Abstract
Biofilms in burns are major problems: bacterial communities rapidly develop antibiotic resistance, and 60% of burn mortality is attributed to biofilms. Key pathogens are Pseudomonas aeruginosa, methicillin-resistant Staphylococcus aureus, and multidrug-resistant Acinetobacter baumanii. Purpose: identify current and novel interventions to reduce biofilms on patients’ burns and hospital surfaces and equipment. Medline and Embase were searched without date or language limits, and 31 possible interventions were prioritised: phages, nano-silver, AgSD-NLs@Cur, Acticoat and Mepilex silver, acetic acid, graphene-metal combinations, CuCo2SO4 nanoparticles, Chlorhexidene acetate nanoemulsion, a hydrogel with moxifloxacin, carbomer, Chitosan and Boswellia, LED light therapy with nano-emodin or antimicrobial blue light + Carvacrol to release reactive oxygen species, mannosidase + trypsin, NCK-10 (a napthalene compound with a decyl chain), antimicrobial peptide PV3 (includes two snake venoms), and polypeptides P03 and PL2. Most interventions aimed to penetrate cell membranes and reported significant reductions in biofilms in cfu/mL or biofilm mass or antibiotic minimal inhibitory concentrations or bacterial expression of virulence or quorum sensing genes. Scanning electron microscopy identified important changes in bacterial surfaces. Patients with biofilms need isolating and treating before full admission to hospital. Cleaning and disinfecting needs to include identifying biofilms on keyboards, tablets, cell phones, medical equipment (especially endoscopes), sinks, drains, and kitchens.
1. Introduction
Biofilms affect > 80% of bacterial infections in humans [1,2] and are the dominant mode of bacterial growth in which millions of bacteria cohabit in a hydrated extracellular matrix [1]. Biofilms are a key problem in burns, and 60% of the mortality from burns is attributed to biofilms [1]. Biofilms are also a key problem in chronic wounds such as diabetic, pressure and venous leg ulcers, lung infections in cystic fibrosis, pneumonia in patients on ventilators, and patients on medical devices and urinary catheters [1]. Most hospital-acquired infections are due to vancomycin-resistant enterococcus (VRE), methicillin-resistant Staphylococcus aureus, Klebsiella pneumoniae, multidrug-resistant Acinetobacter baumanii, Pseudomonas aeruginosa, and extended spectrum beta-lactamase-producing organisms (ESBL) [3].
Biofilms have five life stages: reversible attachment to surfaces, irreversible attachment, maturation-1, maturation-2, and the planktonic form. During the final stage, ~80% of the biomass may convert back to the planktonic form and bacterial susceptibility to antibiotics becomes similar to that of other planktonic cells [1]. Factors related to biomass dispersion are decreased availability of carbon, iron, oxygen, pyruvate and nitric oxide, oxidative stress, and starvation and decreased quorum-sensing molecules such as farnesol, indole and N-acylhomoserine lactone, and increases in bile salts and cis-2-decenomic acid [1].
Bacteria in biofilms can develop resistance 100- to 1000-fold against multiple antimicrobials compared to planktonic cells [4]. Gram-negative bacteria are a key problem in biofilms because they have an outer and an inner cell membrane. Penicillin class antibiotics are degraded by enzymes within bacterial cells [5], and the polar lipopolysaccharides in the outer membrane are hydrophobic to antibiotics such as rifampicin. In the membranes, impaired function of influx pumps and increased function of efflux pumps enable Gram-negatives to acquire resistance against multiple antibiotics because of the resulting sub-minimal inhibitory concentration (MIC) of antibiotics within cells. One group of efflux pumps (resistance-modulation class) effluxes antibiotics using energy from the proton motive force generated by differences in potential between ions and protons across membrane walls and interventions in cell membranes to alter these potential differences could be therapeutic [5]. Changes in membrane porins and decreased passive diffusion of antibiotics also decrease antibiotic concentrations.
Increased resistance to antibiotics is also caused by transfers of resistance genes between bacteria, slow bacterial growth rates within biofilms, persister cells, which are particularly resistant to destruction, and quorum-sensing gene activity between bacteria [4]. Bacteria within biofilms minimise the host immune system activities by using quorum sensing genes to communicate. In P. aeruginosa, the LasI gene affects biofilm formation through the quorum sensing system and also regulates the virulence factors alkaline phosphatase, elastase, exotoxin A, pyocyanin, and rhamnolipid [2]. The RhlR gene uses butyryl acyl homoserine lactone to regulate its own gene and the quorum-sensing operon and the genes coding for pyocyanin, siderophores, and rhamnolipid synthesis enzymes [6]. In A. baumanii, the abal gene affects the quorum sensing system and biofilm formation through acyl homoserine lactose. In S. aureus, the Agr accessory gene regulator affects the quorum sensing mechanism and biofilm formation [2].
Burns are rapidly colonised by Gram-positive bacteria, principally S. aureus, from the patients’ skin and infected environmental surfaces that patients contact and then within hours to a few days the wound is colonised by Gram-negative bacteria, principally P. aeruginosa and A. baumanii. A study of burns in ICUs and burn wards identified 1621 pathogens in 2395 clinical samples, of which 74.2% were Gram-negative and 34.3% were A. baumanii [7]. The early treatment of burns is important to prevent colonisation by multiple bacteria, particularly P. aeruginosa and A. baumanii.
Thus the initial step in burn therapy is wound cleaning and debridement to remove necrotic tissue, which significantly reduces blood flow and immune system access. Topical dressings and antibiotics and then IV antibiotics are applied if the patient does not respond but high resistance levels to multiple antibiotics soon develop. The consequences for patients are failure of skin grafts [8], bacteraemia, infections of multiple organs, and mortality. Multiple therapies have been developed to be applied directly to burns to inhibit bacteria in both the planktonic phase and prevent biofilm formation by Gram-positives and Gram-negatives including interventions to damage bacterial cell walls, especially Gram-negative inner and outer membranes, and facilitate higher entry levels of antibiotics. More than 1000 wound models to test these therapies have been reported with 74% in vivo, 23% in vitro, and 3% ex vivo (ex vivo uses samples from living animals but conducts experiments in laboratory equipment). Pigs are the preferred animal as their skin and immune system are closer to humans than rat or mouse skin [9].
