
β-Glucans Could Be Adjuvants for SARS-CoV-2 Virus Vaccines (COVID-19)

 ,
, 
 and
and 
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Systemic Impact of COVID-19 Infection
3. The Cytokine Response during COVID-19 Infection
4. β-Glucans
4.1. Initial Immunomodulatory Effect of β-Glucans
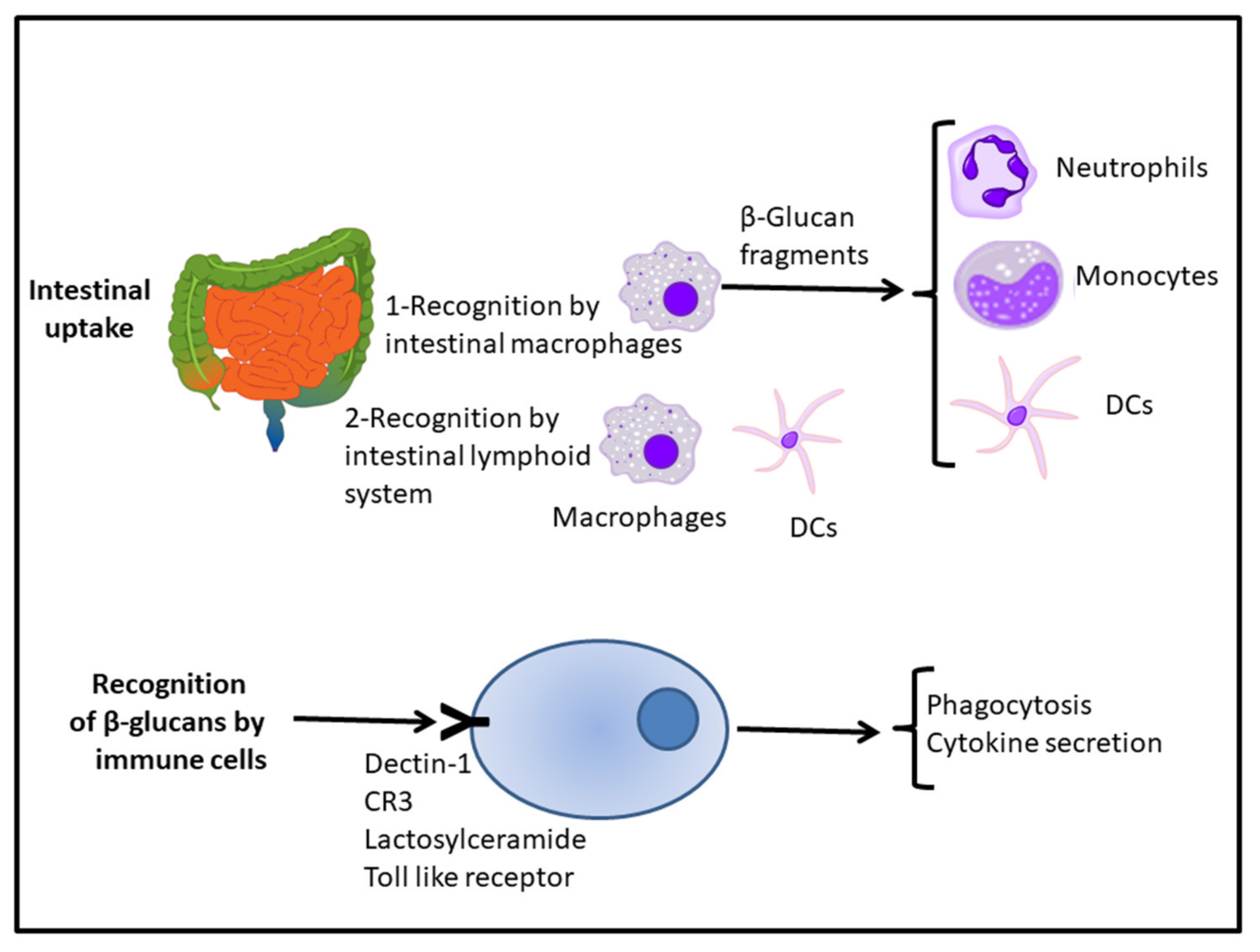
4.2. Receptors for β-Glucans
4.3. Immunostimulatory Activity of β-Glucans
4.4. β-Glucans against Infection
4.5. β-Glucans as Immunomodulators
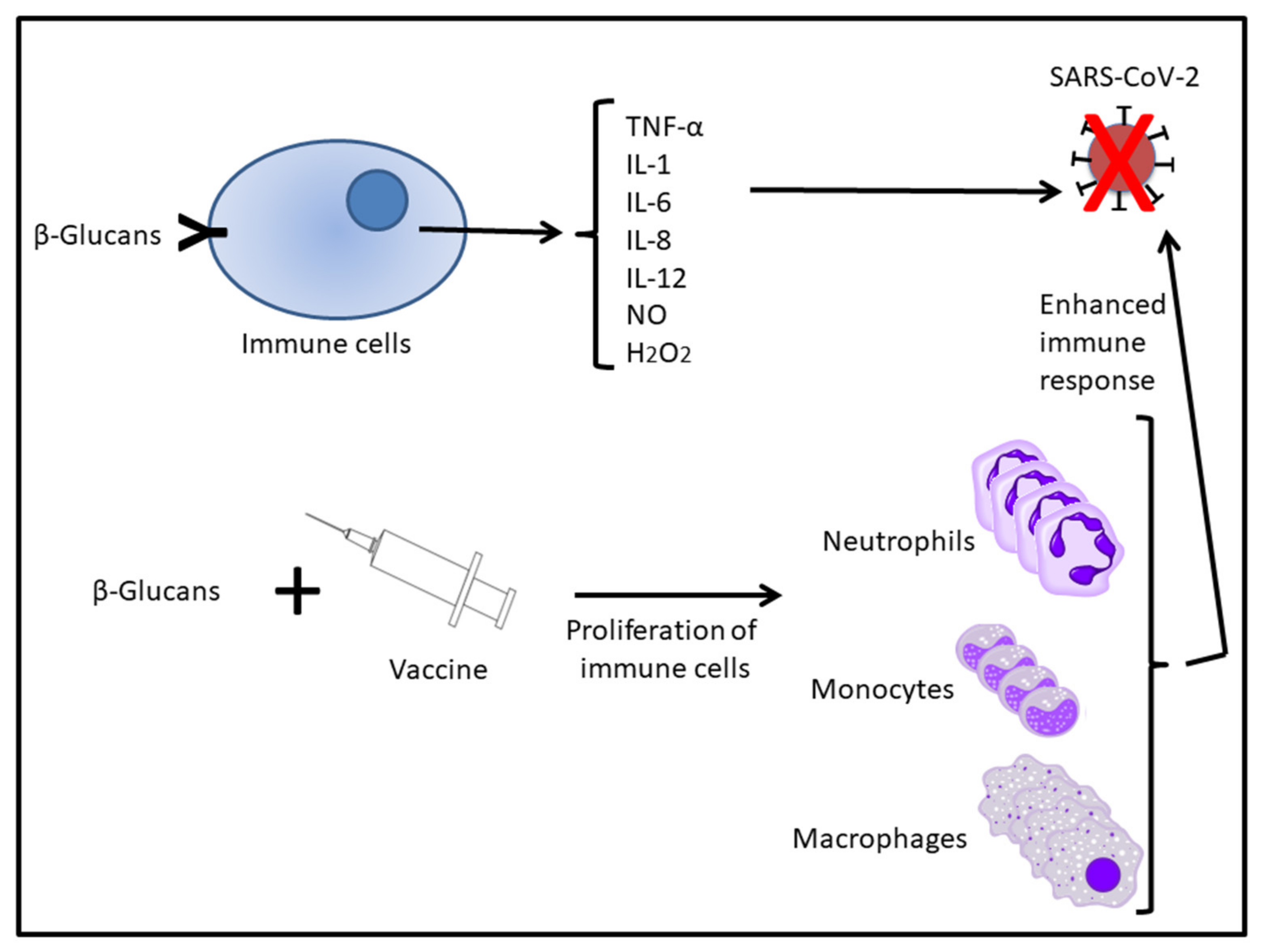
5. Glucans Complementing the Action of Vaccines
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhu, N.; Zhang, D.; Wang, W.; Li, X.; Yang, B.; Song, J.; Zhao, X.; Huang, B.; Shi, W.; Lu, R.; et al. China Novel Coronavirus Investigating and Research Team. A novel coronavirus from patients with pneumonia in China. N. Engl. J. Med. 2020, 382, 727–733. [Google Scholar] [CrossRef] [PubMed]
- Sanders, J.M.; Monogue, M.L.; Jodlowski, T.Z.; Cutrell, J.B. Pharmacologic treatments for coronavirus disease 2019 (COVID-19): A review. Jama 2020, 323, 1824–1836. [Google Scholar] [CrossRef]
- Callaway, E. The race for coronavirus vaccines: A graphical guide. Nat. Cell Biol. 2020, 580, 576–577. [Google Scholar] [CrossRef]
- Yang, L.; Liu, S.; Liu, J.; Zhang, Z.; Wan, X.; Huang, B.; Chen, Y.; Zhang, Y. COVID-19: Immunopathogenesis and Immunotherapeutics. Signal Transduct. Target. Ther. 2020, 5, 1–8. [Google Scholar] [CrossRef]
- Tregoning, J.S.; Brown, E.S.; Cheeseman, H.M.; Flight, K.E.; Higham, S.L.; Lemm, N.-M.; Pierce, B.F.; Stirling, D.C.; Wang, Z.; Pollock, K.M. Vaccines for COVID-19. Clin. Exp. Immunol. 2020, 202, 162–192. [Google Scholar] [CrossRef] [PubMed]
- Forni, G.; Mantovani, A. COVID-19 vaccines: Where we stand and challenges ahead. Cell Death Differ. 2021, 28, 626–639. [Google Scholar] [CrossRef] [PubMed]
- Krammer, F. SARS-CoV-2 vaccines in development. Nature 2020, 586, 516–527. [Google Scholar] [CrossRef]
- Vetvicka, V.; Vannucci, L.; Sima, P.; Richter, J. Beta glucan: Supplement or drug? From laboratory to clinical trials. Molecules 2019, 24, 1251. [Google Scholar] [CrossRef] [Green Version]
- Vetvicka, V.; Vetvickova, J. Natural immunomodulators and their stimulation of immune reaction: True or false? Anticancer. Res. 2014, 34, 2275–2282. [Google Scholar] [PubMed]
- Coffman, R.L.; Sher, A.; Seder, R.A. Vaccine adjuvants: Putting innate immunity to work. Immunity 2010, 33, 492–503. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fischer, G.; Horton, R.E.; Edelman, R. From the national institute of allergy and infectious diseases. Summary of an international symposium on potentiation of the immune response to vaccines. J. Infect. Dis. 1980, 141, 163–166. [Google Scholar] [CrossRef]
- Cox, J.C.; Coulter, A.R. Adjuvants—A classification and review of their modes of action. Vaccine 1997, 15, 248–256. [Google Scholar] [CrossRef]
- Edelman, R. An update on vaccine adjuvants in clinical trial. AIDS Res. Hum. Retroviruses 1992, 8, 1409–1411. [Google Scholar] [CrossRef]
- Geller, A.; Yan, J. Could the induction of trained immunity by β-glucan serve as a defense against COVID-19? Front. Immunol. 2020, 11, 1782. [Google Scholar] [CrossRef]
- Tartof, S.Y.; Slezak, J.M.; Fischer, H.; Hong, V.; Ackerson, B.K.; Ranasinghe, O.N.; Frankland, T.B.; Ogun, O.A.; Zamparo, J.M.; Gray, S.; et al. Effectiveness of mRNA BNT162b2 COVID-19 vaccine up to 6 months in a large integrated health system in the USA: A retrospective cohort study. Lancet 2021, 398, 1407–1416. [Google Scholar] [CrossRef]
