg, R.J. (Eds.) The International Handbook of Creativity; Cambridge University Press: Cambridge, UK, 2006. [Google Scholar] [CrossRef]
- Dutton, D. The Art Instinct: Beauty, Pleasure and Human Evolution; Bloomsbury Press: New York, NY, USA, 2009. [Google Scholar]
- Kaufman, S.B.; Kozbelt, A.; Silvia, P.; Kaufman, J.C.; Ramesh, S.; Feist, G.J. Who finds Bill Gates sexy? Creative mate preferences as a function of cognitive ability, personality, and creative achievement. J. Creat. Behav. 2014, 50, 294–307. [Google Scholar] [CrossRef]
- Li, N.P.; Bailey, M.J.; Kenrick, D.T.; Linsenmeier, J.A. The necessities and luxuries of mate preferences: Testing the tradeoffs. J. Personal. Soc. Psychol. 2002, 82, 947–955. [Google Scholar] [CrossRef]
- Prokosch, M.D.; Coss, R.G.; Scheib, J.E.; Blozis, S.A. Intelligence and mate choice: Intelligent men are always appealing. Evol. Hum. Behav. 2009, 30, 11–20. [Google Scholar] [CrossRef]
- Roney, J.L.; Simmons, Z.L. Hormonal predictors of sexual motivation in natural menstrual cycles. Horm. Behav. 2013, 63, 636–645. [Google Scholar] [CrossRef]
- Storbeck, J.; Clore, G.L. Affective Arousal as Information: How Affective Arousal Influences Judgments, Learning, and Memory. Soc. Personal. Psychol. Compass 2008, 2, 1824–1843. [Google Scholar] [CrossRef] [Green Version]
- Yoest, K.E.; Cummings, J.A.; Becker, J.B. Estradiol, dopamine and motivation. Cent. Nerv. Syst. Agents Med. Chem. 2014, 14, 83–89. [Google Scholar] [CrossRef]
- Gangestad, S.W.; Haselton, M.G.; Welling, L.L.; Gildersleeve, K.; Pillsworth, E.G.; Burriss, R.P.; Puts, D.A. How valid are assessments of conception probability in ovulatory cycle research? Evaluations, recommendations, and theoretical implications. Evol. Hum. Behav. 2016, 37, 85–96. [Google Scholar] [CrossRef]
- Gonzales, J.E.; Ferrer, E. Efficacy of methods for ovulation estimation and their effect 823 on the statistical detection of ovulation-linked behavioral fluctuations. Behav. Res. Methods 2015, 824, 1–20. [Google Scholar] [CrossRef]
- Stirnemann, J.J.; Samson, A.; Bernard, J.P.; Thalabard, J.C. Day-specific probabilities of conception in fertile cycles resulting in spontaneous pregnancies. Hum. Reprod. 2013, 28, 1110–1116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bradley, M.M.; Lang, P.J. Measuring emotion: The Self-Assessment Manikin and the semantic differential. J. Behav. Ther. Exp. Psychiatry 1994, 25, 49–59. [Google Scholar] [CrossRef]
- Batey, M.; Rawles, R.; Furnham, A. Divergent Thinking and Interview Ratings. J. Psychoeduc. Assess. 2009, 27, 57–67. [Google Scholar] [CrossRef]
- Corbalan, J.; Lopez, J.J. Question Generation Test: A new procedure for measuring creativity. J. Creat. Behav. 1992, 26, 3–18. [Google Scholar]
- Nusbaum, E.C.; Silvia, P.J.; Beaty, R.E. Ready, set, create: What instructing people to “be creative” reveals about the meaning and mechanisms of divergent thinking. Psychol. Aesthet. Creat. Arts 2014, 8, 423–432. [Google Scholar] [CrossRef]
- Hayes, A.F. Methodology in the Social Sciences. Introduction to Mediation, Moderation, and Conditional Process Analysis: A Regression-based Approach; Guilford Press: New York, NY, USA, 2013. [Google Scholar]