A systematic review of interventions in hospitals and long-term care facilities identified 14 cluster-randomised controlled trials of cleaning and disinfecting strategies to reduce the incidence of healthcare-associated infections (HAIs) and multiply drug-resistant organisms (MDROs) of particular concern: methicillin-resistant Staphylococcus aureus (MRSA), vancomycin-resistant enterococcus (VRE), multidrug-resistant Acinetobacter species, and extended spectrum beta-lactamase producing organisms (ESBL). Of fourteen c-RCTs, ten were focused to reduce patient infections by four MDROs and/or HAIs. In four c-RCTs, patient MDRO and/or HAI rates were significantly reduced with cleaning and disinfection strategies, including bleach, quaternary ammonium detergents, ultraviolet irradiation, hydrogen peroxide vapour, and copper-treated surfaces or fabrics, but in six there were no significant changes. Three c-RCTs focused on reducing MRSA rates (one had significant results), and one on Clostridioides difficile had no significant results. No study assessed the contributions of biofilms [10]. The disappointing results of these c-RCTs of disinfection suggest that a key preventive strategy is to first detect patients with MDROs or HAIs, isolate them in an isolation unit, and treat them before they enter other areas of medical facilities to transmit infection to patients, staff, and surfaces. Another key strategy is identifying and destroying biofilms in patients’ burns and on surfaces and medical equipment within medical facilities before patients and staff can contact them.
Patients with burn wounds quickly acquire a range of pathogens from their own skin and their environment and if they are hospitalised for care they are very likely to acquire MDROs and HAIs from surfaces, patients, staff, and equipment in the hospital environment. Burn patients have high rates of biofilm infections, and current research on interventions to reduce biofilms in burn patients includes silver and other metals, disinfectants, hydrogels, light and sound therapy to activate sensitiser molecules to release active oxygen species, a variety of small molecules to enable better penetration of cell wall membranes, glycans, lactobacilli, and phage therapy.
A systematic review and meta-analysis of tests of antimicrobial efficacy against biofilms identified five key model parameters that influence outcomes: the biofilm surface area/volume ratio, biofilm areal cell density, fluid static or flow conditions over the biofilm, biofilm age, and the antibiotic chosen as the comparator. The review compared several pairs of studies and concluded that the experimental method used is the most important factor determining the outcome and presented comparisons of studies that showed that the method chosen can “produce extremely different results even for the same microbial agent.” [11].
A systematic review of topical agents for managing chronic biofilm infections identified 39 in vitro, 5 animal and 3 human in vivo studies involving 44 commercially available topical agents and 78 biofilm-forming bacteria and concluded:
“The analysis clearly identifies a large disparity in the translation of laboratory studies to researchers undertaking human trials.” “When analysing the thirty-nine included in vitro studies, a standardised methodological approach to biofilm testing was not observed. Sixteen different biofilm models were used with significant variations between test parameters such as: choices of different bacterial strain or isolate (n = 78), biofilm growth time (24 h to 168 h), starting log densities, agent exposure duration (3 s to 168 h), and adaptation to in vitro models to more closely resemble a wound environment (n = 22 of 39, 56%).” [12], pp. 266–267).
Purpose: To assess if any current or under development therapies intended to be used directly on patient burns have been tested in well-designed trials and have significant outcomes so that they merit large c-RCTs to test benefits for patient outcomes and also current cleaning and disinfection strategies of surfaces and medical equipment in hospitals and medical facilities to improve outcomes for burn patients.
2. Materials and Methods
Literature searches were conducted on 1 December 2021 in Medline, Embase, Cochrane Central, and Web of Science from inception with no language or date limits using the search terms (biofilm) and (hospital or long-term care facility or medical facility) and (burns). Separate searches were undertaken for (1) (medical equipment or exp stethoscopes or keyboard.mp or exp computer or exp telephone or exp Cell Phones or computer tablets.mp or exp computers Handheld or endoscopes) and (biofilm) and (detection or measurement); and (2) (burns) or (hospitals or long-term care facilities or medical facilities) and (systematic reviews or meta-analyses). Titles and abstracts were assessed and data abstracted independently by two researchers with disagreements resolved by discussion. The systematic review is registered with Prospero and follows the PRISMA reporting requirements
3. Results
3.1. Literature Search
The literature search identified 31 studies of novel interventions to prevent biofilm formation (Figure 1).
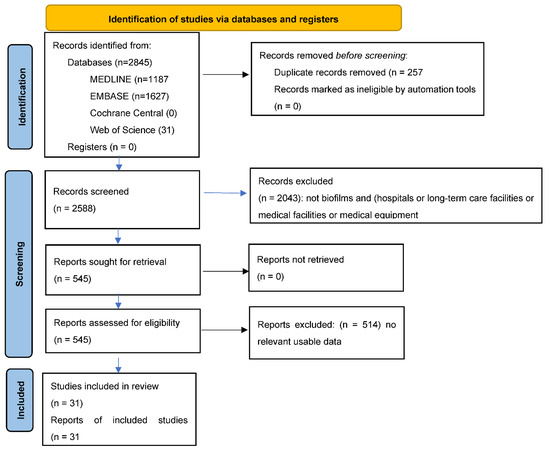
Figure 1.
PRISMA Flow diagram for new systematic reviews which include searches of databases and registers only.
The studies chosen for analysis are those with the most direct relevance to burn care: in vivo human biofilm studies then in vivo studies of pigs (their skin and immune system are closer to human skin than other animals) and then in vivo studies of rats and mice. In vitro studies of pathogens from burn wounds grown in 96-well microplates do not provide as direct evidence because they are not in living mammals and are on abiotic surfaces. All the included studies were from Medline and Embase.
3.2. Interventions
3.2.1. Silver
Silver sulfadiazine has been the principal burn topical therapy for decades. Silver in burns absorbs wound exudates and kills organisms drawn into the dressings; binds to negatively charged proteins, RNA and DNA; damages bacterial cells walls; inhibits replication; and reduces metabolism and growth [13]. With the development of antibiotic resistance including plasmid-mediated resistance, new therapies have been developed which enhance the entry of silver into burn wounds. Gholamrezazadeh found that a nanomolecule formulation of silver (nano-Ag) at 12.5 ng/mL caused a reduction in the number of P. aeruginosa bacteria forming biofilms from 28.5% to 3.5%, whereas benzalkonium MIC at 0.03 mg/mL reduced the number of bacteria forming biofilms from 28.5% to 18.7% [6].
Pourhajibagher sensitised liposomes with the photosensitising agent Cucurmin to create the reactive oxygen species (ROS) superoxide anion O*2¯, hydroxyl radical *OH and singlet oxygen 102 which decrease the expression of bacterial virulence genes. Cucurmin also enables repair and regeneration of damaged eukaryotic cells. In in vitro biofilms with MIC90 doses of silver sulfadiazine nanoliposomes with Curcumin (AgSD-NLs@Cur) activated by light diodes, the cell numbers of A. baumanii in biofilms decreased by 76.4%, with silver sulfadiazine nanoliposomes (AgSD-NKs) by 44.8%, and with silver sulfadiazine (AgSD) by 38.1%. In vivo mouse burn wounds treated with AgSD-NLs@Cur on staining and with light microscopy showed focal epidermis regeneration, fibrosis, and granulation tissue formation, but the controls showed complete loss of the epidermis and hair follicles, hyperaemic vessels, and extensive bacterial colonisation [14].