- Genç, R.; Yakuboğullari, N.; Nalbantsoy, A.; Çöven, F.; Bedir, E. Adjuvant potency of Astragaloside VII embedded cholesterol nanoparticles for H3N2 influenza vaccine. Turk. J. Boil. 2020, 44, 304–314. [Google Scholar] [CrossRef] [PubMed]
- Córdova, A.; Alvárez de Mon, M. Inmunidad en el Deporte; GYMNOS: Madrid, Spain, 2001. [Google Scholar]
- Vardhana, S.A.; Wolchok, J.D. The many faces of the anti-COVID immune response. J. Exp. Med. 2020, 217, e20200678. [Google Scholar] [CrossRef]
- De La Concha, E.G. Role of the immune response in COVID-19. An. RANM 2020, 137, 113–116. [Google Scholar] [CrossRef]
- Merad, M.; Martin, J.C. Author correction: Pathological inflammation in patients with COVID-19: A key role for monocytes and macrophages. Nat. Rev. Immunol. 2020, 20, 448. [Google Scholar] [CrossRef]
- Hu, B.; Huang, S.; Yin, L. The cytokine storm and COVID-19. J. Med. Virol. 2021, 93, 250–256. [Google Scholar] [CrossRef] [PubMed]
- Pontelli, M.C.; Castro, I.A.; Martins, R.B.; Veras, F.P.; La Serra, L.; Nascimento, D.C.; Cardoso, R.S.; Rosales, R.; Lima, T.M.; Souza, J.P.; et al. Infection of human lymphomononuclear cells by SARS-CoV-2. bioRxiv 2020, 11, 1–7. [Google Scholar] [CrossRef]
- Zheng, F.; Zhou, Y.; Zhou, Z.; Ye, F.; Huang, B.; Huang, Y.; Ma, J.; Zuo, Q.; Tan, X.; Xie, J.; et al. SARS-CoV-2 clearance in COVID-19 patients with Novaferon treatment: A randomized, open-label, parallel-group trial. Int. J. Infect. Dis. 2020, 99, 84–91. [Google Scholar] [CrossRef] [PubMed]
- Majano, P.; Roda-Navarro, P.; Alonso-Lebrero, J.L.; Brieva, A.; Casal, C.; Pivel, J.P.; Lopez-Cabrera, M.; Moreno-Otero, R. AM3 inhibits HBV replication through activation of peripheral blood mononuclear cells. Int. Immunopharmacol. 2004, 4, 921–927. [Google Scholar] [CrossRef]
- Prompetchara, E.; Ketloy, C.; Palaga, T. Immune responses in COVID-19 and potential vaccines: Lessons learned from SARS and MERS epidemic. Asian Pac. J. Allergy Immunol. 2020, 38, 1–9. [Google Scholar] [CrossRef]
- Cordova, A. Fisiologia Dinámica; Elsevier-Masson: Barcelona, Spain, 2003. [Google Scholar]
- Pourahmad, J.; Salimi, A. Isolated human peripheral blood mononuclear cell (PBMC), a cost effective tool for predicting immunosuppressive effects of drugs and xenobiotics. Iran. J. Pharm. Res. 2015, 14, 979–980. [Google Scholar]
- Tai, Y.; Wang, Q.; Korner, H.; Zhang, L.; Wei, W. Molecular mechanisms of T cells activation by dendritic cells in autoimmune diseases. Front. Pharmacol. 2018, 9, 642. [Google Scholar] [CrossRef]
- Li, L.; Wo, J.; Shao, J.; Zhu, H.; Wu, N.; Li, M.; Yao, H.; Hu, M.; Dennin, R.H. SARS-coronavirus replicates in mononuclear cells of peripheral blood (PBMCs) from SARS patients. J. Clin. Virol. 2003, 28, 239–244. [Google Scholar] [CrossRef]
- Brufsky, A.; Lotze, M.T. DC/L-SIGNs of hope in the COVID-19 pandemic. J. Med. Virol. 2020, 92, 1396–1398. [Google Scholar] [CrossRef]
- Cui, W.; Fan, Y.; Wu, W.; Zhang, F.; Wang, J.; Ni, A. Expression of lymphocytes and lymphocyte subsets in patients with severe acute respiratory syndrome. Clin. Infect. Dis. 2003, 37, 857–859. [Google Scholar] [CrossRef] [PubMed]
- D’Alessio, F.R.; Heller, N.M. COVID-19 and myeloid cells: Complex interplay correlates with lung severity. J. Clin. Investig. 2020, 130, 6214–6217. [Google Scholar] [CrossRef]
- Johansson, S.M.; Admyre, C.; Scheynius, A.; Gabrielsson, S. Different types of in vitro generated human monocyte-derived dendritic cells release exosomes with distinct phenotypes. Immunology 2008, 123, 491–499. [Google Scholar] [CrossRef]