- Barbot, B. The Dynamics of Creative Ideation: Introducing a New Assessment Paradigm. Front. Psychol. 2018, 9, 2529. [Google Scholar] [CrossRef] [PubMed]
- Gomez-Amor, J.; Martinez-Selva, J.M.; Roman, F.; Zamora, S. Electrodermal activity, hormonal levels and subjective experience during the menstrual cycle. Biol. Psychol. 1990, 30, 125–139. [Google Scholar] [CrossRef]
- Colzato, L.S.; Hertsig, G.; Van Den Wildenberg, W.P.M.; Hommel, B. Estrogen modulates inhibitory control in healthy human females: Evidence from z stop-signal paradigm. Neuroscience 2010, 167, 709–715. [Google Scholar] [CrossRef]
- Baas, M.; De Dreu, C.K.; Nijstad, B.A. A meta-analysis of 25 years of mood-creativity research: Hedonic tone, activation, or regulatory focus? Psychol. Bull. 2008, 134, 779–806. [Google Scholar] [CrossRef]
- Stanton, S.J.; Schultheiss, O.C. Basal and dynamic relationships between implicit power motivation and estradiol in women. Horm. Behav. 2007, 52, 571–580. [Google Scholar] [CrossRef] [PubMed]
- Gino, F.; Ariely, D. The dark side of creativity: Original thinkers can be more dishonest. J. Personal. Soc. Psychol. 2012, 102, 445–459. [Google Scholar] [CrossRef] [Green Version]
- Fisher, M.L. Female intrasexual competition decreases female facial attractiveness. Proc. Biol. Sci. 2004, 271, S283–S285. [Google Scholar] [CrossRef] [Green Version]
- Mafra, A.L.; Varella, M.A.C.; Defelipe, R.P.; Anchieta, N.M.; de Almeida, C.A.; Valentova, J.V. Makeup usage in women as a tactic to attract mates and compete with rivals. Personal. Individ. Differ. 2020, 163, 110042. [Google Scholar] [CrossRef]
- Jungheim, E.S.; Moley, K.H. Current knowledge of obesity’s effects in the pre- and periconceptional periods and avenues for future research. Am. J. Obstet. Gynecol. 2010, 203, 525–530. [Google Scholar] [CrossRef] [Green Version]
- Dale, P.O.; Tanbo, T.; Haug, E.; Abyholm, T. The impact of insulin resistance on the outcome of ovulation induction with low-dose follicle stimulating hormone in women with polycystic ovary syndrome. Hum. Reprod. 1998, 13, 567–570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bianchi, S.; Nottola, S.A.; Torge, D.; Palmerini, M.G.; Necozione, S.; Macchiarelli, G. Association between Female Reproductive Health and Mancozeb: Systematic Review of Experimental Models. Int. J. Environ. Res. Public Health 2020, 17, 2580. [Google Scholar] [CrossRef] [Green Version]
- Warren, A.M.; Gurvich, C.; Worsley, R.; Kulkarni, J. A systematic review of the impact of oral contraceptives on cognition. Contraception 2014, 90, 111–116. [Google Scholar] [CrossRef]
- Pletzer, B.A.; Kerschbaum, H.H. 50 years of hormonal contraception-time to find out, what it does to our brain. Front. Neurosci. 2014, 8, 256. [Google Scholar] [CrossRef] [Green Version]
- Robinson, S.A.; Dowell, M.; Pedulla, D.; McCauley, L. Do the emotional side-effects of hormonal contraceptives come from pharmacologic or psychological mechanisms? Med. Hypotheses 2004, 63, 268–273. [Google Scholar] [CrossRef]
- Ashby, F.G.; Isen, A.M.; Turken, A.U. A neuropsychological theory of positive affect and its influence on cognition. Psychol. Rev. 1999, 106, 529–550. [Google Scholar] [CrossRef] [PubMed]