A comparison of eleven antimicrobial burn dressings found that 72 h after infection with A. baumanii, two silver formulations reduced A. baumanii numbers in biofilms (commercial silver formulations Acticoat by 96%, Mepilex Ag by 95.9%) and acetic acid by 90–93%. P. aeruginosa numbers were reduced by Acticoat by 100%, Mepilex Ag by 100% and acetic acid by 86–93% [15].
3.2.2. Other Metals
Karaky’s in vitro study found that eight metal-graphene combinations reduced P. aeruginosa biofilms by ≥90% (platinum-graphene oxide, gallium-graphene oxide, molybdenum-graphene oxide, gold-graphene oxide, silver-graphene, gallium-graphene, and molybdenum-graphene). The greatest reduction of biofilm metabolic activity occurred with gold-graphene oxide (94%), molybdenum-graphene oxide (93%), silver (91%), and silver-graphene (91%) [8]. Li assessed the effect of H2O2 (2 nM) and CuCo2S4 nanoparticles (100 μg/mL) on mouse burns, and after two days of treatment there was no inflammatory response. The burn wounds contracted whereas the control group showed a severe inflammatory response with suppuration. After six days the group treated with H2O2 at 2 nM and CuCo2S4 at 100 μg/mL showed enhanced healing and 83.7% wound closure, with CuCo2S4 71%, with H2O2 63.3%, and the control 59%. At two weeks, the H2O2, and CuCo2S4- treated wounds were completely closed and healed [15].
Nozari compared chitosan/alginate + ZnO nanoparticles to chitosan/bentonite + ZnO nanoparticles and noted ~1 × 104 lower cfu/mL rates of S. aureus and P. aeruginosa compared to control (99.99% reductions). With in vivo mouse burns at 7 days for the treated rats there were re-epithelialisation, active fibroblasts, and hair follicles, and sebaceous glands were detected but there was no re-epithelialisation in untreated rats [16].
3.2.3. Disinfectants
Halstead assessed the effect of acetic acid at concentrations ranging from 0.31% to 5% against 23 isolates of 6 MDROs and in vivo biofilms; the acetic acid MBIC was 0.31%, the MBEC against formed biofilms ranged from ≤0.10% to 2.5% and eradication of mature biofilms was observed for all isolates after three hours of exposure [17].
Song treated mouse burn wounds with Chlorhexidene acetate nanoemulsion (CNE) 2 μg/mL, and according to scanning electron microscopy MRSA biofilms were “dispersed and disrupted and obvious reduction in number of bacteria,” there were large vacuoles between the cell wall and cytoplasm. There was also leakage of DNA, proteins, K+, and Mg2+. The dead/live cell ratio with CNE was 83.6% [18].
Tiwari compared the effects of sodium hypochlorite and ethanol on S. aureus in vitro biofilms and found no significant differences in reductions of strong compared to weak S. aureus biofilm formers. However, electron microscopy of strong biofilm producers showed significant depressions and irregular craters on their surface [19].
3.2.4. Hydrogels
Andersson’s study of Göttingen minipig burn biofilms compared Prontosan (0.1% polyhexamethylene biguanide and 0.1% undecylenamidopropyl betaine) or levofloxacin on S. aureus and P. aeruginosa and on scanning electron microscopy both antibacterial treatments “visibly reduced” the number of bacterial cells on the wound surfaces and that perturbations and bacterial clumping and debris were noted on the Prontosan-treated biofilms. The bioluminescence levels of luminescent S. aureus and P. aeruginosa at two hours after antibiotic therapy were significantly reduced by Protosan (p < 0.001 to 0.0001) [9].
Chhibber treated MRSA burn wounds with a hydrogel (moxifloxacin 0.5% w/v, carbomer 1% w/v, Chitosan 5 mg/mL, Boswellia gum 0.5%) and showed a 3.5 log10 reduction on day 1 and a 4.8 log10 reduction on day 2, whereas the control increased to 6.9 log10 on day 3. At 4 h, there was complete eradication of MRSA from the wounds but MRSA was established in the control mice. At 24 h there was subdued inflammation and signs of healing in the treated mice but in the control mice there was loss of epithelium, proliferation of neutrophils, and a thick layer of inflammatory cells [20].
3.2.5. Sound
Pourhajibagher used ultrasound on nanoemodin (2.5 × 10−4 g/L) to release oxygen species (ROS) including superoxide anion O*2¯, hydroxyl radical *OH, and singlet oxygen 1O2, which damaged bacterial cell membranes, proteins, and DNA. There was a reduction in Staphylococcus aureus, Pseudomonas aeruginosa, and Acinetobacter baumannii following sonotherapy at 1/2 MIC of N-EMO of 81.5%; at 1/16 MBIC 71.0%; and at 1/128 MBEC 57.8 with reductions in log10 cfu/mL of 99.99%, 99.97%, and 99.48% but no effect with ultrasound alone [21].
3.2.6. Light
Ishiwata compared ethylene diamine-tetra-acetic acid disodium salt (EDTA), which suppresses biofilms, and dimethyl sulfoxide (DMSO), which is a tissue-penetration enhancer on rat burns. Only 2/14 of the rats survived to 7 days, but 11/14 of those also exposed to methylene blue 665 nm LED diodes at 45 mW/cm2 at 2.5 cms three times daily for 20 min × 7 days, presumably because the recurrent reductions in the P. aeruginosa levels gave the LED-treated rats a better chance [22].
Lu used the phytochemical carvacrol at 0.2 mg/mL and blue light 450 nm at 75 J/cm2 to excite porphyrin-like derivatives in bacterial cells to produce reactive oxygen species (ROS). In vitro biofilms of Acinetobacter baumanni, Pseudomonas aeruginosa, and MRSA at 107 CFU were completely eliminated (p < 0.0001). The thickness of Acinetobacter baumanni biofilm was reduced from 58.6 μm to 1.4 μm and MRSA biofilm from 32.4 μm to 1.7 μm. In in vivo mouse burns infected with Acinetobacter baumainii at 5 × 105 cfu/mL, carvacrol 50 μL at 1 mg/mL and blue light for 12 min (40 J/cm2) eliminated log 8 of luminescence in luminescent bacteria, blue light alone 2.3 log, and carvacrol 0.8 log. Reactive oxygen species increased 14-fold in the Acinetobacter baumanni group, 12-fold in the Pseudomonas aeruginosa group, and 8-fold in the MRSA group [3].
Pourhajibagher exposed Acinetobacter baumannii, Pseudomonas aeruginosa, and Staphylococcus aureus to photodynamic therapy (aPDT) with the photosensitiser indocyanine green (ICG) at 1000 μg/mL and a diode laser at 810 nm. There was a significant reduction in cell viability of A. baumannii to 1.5 × 105 cfu/mL, P. aeruginosa to 1 × 105 cfu/mL and S. aureus to 1.0 × 105 cfu/mL compared to control at 4.5 × 105 cfu/mL (all p < 0.05). There was also a 54% increase in reactive oxygen species (ROS) compared to controls and decreases in the expression of the P. aeruginosa quorum-sensing abal gene by 1.9-fold, agrA by 3.7-fold and lasI by 4.9-fold. As shown by scanning electron microscopy with a diode laser + ICG there was a reduction in cell size and numbers, cell elongation, and increased cell destruction but no change with the diode laser or ICG individually [2].