- Garcia-Vallejo, J.J.; van Kooyk, Y. The physiological role of DC-SIGN: A tale of mice and men. Trends Immunol. 2013, 34, 482–486. [Google Scholar] [CrossRef] [PubMed]
- Hammer, Q.; Rückert, T.; Romagnani, C. Natural killer cell specificity for viral infections. Nat. Immunol. 2018, 19, 800–808. [Google Scholar] [CrossRef] [PubMed]
- Serrano-Gómez, D.; Martinez-Nunez, R.T.; Sierra-Filardi, E.; Izquierdo-Useros, N.; Colmenares, M.; Pla, J.; Rivas, L.; Martinez-Picado, J.; Jimenez-Barbero, J.; Alonso-Lebrero, J.L.; et al. AM3 modulates dendritic cell pathogen recognition capabilities by targeting DC-SIGN. Antimicrob. Agents Chemother. 2007, 51, 2313–2323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, C.; Wang, X.-M.; Li, S.-R.; Twelkmeyer, T.; Wang, W.-H.; Zhang, S.-Y.; Wang, S.-F.; Chen, J.-Z.; Jin, X.; Wu, Y.-Z.; et al. NKG2A is a NK cell exhaustion checkpoint for HCV persistence. Nat. Commun. 2019, 10, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Fauriat, C.; Long, E.O.; Ljunggren, H.-G.; Bryceson, Y. Regulation of human NK-cell cytokine and chemokine production by target cell recognition. Blood J. Am. Soc. Hematol. 2010, 115, 2167–2176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moya, P.; Baixeras, E.; Barasoain, I.; Rojo, J.M.; Ronda, E.; Alonso, M.L.; Portolés, A. Inmunoferon (Am3) enhances the activities of early–type interferon inducers and natural killer cells. Immunopharmacol. Immunotoxicol. 1987, 9, 243–256. [Google Scholar] [CrossRef]
- Campbell, J.B.; Grunberger, T.; Kochman, M.A.; White, S.L. A microplaque reduction assay for human and mouse interferon. Can. J. Microbiol. 1975, 21, 1247–1253. [Google Scholar] [CrossRef]
- Gruber, C. Impaired interferon signature in severe COVID-19. Nat. Rev. Immunol. 2020, 20, 353. [Google Scholar] [CrossRef]
- Demaria, O.; Carvelli, J.; Batista, L.; Thibult, M.-L.; Morel, A.; André, P.; Morel, Y.; Vély, F.; Vivier, E. Identification of druggable inhibitory immune checkpoints on Natural Killer cells in COVID-19. Cell. Mol. Immunol. 2020, 17, 995–997. [Google Scholar] [CrossRef]
- Rajaram, S.; Canaday, L.; Ochayon, D.E.; Rangel, K.M.; Ali, A.; Gyurova, I.E.; Krishnamurthy, D.; Fletcher, J.S.; Reighard, S.D.; Cox, A.; et al. The promise and peril of natural killer cell therapies in pulmonary infection. Immunology 2020, 52, 887–889. [Google Scholar] [CrossRef]
- Córdova, A.; Sureda, A.; Pons, A.; Alvarez-Mon, M. Modulation of TNF-α, TNF-α receptors and IL-6 after treatment with AM3 in professional cyclists. J. Sports Med. Phys. Fitness 2014, 55, 345–351. [Google Scholar]
- Cordova, A.; Monserrat, J.; Villa, G.; Reyes, E.; Soto, M.A.-M. Effects of AM3 (Inmunoferon®) on increased serum concentrations of interleukin-6 and tumour necrosis factor receptors I and II in cyclists. J. Sports Sci. 2006, 24, 565–573. [Google Scholar] [CrossRef]
- Brieva, A.; Guerrero, A.; Alonso-Lebrero, J.; Pivel, J. Inmunoferon®, a glycoconjugate of natural origin, inhibits LPS-induced TNF-α production and inflammatory responses. Int. Immunopharmacol. 2001, 1, 1979–1987. [Google Scholar] [CrossRef]
- Brieva, A.; Guerrero, A.; Pivel, J. Inmunoferon®, a glycoconjugate of natural origin, regulates the liver response to inflammation and inhibits TNF-α production by an HPA axis-dependent mechanism. Int. Immunopharmacol. 2002, 2, 807–813. [Google Scholar] [CrossRef]