- Haselton, M.G.; Gangestad, S.W. Conditional expression of women’s desires and men’s mate guarding across the ovulatory cycle. Horm. Behav. 2006, 49, 509–518. [Google Scholar] [CrossRef] [PubMed]
- Pillsworth, E.G.; Haselton, M.G. Male sexual attractive-ness predicts differential ovulatory shifts in female extra-pairattraction and malemate retention. Evol. Hum. Behav. 2006, 27, 247–258. [Google Scholar] [CrossRef]
- Pillsworth, E.G.; Haselton, M.G.; Buss, D.M. Ovulatory shifts in female sexual desire. J. Sex Res. 2004, 41, 55–65. [Google Scholar] [CrossRef] [PubMed]
- Vartanian, O.; Saint, S.A.; Herz, N.; Suedfeld, P. The Creative Brain under Stress: Considerations for Performance in Extreme Environments. Front. Psychol. 2020, 11, 585969. [Google Scholar] [CrossRef]
- Palomba, S.; Daolio, J.; Romeo, S.; Battaglia, F.A.; Marci, R.; La Sala, G.B. Lifestyle and fertility: The influence of stress and quality of life on female fertility. Reprod. Biol. Endocrinol. 2018, 16, 113. [Google Scholar] [CrossRef] [Green Version]
- Moriarty, O.; McGuire, B.E.; Finn, D.P. The effect of pain on cognitive function: A review of clinical and preclinical research. Prog. Neurobiol. 2011, 93, 385–404. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
| Variables | M | SD | 1 | 2 | 3 | 4 | 5 |
|---|---|---|---|---|---|---|---|
| 1. Creative fluency | 9.06 | 4.67 | |||||
| 2. Creative flexibility | 5.32 | 2.1 | 0.79 *** | ||||
| 3. Creative originality | 0.9 | 1.13 | 0.49 *** | 0.53 *** | |||
| 4. Arousal | 2.67 | 0.9 | −0.00 | 0.04 | 0.02 | ||
| 5. Valence | 3.5 | 0.87 | −0.00 | −0.01 | 0.01 | 0.18 *** | |
| 6. Probability of conception | 3.15 | 3.85 | −0.04 | 0.07 * | 0.24 *** | 0.05 | 0.00 |
| Pathways | Originality | Flexibility | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| b | SE | t | p | CI 95% | b | SE | t | p | CI 95% | |||
| LL | UL | LL | UL | |||||||||
| Direct | ||||||||||||
| a | 0.012 | 0.009 | 1.414 | 0.158 | −0.005 | 0.029 | 0.012 | 0.009 | 1.414 | 0.158 | −0.005 | 0.029 |
| b | 0.017 | 0.045 | 0.383 | 0.702 | −0.071 | 0.105 | 0.099 | 0.085 | 1.175 | 0.240 | −0.067 | 0.267 |
| c’ | 0.069 | 0.011 | 6.545 | <0.001 | 0.048 | 0.089 | 0.035 | 0.020 | 1.748 | 0.081 | −0.004 | 0.074 |
| c | 0.069 | 0.011 | 6.578 | <0.001 | 0.048 | 0.090 | 0.036 | 0.020 | 1.811 | 0.071 | −0.003 | 0.075 |
| Indirect | ||||||||||||
| 0.000 | 0.001 | −0.001 | 0.002 | 0.001 | 0.002 | −0.001 | 0.005 | |||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Galasinska, K.; Szymkow, A. The More Fertile, the More Creative: Changes in Women’s Creative Potential across the Ovulatory Cycle. Int. J. Environ. Res. Public Health 2021, 18, 5390. https://doi.org/10.3390/ijerph18105390
Galasinska K, Szymkow A. The More Fertile, the More Creative: Changes in Women’s Creative Potential across the Ovulatory Cycle. International Journal of Environmental Research and Public Health. 2021; 18(10):5390. https://doi.org/10.3390/ijerph18105390
Chicago/Turabian StyleGalasinska, Katarzyna, and Aleksandra Szymkow. 2021. "The More Fertile, the More Creative: Changes in Women’s Creative Potential across the Ovulatory Cycle" International Journal of Environmental Research and Public Health 18, no. 10: 5390. https://doi.org/10.3390/ijerph18105390