Wang exposed 72 h old A. baumanii biofilms to antimicrobial blue light (aBL) 432 J/cm2 for 72 min, which resulted in the inactivation of 3.18 log10 cfu/mL and the exposure of P. aeruginosa biofilms to aBL in the inactivation of 3.12 log10 cfu/mL for 72 h old biofilms, but control biofilms showed only a <0.27 log10 cfu/mL loss of viability for A. baumanii and <0.42 log10 cfu/mL for P. aeruginosa. An in vivo study of mouse burn wound biofilms infected with 5 × 106 cfu/mL A. baumanii at 24 h required 360 J/cm2 at 48 h 540 J/cm2 to inactivate 3 log10 cfu/mL [4].
3.3. Small Molecules
Mannosidase and trypsin enzymes Mannosidase and trypsin enzymes attack the biofilm matrix of P. aeruginosa, which consists of three layers: alginate (a polymer of β-d-mannuronic acid and α-l-glucuronic acid which provides structural stability and protection of the biofilm); a repeating polysaccharide of D-mannose, D-glucose, and L rhamnose with an important role in biofilm formation and protection; and a glucose-rich layer (Pel). Banar compared ceftazidime (CAZ) 1024 μg/mL, CAZ + α-mannosidase 4 μg/mL, CAZ + β-mannosidase 4–8 μg/mL, and CAZ + trypsin 8–32 μg/mL, and all combinations killed bacterial biofilm cells at these concentrations [23].
Antimicrobial compounds with an aromatic naphalene (N) or benzene (B) core, a L-lysine moiety and a variable lipophilic chain Ghosh found that a compound with a naphthalene core and a decyl chain appendage (NCK-10) was the most active against NDM-1-producing Gram-negative pathogens. For in vitro biofilms of A. baumanii, E. coli, K. pneumoniae, and P. aeruginosa, the MIC for NCK-10 was 4.5 μg/mL. NCK-10 completely lysed persister cells of 5 log cfu/mL E. coli after 2 h but colonies persisted in the control group at 5 log cfu/mL. To disrupt biofilms, the EC50 was 30 μM against biofilms of A. baumanii (MTCC 1425), 20 μM against E. coli (MTCC 443), 26 μM against K. pneumoniae (ATCC 700603), and 19 μM against P. aeruginosa (MTCC 424). On confocal microscopy in the treated samples the biofilms were completely disrupted. NTK-10 did not induce bacterial resistance (there was no change in MIC after 20 passages) but the MIC of colistin increased 250-fold. In burn wounds of mice there was significant reduction in bacterial burden after daily topical treatments with 40 mg/kg × 7 days compared to control [24].
Pyruvate-dehydrogenase PDH catalyses pyruvate to acetyl-CoA in the presence of CoA and NAD+ and the microcolony formation factor MifR. Goodwine assessed if enzyme pyruvate-dehydrogenase (PDH) would increase the efficacy of tobramycin killing of biofilms of P. aeruginosa and S. aureus strains from wound debridement samples. In in vitro human wound samples, there was a 2.2-fold reduction in bacteria after exposure to 5 mU DPH and 2.9-fold reduction after 10–20 mU. In in vitro biofilms investigated with confocal laser scanning microscopy, 60% of microcolonies in PDH-treated biofilms showed signs of dispersion with central voids, but only 8% of untreated biofilms [1].
Li-F type peptide AMP-jsa9 Han assessed whether the Li-F type peptide AMP-jsa9 (which both kills planktonic cells and penetrates MRSA cell membranes) would reduce S. aureus biofilms. Cell viability was reduced to 10% with 8 × MIC vancomycin (8 μg/mL) and to 10% with AMP-jsa9 at 8 × MIC (128 μg/mL). Biomass was reduced to 15% with 8 × MIC Vancomycin (8 μg/mL) and to 15% with AMP-jsa9 at 8 × MIC (128 μg/mL). The viable cell counts in mouse skin burns treated with vancomycin or AMP-jsa9 were 101 to 102 on days 3 and 7 and at 3 days compared to those treated with kanamycin 2–3 × 104 or saline 5–6 × 105 [25].
D-LANA-14. D-lysine conjugated aliphatic norspermidine analogue with a tetradecanoyl chain, which can depolarise Gram-negative cell membranes. Konai found D-LANA-14 was “moderately active” at MICs 32–64 μg/mL against three strains of A. baumanii and four strains of P. aeruginosa, but the combination of D-LANA-14 at sub-MIC levels enabled tetracycline at 4 μg/mL and rifampicin at 2 μg/mL to be active against both bacteria. Against in vitro biofilms, confocal scanning electron microscopy showed that D-LANA-14 (8 μg/mL) plus colistin (8 μg/mL) resulted in >80% reduction in the biofilm mass of A. baumanii-R674 and P. aeruginosa-R590, but D-LANA-14 (8 μg/mL) alone showed no effect, and rifampicin (8 μg/mL) 25–30% disruption. In burn wounds in mice D-LANA-14 (40 mg/kg) plus rifampicin (40 mg/kg) caused a 4.9 log reduction in A. baumanii-R674 and a 4.0 log reduction in P. aeruginosa-R5902, but for D-LANA-14 alone it was 2.3 log and 1.3 log, and for rifampicin alone it was 3.0 log and 1.6 log [5].
PV3 antimicrobial peptides can disrupt negatively charged bacterial cell membranes. PV3 includes terminal residues from two snakes: pEM-2 from Bothrops asper and Mastopparan-VT-1 from Vespa tropica. For multi-drug-resistant strains of P. aeruginosa from burn wounds of hospitalised patients in Iran, the MIC and MBC of PV3 were 2–4 μg/mL, and for ceftazidime, the MIC was 16–256 μg/mL. For the in vitro biofilms, PV3 at 8 × MIC at 24 h resulted in “almost” 100% killing of cells and 95% biomass removal and scanning electron microscopy of the results showed that the PV3-treated cells were shorter, with blisters on membranes, roughness, and blebbing [26].
Poly(l-ornithine)s and poly(l-lysines) Pan assessed the ability of amino-acid-based star-shaped poly(l-ornithine)s and poly(l-lysines) with varying surface charge/hydrophobicity balances (P03, PL2, PH3) to disrupt bacterial cells and biofilms. In vitro P03 reduced the biomass of P. aeruginosa biofilms by 76.9%, PL2 by 35.1%, PH2 by 31.45%, and Polymixin by 7.8%. In mouse burn wounds, P03 caused a 78.2% reduction in P. aeruginosa and PL2 caused a 49.3% reduction compared to Polymixin B [27].