- Martínez, A.C.; Lázaro, D.F. New trends in biological aids to recovery after exercise: Immunomodulators. J. Hum. Sport Exerc. 2018, 13, 116–128. [Google Scholar] [CrossRef] [Green Version]
- Catanzaro, M.; Fagiani, F.; Racchi, M.; Corsini, E.; Govoni, S.; Lanni, C. Immune response in COVID-19: Addressing a pharmacological challenge by targeting pathways triggered by SARS-CoV-2. Signal Transduct. Target. Ther. 2020, 5, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Barbosa, M.L.D.C.; Fumian, M.M.; Lima, L.M.; De Miranda, A.L.P.; Barreiro, E.J. Therapeutic approaches for tumor necrosis factor inhibition. Braz. J. Pharm. Sci. 2011, 47, 427–446. [Google Scholar] [CrossRef]
- Hoffman, O.A.; Standing, J.E.; Limper, A.H. Pneumocystis carinii stimulates tumor necrosis factor-alpha release from alveolar macrophages through a beta-glucan-mediated mechanism. J. Immunol. 1993, 150, 3932–3940. [Google Scholar]
- Xu, X.; Yasuda, M.; Nakamura-Tsuruta, S.; Mizuno, M.; Ashida, H. β-Glucan from lentinus edodes inhibits nitric oxide and tumor necrosis factor-α production and phosphorylation of mitogen-activated protein kinases in lipopolysaccharide-stimulated Murine RAW 264.7 macrophages. J. Biol. Chem. 2012, 287, 871–878. [Google Scholar] [CrossRef] [Green Version]
- Loo, Y.-M.; Fornek, J.; Crochet, N.; Bajwa, G.; Perwitasari, O.; Martinez-Sobrido, L.; Akira, S.; Gill, M.A.; Garcia-Sastre, A.; Katze, M.G.; et al. Distinct RIG-I and MDA5 signaling by RNA viruses in innate immunity. J. Virol. 2008, 82, 335–345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Kurt-Jones, E.A.; Finberg, R.W. Innate immunity to respiratory viruses. Cell. Microbiol. 2007, 9, 1641–1646. [Google Scholar] [CrossRef]
- Li, G.; Fan, Y.; Lai, Y.; Han, T.; Li, Z.; Zhou, P.; Pan, P.; Wang, W.; Hu, D.; Liu, X.; et al. Coronavirus infections and immune responses. J. Med. Virol. 2020, 92, 424–432. [Google Scholar] [CrossRef]
- Le Page, C.; Génin, P.; Baines, M.G.; Hiscott, J. Interferon activation and innate immunity. Rev. Immunogenet. 2000, 2, 374–386. [Google Scholar]
- Hadjadj, J.; Yatim, N.; Barnabei, L.; Corneau, A.; Boussier, J.; Smith, N.; Péré, H.; Charbit, B.; Bondet, V.; Chenevier-Gobeaux, C.; et al. Impaired type I Interferon activity and exacerbated inflammatory responses in severe COVID-19 patients. Science 2020, 369, 718–724. [Google Scholar] [CrossRef] [PubMed]
- Vidal, S.M.; Khakoo, S.I.; Biron, C.A. Natural killer cell responses during viral infections: Flexibility and conditioning of innate immunity by experience. Curr. Opin. Virol. 2011, 1, 497–512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, G.; Wu, D.; Guo, W.; Cao, Y.; Huang, D.; Wang, H.; Wang, T.; Zhang, X.; Chen, H.; Yu, H.; et al. Clinical and immunologic features in severe and moderate forms of coronavirus disease. J. Clin. Investig. 2020, 130, 2620–2629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lagunas-Rangel, F.A.; Chávez-Valencia, V. High IL-6/IFN-γ ratio could be associated with severe disease in COVID-19 patients. J. Med. Virol. 2020, 92, 1789–1790. [Google Scholar] [CrossRef]
- De Wit, E.; Van Doremalen, N.; Falzarano, D.; Munster, V.J. SARS and MERS: Recent insights into emerging coronaviruses. Nat. Rev. Microbiol. 2016, 14, 523–534. [Google Scholar] [CrossRef]
- Bianchi, M.; Benvenuto, D.; Giovanetti, M.; Angeletti, S.; Ciccozzi, M.; Pascarella, S. Sars-CoV-2 envelope and membrane proteins: Structural differences linked to virus characteristics? BioMed. Res. Int. 2020, 2020, 1–6. [Google Scholar] [CrossRef]