Platensimycin (PTM) and Platensimycin-thioether analogues (PTM-2t) target the Fab/FabF of bacterial fatty acid synthases. For in vitro biofilms of S. aureus, ATCC 291213 after treatment with 2 μg/mL of PTM or PTM-2t the biofilm was reduced by 95%. For the mouse burns treated with 4 mg of PTM or PTM-2t cream on the wound twice daily × 7 days, PTM reduced S. aureus to 2 × 106 cfu/g and PTM-2t to 8.6 × 106 cfu/g compared to 2.5 × 106 cfu/g for mupirocin and untreated mice 4.3 × 108 cfu/g. On haematoxylin and eosin staining, untreated mice showed partially destroyed hair follicles, an incomplete fat layer, and a large number of inflammatory cells in the muscle layer, but treated mice had “relatively healed skin structure” [28].
Salicylidene acylhydrazide INP0341 was assessed for its ability to inhibit the P. aeruginosa type III secretion system (T3SS) which translocates virulence factors; the four exoenxyme (Exo) molecules S, T, U, and Y from the bacterial cytosol directly into the host cytoplasm where they alter cell function to permit bacterial growth; and the flagellum system required for biofilm formation and motility. In P. aeruginosa, flagella are required for swimming, and flagellate and type IV pili for swarming. In vitro INP0341 significantly reduced in a dose-dependent manner the expression and secretion of the Type III secretion system T3SS ExoS required by P. aeruginosa for colonisation and survival in host cells. INP0341 disarmed but did not kill P. aeruginosa cells. In in vitro biofilms, INP0321 at 100 μM reduced biofilm mass to 40% of control (p < 0.05), inhibited P. aeruginosa swarming, and prevented movement across semisolid surfaces, which requires flagella and type IV pili. In vivo, the treated mice died at 36 h and controls at 42 h (p < 0.05) [29].
3.4. Glycans
Mucus lines all wet epithelial cells in the human body, including lungs, gastrointestinal and urogenital tracts, and eyes. It is the body’s first line of defence against pathogens and is occupied by trillions of sensing bacteria and white cells. Mucins suppress virulence genes, encourage the planktonic state in bacteria, prevent attachment to surfaces, and reduce bacterial toxicity to human cells. Mucins have many complex glycan structures covalently linked to serine and threonine and have been assessed as potential methods of decreasing biofilm formation. Wheeler exposed P. aeruginosa PA01 biofilms to MUC5AC glycans from fresh pig intestines, MUC2 from pig stomachs, and MUCB human salivary glycans. P. aeruginosa PA01 biofilms were exposed to these mucins, and 70% of the cells dissociated from surfaces into the planktonic phase (p < 0.0001). Intestinal mucins suppressed quorum sensing (lasR), siderophore biosynthesis (pvdA), and type-three secretion (pcrV) genes. MUC5AC and MUC5B (0.5% w/v) suppressed virulence pathways 1, 2, 3, and 6 secretion systems; siderophore biosynthesis; pyoverdine and pyochelin; and quorum sensing. MUC5AC suppressed P. aeruginosa PA01 attachment to plastic and glass surfaces and attachment to live HT human epithelial cells in a concentration-dependent manner. In pig burn wounds injected with P. aeruginosa and PA01 (1 × 105 cfu/mL) and treated with MUC5AC one week later, there were two-log reductions in P. aeruginosa cfu/mL but no reduction without mucins [30].
3.5. Lactobacilli
Lactobacilli enhance phagocytic activity, inhibit neutrophil and macrophage apoptosis, produce lactic acid (which reversibly damages DNA, RNA, and proteins within E. Coli, P. aeruginosa and S. enterica) and also produce hydrogen peroxide, which reduces pyocyanin, elastase, and rhamnolipid produced by P. aeruginosa. Lactobaccillus gasseri supernatant inhibited the growth of P. aeruginosa strain PAO1 on mouse burn wounds, reduced biofilm development 40-fold at 8 h (the control increased significantly), and eliminated biofilms at 28 h. Treatment of the wound did not inhibit P. aeruginosa growth at 24 h but prevented transfer to the blood stream, liver, or spleen and 100% of the mice survived at 7 days. A second dose of the supernatant 24 h after the first dose completely eliminated P. aeruginosa in the wounds. In untreated mice, the death rate due to sepsis was 100%, and the mice had ~107 cfu/mL P. aeruginosa g−1 in their livers and spleens [31].
3.6. Bacteriophages
A systematic review of 95 studies of phage therapy concluded that phage therapy provided 100% protection against MDRO infections and that bio-sanitisation of foods, beverages and surfaces was 100% successful [32]. Alves injected partial-thickness second-degree burn wounds on pig skin (not live pigs) with MRSA252-Rif (resistant to rifampicin) at 104 cfu/mL and then exposed the wounds to Phages DRA88 and SAB4238-A at 109 pfu/mL. On the ex vivo biofilms 24 h after phage treatment, the phage-treated pigskins had 106.5 MRSA252-Rif cfu/mL compared to the control at 107.5 cfu/mL (p ≤ 0.0001). At 48 h after phage treatment, the phage treated pigskins had 107 cfu/mL MRSA252-Rif compared to control 107 cfu/mL (n.s.). Although the results were non-significant at 24 h, there was a 17.25-fold increase in phage numbers and at 48 h a 64.6-fold increase. Why phage numbers increased but infections were not controlled merits investigation [33].
Ho, in four ICU rooms in a 6-month intervention in a 945-bed Taiwanese teaching hospital, used aerosols of eight phages (5.5 × 104 pfu/cm2) and found that Carbapenem-resistant Acinetobacter baumanii (CRAB) rates declined from 8.57/1000 patient-days pre-intervention to 5.11 during the aerosol intervention period (p = 0.0029). Resistant isolates decreased from 87.6% to 46.07% (p = 0.001). Colistin decreased from 7876 DDD/1000 patient-days to 3158 (p =0.0177), tigecycline 2737 to 753 (p = 0.0005), meropenem 5084 to 2469 (p = 0.0385), and imipenem 1384 to 1101 (ns) [34].
Holguín exposed three multiply-drug-resistant P. aeruginosa strains to the ΦPan70 phage at 6.5 × 107 pfu/mL. Eighteen hours after phage therapy, P. aeruginosa P1 decreased from 107.5 to 104 cfu/mL, P2 108 to 104.5, and P4 107.5 to 102.5. In the in vitro biofilms for P1, there was a 17% reduction at 0 h (p = 0.003), 34% at 24 h (p = 0.134), and 55% at 48 h (p = 0.005); for P3 a 59% reduction at 0 h (p = 0.00001), 56% at 24 h (p = 0.034), and 75% at 48 h (p = 0.0004); and for P4 a 68% reduction at 0 h (p = 0.015), 15% at 24 h (p = 0.036) and 21% at 48 h (p = 0.286). When ΦPan70 was given to the mice immediately after P. aeruginosa infection, 4/5 mice survived, and for those who received ΦPan70 45 min after infection, 5/5 survived. For those who received phage therapy 24 and 48 h after infection, 4/5 mice survived, but in the control, all mice died on days 3 or 4 [35].
O’Flaherty in an in vitro study of 28 S. aureus strains from outpatients, inpatients, and hospital staff in Ireland over a 3-year period, found that 14/28 of these S. aureus strains were sensitive to phage K at 107 cfu/mL, no bacteria remained after 2 h, and there were no bacteriophage-insensitive mutants after 25 h. MRSA strain DPC5645 was reduced within 2 h from 5.7 × 106 cfu/mL to undetectable levels, and in vivo MRSA strain DPC5246 on skin was reduced 100-fold with phage K 1.4 × 108 pfu/mL [36].
Pallavali for P. aeruginosa, S. aureus, K. pneumoniae, and E. coli used bacteriophages at 1 × 109 pfu/mL for in vitro biofilms and after 4 h of phage therapy at 96 h, the optical density (which corresponds to biomass) for P. aeruginosa was 0.47 ± 0.035 and decreased to 0.17 ± 0.024; for E. coli it was 0.47 ± 0.035 and decreased to 0.15 ± 0.026, for K. pneumoniae it was 0.47 ± 0.035 and decreased to 0.17 ± 0.022, and for S. aureus it was 0.47 ± 0.036 and decreased to 0.16 ± 0.032. In vitro confocal microscopy showed “predominant numbers of dead cells” after 4 h of phage therapy [37].
3.6.1. Risk of Bias Assessment: Numbers of Bacterial Strains Tested, Numbers of In Vivo Tests Using Animals, and Summary Measures Used in In Vitro and In Vivo Biofilm Outcomes
Authors often tested few bacterial strains, many made no animal studies, and in the animal studies numbers were very small. Of the 31 studies, 6 tested 1 strain of Acinetobacter baumanii, 1 tested 2 strains, 2 tested 3 strains, 1 tested 8 strains, and 1 tested 100 clinical isolates from burn patients. Twelve studies tested one strain of Pseudomonas aeruginosa, two tested two strains, one tested three strains, one tested seven, one tested nine, one tested twelve, one tested twenty-eight strains, and one tested fifty-seven clinical isolates from burn patients. Many tested one strain (PA01), which is not usually pathogenic in humans without risk factors. Eight studies tested one strain of MRSA, one tested twenty-seven, and one tested thirty-six strains. One study tested one strain of Klebsiella pneumoniae, and two tested two strains. Four strains of E. coli were tested. Three studies tested burns on 20 mice, one on 32, one on 35, one on 54, and seven studies did not state the number of mice. One study tested 3 rats and another tested 4 rats. One study tested 3 pigs, one 4, one did not state the number, and one used pigskins and not live pigs (Table 1).

Table 1.
Reductions in vivo and in vitro biofilms after interventions.
The multiple outcomes used by authors made summaries difficult and no meta-analysis could be undertaken. In particular, the variety of microscopy techniques to measure changes in biofilms could not be summarised. Fourteen studies used one or more of these as their main outcome measure: % reductions in the numbers of bacteria forming biofilms or cell counts, intact biofilms, bacterial viability, or biofilm optical density. Of these, 13 reported cfu/mL; 11 reported MICs; 5 reported Minimal Biofilm Inhibitory Concentrations (MBICs); and 3 reported fold-reductions in gene expression or production of reactive oxygen species (ROS). Some reported electron scanning and other microscopy techniques of the effects of interventions on biofilms. Two studies reported if the bacteria were strong, intermediate, or weak biofilm formers [19,23], but in other studies, if there were weak bioformers an intervention could be incorrectly assessed as being the cause of biofilm reduction. The reporting of the technical details of the preparation of samples and measurement methods was extensive, whereas few tested interventions at different strengths or replicated their assays (Table S1).
3.6.2. Identification of Candidate Interventions for Further Testing in Large Scale c-RCTs
It is not possible to identify a simple metric such as reduction in cfu/mL by which to rank studies for further large-scale testing because of the heterogeneity of bacteria tested, whether dose ranging studies were conducted, the number of replications of experiments, whether tested in animals, ability of bacteria to form strong biofilms, biofilm outcome measures, and reporting of scanning electron microscopy of biofilms. Interventions meriting further testing on the basis of one or more outcome measures have been starred *** in Table 1. These include Gholamrezazadeh’s study of nano-silver [6], Pourhajibagher’s study of AgSD-NLs@Cur [14], Halstead’s studies of silver (Acticoat, Mepilex) and acetic acid [13,17], Karaky’s study of graphene–metal combinations [8], Li’s study of CuCo2S4 nanoparticles [15], Song’s study of Chlorhexidene acetate nanoemulsion (CNE) [18], Chhibber’s study of a novel hydrogel (moxifloxacin, carbomer, Chitosan, and Boswellia) [20], Pourhajibagher’s study of nano-emodin to release reactive oxygen species (ROS) [21], Lu’s study of blue light + carvacrol [3], Wang’s study of antimicrobial blue light [4], Banar’s study of mannosidase + trypsin [23], Ghosh’s study of NCK-10, a naphthalene core compound with a decyl chain appendage [1], Han’s study of AMP-jsa9 [25], Ghosh’s study of NCK-10 [24], Konai’s study of D-LANA [5], Memariani’s study of the antimicrobial peptide PV3, which includes two snake venoms [26], Pan’s study of the polypeptides P03 and PL2 [27], and the studies of phage therapy by Ho [34] and O’Flaherty [36]. Replications in large RCTs using animal models then in c-RCTs of patients, surfaces in hospital, and medical facility and medical equipment and using uniform experimental methods would best be accomplished by a large consortium of burn researchers.
4. Discussion
Bacteria are able to colonise and form biofilms on patients, especially those with burns or other wounds, who are immunosuppressed, are frail, have multiple co-morbidities, or have indwelling devices. Bacteria are able to colonise and form biofilms on multiple surfaces in medical facilities, on surfaces that patients contact and on medical equipment. Thus, a comprehensive and integrated approach is needed to identify and test interventions to treat and eliminate biofilms in all of these areas. Burn patients are at risk because they are colonised with Gram-positives and then with multiply-drug-resistant Gram-negatives. Many interventions both current and at the laboratory stage are potentially applicable to both patients and the surfaces and equipment in hospitals they are admitted to. These interventions have been starred *** in Table 1 although the studies are small.