- Astuti, I.; Ysrafil. Severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2): An overview of viral structure and host response. Diabetes Metab. Syndr. Clin. Res. Rev. 2020, 14, 407–412. [Google Scholar] [CrossRef] [PubMed]
- Synytsya, A.; Novak, M. Structural analysis of glucans. Ann. Transl. Med. 2014, 2, 17. [Google Scholar] [CrossRef]
- Akramienė, D.; Kondrotas, A.; Didžiapetrienė, J.; Kėvelaitis, E. Effects of ß-glucans on the immune system. Medicina 2007, 43, 597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wasser, S.P. Medicinal mushrooms as a source of antitumor and immunomodulating polysaccharides. Appl. Microbiol. Biotechnol. 2002, 60, 258–274. [Google Scholar] [CrossRef]
- Pizarro, C.; Ronco, M.; Maria, A.; Gotteland, R. Beta-glucans: What types exist and what are their health benefits? Rev. Chil. Nutr. 2014, 41, 439–446. [Google Scholar] [CrossRef] [Green Version]
- Ooi, V.E.; Liu, F. Immunomodulation and anti-cancer activity of polysaccharide-protein complexes. Curr. Med. Chem. 2000, 7, 715–729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chan, G.C.-F.; Chan, W.K.; Sze, D.M.-Y. The effects of β-glucan on human immune and cancer cells. J. Hematol. Oncol. 2009, 2, 25. [Google Scholar] [CrossRef] [Green Version]
- Schwartz, B.; Hadar, Y. Possible mechanisms of action of mushroom-derived glucans on inflammatory bowel disease and associated cancer. Ann. Transl. Med. 2014, 2, 19. [Google Scholar] [CrossRef]
- Brown, G.D.; Gordon, S. Fungal β-glucans and mammalian immunity. Immunity 2003, 19, 311–315. [Google Scholar] [CrossRef] [Green Version]
- Hong, F.; Yan, J.; Baran, J.T.; Allendorf, D.J.; Hansen, R.D.; Ostroff, G.R.; Xing, P.X.; Cheung, N.-K.V.; Ross, G.D. Mechanism by which orally administered β-1,3-glucans enhance the tumoricidal activity of antitumor monoclonal antibodies in murine tumor models. J. Immunol. 2004, 173, 797–806. [Google Scholar] [CrossRef] [Green Version]
- Sun, L.; Zhao, Y. The biological role of dectin-1 in immune response. Int. Rev. Immunol. 2007, 26, 349–364. [Google Scholar] [CrossRef] [PubMed]
- Schorey, J.S.; Lawrence, J.S.S.A.C. The pattern recognition receptor dectin-1: From fungi to mycobacteria. Curr. Drug Targets 2008, 9, 123–129. [Google Scholar] [CrossRef]
- Brown, G.D. Dectin-1: A signalling non-TLR pattern-recognition receptor. Nat. Rev. Immunol. 2005, 6, 33–43. [Google Scholar] [CrossRef]
- Dixon, D.; Darveau, R. Lipopolysaccharide heterogeneity: Innate host responses to bacterial modification of lipid a structure. J. Dent. Res. 2005, 84, 584–595. [Google Scholar] [CrossRef] [PubMed]
- Goodridge, H.S.; Simmons, R.M.; Underhill, D. Dectin-1 stimulation by candida albicans yeast or zymosan triggers NFAT activation in macrophages and dendritic cells. J. Immunol. 2007, 178, 3107–3115. [Google Scholar] [CrossRef] [Green Version]
- Gross, O.; Gewies, A.; Finger, K.; Schäfer, M.; Sparwasser, T.; Peschel, C.; Förster, I.; Ruland, J. Card9 controls a non-TLR signalling pathway for innate anti-fungal immunity. Nat. Cell Biol. 2006, 442, 651–656. [Google Scholar] [CrossRef]
- Rogers, N.C.; Slack, E.C.; Edwards, A.D.; Nolte, M.A.; Schulz, O.; Schweighoffer, E.; Williams, D.L.; Gordon, S.; Tybulewicz, V.L.; Brown, G.D.; et al. Syk-dependent cytokine induction by dectin-1 reveals a novel pattern recognition pathway for C type lectins. Immunity 2005, 22, 507–517. [Google Scholar] [CrossRef] [Green Version]
- Gantner, B.N.; Simmons, R.M.; Canavera, S.J.; Akira, S.; Underhill, D.M. Collaborative induction of inflammatory responses by dectin-1 and toll-like receptor 2. J. Exp. Med. 2003, 197, 1107–1117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soltanian, S.; Stuyven, E.; Cox, E.; Sorgeloos, P.; Bossier, P. Beta-glucans as immunostimulant in vertebrates and invertebrates. Crit. Rev. Microbiol. 2009, 35, 109–138. [Google Scholar] [CrossRef] [PubMed]
- Taylor, P.R.; Brown, G.; Herre, J.; Williams, D.L.; Willment, J.; Gordon, S. The Role of SIGNR1 and the β-Glucan receptor (Dectin-1) in the nonopsonic recognition of yeast by specific macrophages. J. Immunol. 2004, 172, 1157–1162. [Google Scholar] [CrossRef] [Green Version]
- Lin, Y.-L.; Liang, Y.-C.; Lee, S.-S.; Chiang, B.-L. Polysaccharide purified from Ganoderma luciduminduced activation and maturation of human monocyte-derived dendritic cells by the NF-κB and p38 mitogen-activated protein kinase pathways. J. Leukoc. Biol. 2005, 78, 533–543. [Google Scholar] [CrossRef] [Green Version]
- Cobb, B.A.; Wang, Q.; Tzianabos, A.O.; Kasper, D.L. Polysaccharide processing and presentation by the MHCII pathway. Cell 2004, 117, 677–687. [Google Scholar] [CrossRef]
- Klein, E.; Di Renzo, L.; Yefenof, E. Contribution of CR3, CD11b/CD 18 to cytolysis by human NK cells. Mol. Immunol. 1990, 27, 1343–1347. [Google Scholar] [CrossRef]
- Di Renzo, L.; Yefenof, E.; Klein, E. The function of human NK cells is enhanced by β-glucan, a ligand of CR3 (CD11b/CD18). Eur. J. Immunol. 1991, 21, 1755–1758. [Google Scholar] [CrossRef] [PubMed]
- Mizuno, M.; Nishitani, Y. Immunomodulating compounds in basidiomycetes. J. Clin. Biochem. Nutr. 2013, 52, 202–207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vannucci, L.; Krizan, J.; Sima, P.; Stakheev, D.; Čaja, F.; Rajsiglová, L.; Horak, V.; Saieh, M. Immunostimulatory properties and antitumor activities of glucans. Int. J. Oncol. 2013, 43, 357–364. [Google Scholar] [CrossRef] [Green Version]
- Xu, X.; Yan, H.; Tang, J.; Chen, J.; Zhang, X. Polysaccharides inLentinus edodes: Isolation, Structure, Immunomodulating Activity and Future Prospective. Crit. Rev. Food Sci. Nutr. 2013, 54, 474–487. [Google Scholar] [CrossRef]
- Jung, K.; Ha, Y.; Ha, S.K.; Han, D.U.; Kim, D.W.; Moon, W.K.; Chae, C. Antiviral effect of saccharomyces cerevisiaebeta-glucan to swine influenza virus by increased production of interferon-gamma and nitric oxide. J. Vet. Med. Ser. B 2004, 51, 72–76. [Google Scholar] [CrossRef]
- Leblanc, B.W.; Albina, J.E.; Reichner, J.S. The effect of PGG-β-glucan on neutrophil chemotaxis in vivo. J. Leukoc. Biol. 2006, 79, 667–675. [Google Scholar] [CrossRef]
- Babineau, T.J.; Marcello, P.; Swails, W.; Kenler, A.; Bistrian, B.; Forse, R.A. Randomized phase I/II trial of a macrophage-specific immunomodulator (PGG-glucan) in high-risk surgical patients. Ann. Surg. 1994, 220, 601–609. [Google Scholar] [CrossRef]
- Dellinger, E.P.; Babineau, T.J.; Bleicher, P.; Kaiser, A.B.; Seibert, G.B.; Postier, R.G.; Vogel, S.B.; Norman, J.; Kaufman, D.; Galandiuk, S.; et al. Effect of PGG-glucan on the rate of serious postoperative infection or death observed after high-risk gastrointestinal operations. Arch. Surg. 1999, 134, 977–983. [Google Scholar] [CrossRef] [Green Version]
- Murphy, E.J.; Masterson, C.; Rezoagli, E.; O’Toole, D.; Major, I.; Stack, G.D.; Lynch, M.; Laffey, J.G.; Rowan, N.J. β-Glucan extracts from the same edible shiitake mushroom Lentinus edodes produce differential in-vitro immunomodulatory and pulmonary cytoprotective effects—Implications for coronavirus disease (COVID-19) immunotherapies. Sci. Total. Environ. 2020, 732, 139330. [Google Scholar] [CrossRef] [PubMed]