More intensive research is also needed to increase the effectiveness of current cleaning and disinfecting routines by making surfaces in medical facility patient rooms, treatment and common rooms, and kitchens less hospitable to bacteria and viruses by plating them with copper, silver, titanium, and other metals; impregnating curtains, bedding, and gowns with antibacterial chemicals; using ultraviolet light to disinfect keyboards on computers, phones, cell phones, and shoes, which transfer pathogens between floors and rooms.
If the following surfaces are not included in current cleaning and disinfection routines, they need to be added and carefully monitored: sinks, drains, and toilets with their constant fluid flows are good environments for biofilms, and aerosols result when fluids are poured into them. One study of a hospital and its related LTCF found that a peracetic acid and hydrogen peroxide foam reduced Gram-negatives for an average of three days, but repeated treatments were needed [38]. Kitchens and food service also need to be included in disinfection routines and monitoring.
A review of infection and prevention guidelines (search to April 2019) identified 31 guidelines with 1855 recommendations and rated 28 (1.5%) of the recommendations as based on systematic reviews and meta-analyses of RCTs, with 13 recommendations for devices and 311 (16.8%) recommendations as based on well-designed RCTs with strong recommendation from high-quality evidence with 64 recommendations for devices. However, only six guidelines had a GRADE recommendation. A major initiative is needed to assess the strength of evidence for cleaning medical equipment [39].
Cleaning biofilms from endoscopes is a major concern. A study of P. aeruginosa biofilms showed that they were eliminated only with 2500 ppm of peracetic acid, whereas planktonic cells were eliminated with 20 ppm [40]. A study of a quality circle to improve biofilm removal from urological endoscopes used 500 relative light units/piece as a measure of endoscopic biofilm clearance and noted an improvement from 50% to 90% when the quality circle recommendations were followed [41]. Enzymatic and alkaline detergents, bristle brushes, and Pull Thru channel cleaners to sterilise endoscopes were compared to a water flush for 108 cfu/mL of P. aeruginosa and E. faecalis, and it was found that friction applied to all surfaces of the endoscope was crucial to remove enough biofilm, and then glutaraldehyde could kill the remaining microorganisms [42]. A study of argon-plasma-activated gas on endoscopes contaminated with biofilms of MRSA, S. aureus, P. aeruginosa, or E. coli found that there was >8 log reduction in viable cells and the dispersal of 24 and 48 h biofilms of all bacteria [43].
Pathogens are transferred between rooms on the shoes of staff and patients. After shoe soles were decontaminated with ultraviolet light, there were significant reductions (p < 0.01) on floors, beds, furniture, and patient dummies of log10 2.8 for E. coli, S. aureus, and E. faecalis but not for C. difficile [44].
Beds and mattresses are the surfaces patients spend most time in contact with. They often become soiled and are a good residence for bacteria. A small study of a surgery ward randomised beds to launderable or non-launderable covers and the launderable covers had significantly lower cfu counts/30 cm2 both after admission and discharge (p < 0.001) [45]. On a 36-bed medical ward 77% of bedside surfaces in contact with MRSA-negative patients were MRSA positive and 83% of bedside surfaces of MRSA-positive patients. One hour after hypochlorite disinfection, 7.4% of the bedside tables and 17.6% of the bedrails were MRSA positive, but four hours later, bedside surface contamination increased by 80% (p < 0.01). Using hypochlorite plus nano-organosilicon quaternary ammonium chloride spray, the MRSA level by midday declined from 4.4 ± 8.7 cfu/cm2 to 0.07 ± 2.6 [46]. Two LTCFs laundered bed covers with, chlorine, detergent and hot water and the C. difficile rate decreased by 49% [47].
This review found no RCTs of interventions to assess if there were biofilms on computers, phones, tablets, or cell phones, which are widely used by hospital and LTCF staff. Computers are used frequently by HCWs for patient care, and physicians frequently search their tablets and cell phones for data. A study of hand calculators cleaned with QUAT found after an average of 73 keystrokes that 80% of the QUAT had been removed [48]. More effective was ultraviolet light: a study of clinicians’ smartphones and wearable devices detected pathogenic bacteria on 20%, but there was a significant reduction to 4% after 30 s of ultraviolet light (p = 0.002) [49]. However, none of these studies mentioned or researched biofilms, and an essential part of an integrated approach to removing biofilms is to test all of these surfaces for biofilms and then set in place monitoring processes to remove and ensure that surfaces remain free of biofilms.
The key problem thus becomes to comprehensively and accurately detect biofilms on surfaces in medical institutions (especially high touch surfaces) and on medical equipment. The current research methods of identifying and characterising biofilms are to take samples, culture them, and submit them to PCR testing and microscopy. Staining methods include the nucleic acid dye SYT09, which penetrates the membranes of bacterial cells and attaches to the DNA of both live and dead cells and fluoresces green, and the PI dye, which attaches only to dead bacterial cells and fluoresces red [50]. A key issue is whether interventions designed to identify biofilms in human burns and other wounds can also be used cost-effectively to detect biofilms on surfaces and equipment.
Other approaches are to blot or to sonicate surfaces and wounds. A small study blotted rats’ burn wounds with a membrane to take a sample of the biofilm infected with Ps. aeruginosa PA01 and found that the alcian blue stain correlated 100% with the native PAGE (polyacrylamide gel electrophoresis) test, which quantifies total biofilm biomass [51]. A study of orthopaedic screws sonicated and cultured the S. aureus, P. aeruginosa, and C. albicans in the biofilms and found that MALDI-TOF mass spectrometry detected microorganisms with 99.9% reliability [52]).
The lengthy process of culturing biofilm organisms may be bypassed with optical methods. A review assessed multiple methods of optical identification of bacteria including infrared spectroscopy, Fourier-transformed infrared spectroscopy, ultraviolet resonance, Raman spectroscopy, surface-enhanced Raman spectroscopy, fluorescence spectroscopy, and optical coherence tomography and concluded that Fourier-transformed infrared spectroscopy provides superior use of data particularly linked to large databases of bacterial characteristics [53].
Bacteriological typing methods include cultures, amplification, fragment, genome, and sequence methods. Amplification methods include random amplification of polymorphic DNA, rep-PCR, arbitrarily primed-PCR, and variable number repeat typing. Rep-PCR amplifies repetitive intergenic sequences, which are then subjected to electrophoresis. Fragment methods use restriction enzymes to digest DNA, and pulsed-field gel electrophoresis is the standard reference-typing method. In sequence amplification, either single-locus or multilocus genes are compared to international standards and both housekeeping and virulence genes can be assessed. In genomic methods, next-generation sequencing is the reference standard because of accurate phenotyping of many genes and advanced computerisation [54]. Because bacteria in biofilms may be in starvation mode and thus less active or are persister bacteria, the best method in the case of biofilms on hospital surfaces and medical equipment will depend on the ability to actually identify biofilms and recover bacteria from them.
In view of the high cost of biofilms in terms of morbidity, mortality, and medical costs, intensive research on optimal comprehensive and cost-effective methods of detecting biofilms on medical surfaces and equipment is a priority. Even if the detection methods are initially expensive, the costs of biofilms to patients are enormous.