- Tzianabos, A.O. Polysaccharide immunomodulators as therapeutic agents: Structural aspects and biologic function. Clin. Microbiol. Rev. 2000, 13, 523–533. [Google Scholar] [CrossRef]
- Schepetkin, I.A.; Quinn, M.T. Botanical polysaccharides: Macrophage immunomodulation and therapeutic potential. Int. Immunopharmacol. 2006, 6, 317–333. [Google Scholar] [CrossRef]
- Vetvicka, V.; Yvin, J.-C. Effects of marine β−1,3 glucan on immune reactions. Int. Immunopharmacol. 2004, 4, 721–730. [Google Scholar] [CrossRef]
- Soares, E.; Cordeiro, R.; Faneca, H.; Borges, O. Polymeric nanoengineered HBsAg DNA vaccine designed in combination with β-glucan. Int. J. Biol. Macromol. 2019, 122, 930–939. [Google Scholar] [CrossRef]
- Cavaleri, M.; Enzmann, H.; Straus, S.; Cooke, E. The European medicines agency’s EU conditional marketing authorisations for COVID-19 vaccines. Lancet 2021, 397, 355–357. [Google Scholar] [CrossRef]
- Krause, P.R.; Gruber, M.F. Emergency use authorization of Covid vaccines—safety and efficacy follow-up considerations. N. Engl. J. Med. 2020, 383, e107. [Google Scholar] [CrossRef] [PubMed]
- Ye, T.; Zhong, Z.; García-Sastre, A.; Schotsaert, M.; De Geest, B.G. Current status of COVID-19 (Pre)clinical vaccine development. Angew. Chem. Int. Ed. 2020, 59, 18885–18897. [Google Scholar] [CrossRef] [PubMed]
- Di Luzio, N.R.; Williams, D.L. Protective effect of glucan against systemic Staphylococcus aureus septicemia in normal and leukemic mice. Infect. Immun. 1978, 20, 804–810. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bistoni, F.; Verducci, G.; Perito, S.; Vecchiarelli, A.; Puccetti, P.; Marconi, P.; Cassone, A. Immunomodulation by a low-virulence, agerminative variant of Candida albicans. Further evidence for macrophage activation as one of the effector mechanisms of nonspecific anti-infectious protection. J. Med. Vet. Mycol. 1988, 26, 285–299. [Google Scholar] [CrossRef] [PubMed]
- Aaby, P.; Samb, B.; Simondon, F.; Seck, A.M.C.; Knudsen, K.; Whittle, H. Non-specific beneficial effect of measles immunisation: Analysis of mortality studies from developing countries. BMJ 1995, 311, 481–485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakurai, T.; Hashimoto, K.; Suzuki, I.; Ohno, N.; Oikawa, S.; Masuda, A.; Yadomae, T. Enhancement of murine alveolar macrophage functions by orally administered β-glucan. Int. J. Immunopharmacol. 1992, 14, 821–830. [Google Scholar] [CrossRef]
- Xiao, Z.; Trincado, C.A.; Murtaugh, M.P. Beta-glucan enhancement of T cell IFNupsilon response in swine. Vet. Immunol. Immunopathol. 2004, 102, 315–320. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, J.A.; Kastello, M.D.; Harrington, D.G.; Crabbs, C.L.; Peters, C.J.; Jemski, J.V.; Scott, G.H.; Di Luzio, N.R. Glucan-induced enhancement of host resistance to selected infectious diseases. Infect. Immun. 1980, 30, 51–57. [Google Scholar] [CrossRef] [Green Version]
- Van der Meer, J.W.; Joosten, L.A.; Riksen, N.; Netea, M.G. Trained immunity: A smart way to enhance innate immune defence. Mol. Immunol. 2015, 68, 40–44. [Google Scholar] [CrossRef]
- Keating, S.T.; Groh, L.; van der Heijden, C.D.; Rodriguez, H.; dos Santos, J.C.; Fanucchi, S.; Okabe, J.; Kaipananickal, H.; van Puffelen, J.; Helder, L.; et al. The Set7 lysine methyltransferase regulates plasticity in oxidative phosphorylation necessary for trained immunity induced by β-glucan. Cell Rep. 2020, 31, 107548. [Google Scholar] [CrossRef]
- Saeed, S.; Quintin, J.; Kerstens, H.H.D.; Rao, N.A.; Aghajanirefah, A.; Matarese, F.; Cheng, S.-C.; Ratter, J.; Berentsen, K.; van der Ent, M.A.; et al. Epigenetic programming of monocyte-to-macrophage differentiation and trained innate immunity. Science 2014, 345, 1251086. [Google Scholar] [CrossRef] [Green Version]
- Yun, C.-H.; Estrada, A.; Van Kessel, A.; Park, B.-C.; Laarveld, B. β-Glucan, extracted from oat, enhances disease resistance against bacterial and parasitic infections. FEMS Immunol. Med. Microbiol. 2003, 35, 67–75. [Google Scholar] [CrossRef] [Green Version]
- Dos Santos, J.C.; de Figueiredo, A.M.B.; Silva, M.V.T.; Cirovic, B.; de Bree, L.C.J.; Damen, M.S.; Moorlag, S.J.; Gomes, R.S.; Helsen, M.M.; Oosting, M.; et al. β-Glucan-induced trained immunity protects against leishmania braziliensis infection: A crucial role for IL-32. Cell Rep. 2019, 28, 2659–2672. [Google Scholar] [CrossRef] [Green Version]
- Jesenak, M.; Majtan, J.; Rennerova, Z.; Kyselovic, J.; Banovcin, P.; Hrubisko, M. Immunomodulatory effect of pleuran (β-glucan from Pleurotus ostreatus) in children with recurrent respiratory tract infections. Int. Immunopharmacol. 2013, 15, 395–399. [Google Scholar] [CrossRef]
- Adams, D.S.; Pero, S.C.; Petro, J.B.; Nathans, R.; Mackin, W.M.; Wakshull, E. PGG-Glucan activates NF-κB-like and NF-IL-6-like transcription factor complexes in a murine monocytic cell line. J. Leukoc. Biol. 1997, 62, 865–873. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neyrinck, A.; Mouson, A.; Delzenne, N. Dietary supplementation with laminarin, a fermentable marine β (1–3) glucan, protects against hepatotoxicity induced by LPS in rat by modulating immune response in the hepatic tissue. Int. Immunopharmacol. 2007, 7, 1497–1506. [Google Scholar] [CrossRef] [PubMed]
- Talbott, S.M.; Talbott, J.A. Baker’s yeast beta-glucan supplement reduces upper respiratory symptoms and improves mood state in stressed women. J. Am. Coll. Nutr. 2012, 31, 295–300. [Google Scholar] [CrossRef]
- Nicholls, E.F.; Madera, L.; Hancock, R. Immunomodulators as adjuvants for vaccines and antimicrobial therapy. Ann. N. Y. Acad. Sci. 2010, 1213, 46–61. [Google Scholar] [CrossRef] [PubMed]
- Le, T.H.; Le, T.B.; Doan, T.H.T.; Van Quyen, D.; Le, K.X.T.; Pham, V.C.; Nagataki, M.; Nomura, H.; Ikeue, Y.; Watanabe, Y.; et al. The adjuvant effect of sophy β-glucan to the antibody response in poultry immunized by the avian influenza a H5N1 and H5N2 vaccines. J. Microbiol. Biotechnol. 2011, 21, 405–411. [Google Scholar] [CrossRef]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Córdova-Martínez, A.; Caballero-García, A.; Roche, E.; Noriega, D.C. β-Glucans Could Be Adjuvants for SARS-CoV-2 Virus Vaccines (COVID-19). Int. J. Environ. Res. Public Health 2021, 18, 12636. https://doi.org/10.3390/ijerph182312636
Córdova-Martínez A, Caballero-García A, Roche E, Noriega DC. β-Glucans Could Be Adjuvants for SARS-CoV-2 Virus Vaccines (COVID-19). International Journal of Environmental Research and Public Health. 2021; 18(23):12636. https://doi.org/10.3390/ijerph182312636
Chicago/Turabian StyleCórdova-Martínez, Alfredo, Alberto Caballero-García, Enrique Roche, and David C. Noriega. 2021. "β-Glucans Could Be Adjuvants for SARS-CoV-2 Virus Vaccines (COVID-19)" International Journal of Environmental Research and Public Health 18, no. 23: 12636. https://doi.org/10.3390/ijerph182312636