Strengths: The systematic review was conducted with no language or date limits. The study focused on a comprehensive approach to identify both current and novel interventions to detect and destroy biofilms in burn wounds, surfaces, and medical equipment patient contacts in hospitals or LTCFs.
Weaknesses: The key problems are the heterogeneity in the numbers of bacteria tested, comparator interventions, dose ranging studies, replication of experiments, in vivo tests, outcome measures, and microscopy methods. Many studies did not identify if they were testing bacteria that were strong, intermediate, or weak biofilm formers, and thus the effects of interventions could have been incorrectly attributed. Simple metrics such as enough reductions in cfu/mL are not available to rank studies for further large-scale testing. Interventions with positive outcome measures which merit testing are starred *** in Table 1.
5. Conclusions
The essential step is to reduce entry of biofilms into hospitals by establishing isolation rooms and providing the staff to administer tests there to ensure that when patients are admitted they are tested for MDROs, HAIs, and biofilms and are treated before being admitted to the hospital or returned to long-term care homes. The admission of patients with established biofilms needs to be conducted with full precautions.
An integrated and comprehensive approach is required to detect and eliminate biofilms on both patients and on surfaces and equipment they contact. This review identified 20 interventions that merit further testing and replication in laboratory RCTs and then in large c-RCTs with patients, hospital surfaces, and equipment. The large number of studies required would best be conducted by a consortium of burn centres and infectious disease specialists. These interventions include phages, nano-silver, AgSD-NLs@Cur, silver in the form of Acticoat and Mepilex, acetic acid, graphene-metal combinations, CuCo2SO4 nanoparticles, Chlorhexidene acetate nanoemulsion, a hydrogel with moxifloxacin, carbomer, Chitosan and Boswellia, light therapy using LEDs with nano-emodin to release reactive oxygen species, blue light + Carvacrol, antimicrobial blue light, mannosidase + trypsin, NCK-10 (a napthalene core compound with a decyl chain appendage), the antimicrobial peptide PV3, which includes two snake venoms, and the polypeptides P03 and PL2. Most of these interventions are aimed at penetrating cell membranes.
Some current cleaning routines monitor thoroughness of cleaning by environmental service workers by touching random samples of surfaces with invisible markers then use fluorescent light detectors to see if the markers have been removed. The reliability of methods such as blotting and sonification of surfaces to detect biofilms and the effectiveness of their removal needs to be tested.
The number and types of surfaces and equipment currently cleaned and disinfected needs to be substantially augmented to include keyboards, tablets, cell phones, medical equipment (especially endoscopes), sinks, drains, and food preparation in kitchens.
Supplementary Materials
The following are available online at https://www.mdpi.com/article/10.3390/ijerph182413195/s1, Table S1. Interventions to reduce infections in burns.
Author Contributions
Conceptualisation, R.E.T.; methodology, R.E.T.; software, R.E.T.; validation, R.E.T., B.C.T.; formal analysis, R.E.T., B.C.T.; data curation, R.E.T.; writing—original draft preparation, R.E.T.; writing—review and editing, R.E.T., B.C.T.; visualisation, R.E.T.; project administration, R.E.T. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Goodwine, J.; Gil, J.; Doiron, A.; Valdes, J.; Solis, M.; Higa, A.; Davis, S.; Sauer, K. Pyruvate-depleting conditions induce biofilm dispersion and enhance the efficacy of antibiotics in killing biofilms in vitro and in vivo. Sci. Rep. 2019, 9, 1–16. [Google Scholar] [CrossRef]
- Pourhajibagher, M.; Mahmoudi, H.; Rezaei-Soufi, L.; Alikhani, M.Y.; Bahador, A. Potentiation effects of antimicrobial photodynamic therapy on quorum sensing genes expression: A promising treatment for multi-species bacterial biofilms in burn wound infections. Photodiagn. Photodyn. Ther. 2020, 30, 101717. [Google Scholar] [CrossRef]
- Lu, M.; Wang, S.; Wang, T.; Hu, S.; Bhayana, B.; Ishii, M.; Kong, Y.; Cai, Y.; Dai, T.; Cui, W.; et al. Bacteria-specific phototoxic reactions triggered by blue light and phytochemical carvacrol. Sci. Transl. Med. 2021, 13, eaba3571. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wu, X.; Chen, J.; Amin, R.; Lu, M.; Bhayana, B.; Zhao, J.; Murray, C.K.; Hamblin, M.R.; Hooper, D.C.; et al. Antimicrobial Blue Light Inactivation of Gram-Negative Pathogens in Biofilms: In Vitro and In Vivo Studies. J. Infect. Dis. 2016, 213, 1380–1387. [Google Scholar] [CrossRef] [PubMed]
- Konai, M.M.; Haldar, J. Lysine-Based Small Molecule Sensitizes Rifampicin and Tetracycline against Multidrug-Resistant Acinetobacter baumannii and Pseudomonas aeruginosa. ACS Infect. Dis. 2019, 6, 91–99. [Google Scholar] [CrossRef] [PubMed]
- Gholamrezazadeh, M.; Shakibaie, M.R.; Monirzadeh, F.; Masoumi, S.; Hashemizadeh, Z. Effect of nano-silver, nano-copper, deconex and benzalkonium chloride on biofilm formation and expression of transcription regulatory quorum sensing gene (rh1 R) in drug-resistance Pseudomonas aeruginosa burn isolates. Burns 2018, 44, 700–708. [Google Scholar] [CrossRef]
- Yali, G.; Jing, C.; Chunjiang, L.; Cheng, Z.; Xiaoqiang, L.; Yizhi, P. Comparison of pathogens and antibiotic resistance of burn patients in the burn ICU or in the common burn ward. Burns 2014, 40, 402–407. [Google Scholar] [CrossRef]
- Karaky, N.; Kirby, A.; McBain, A.; Butler, J.; El Mohtadi, M.; Banks, C.E.; Whitehead, K.A. Metal ions and graphene-based compounds as alternative treatment options for burn wounds infected by antibiotic-resistant Pseudomonas aeruginosa. Arch. Microbiol. 2020, 202, 995–1004. [Google Scholar] [CrossRef]
- Andersson, M.; Madsen, L.; Schmidtchen, A.; Puthia, M. Development of an Experimental Ex Vivo Wound Model to Evaluate Antimicrobial Efficacy of Topical Formulations. Int. J. Mol. Sci. 2021, 22, 5045. [Google Scholar] [CrossRef]
- Thomas, R.E.; Thomas, B.C.; Conly, J.; Lorenzetti, D. Cleaning and disinfecting surfaces in hospitals and long-term care facilities for reducing hospital and facility-acquired bacterial and viral infections: A systematic review. J. Hosp. Infect. 2021, in press. [Google Scholar]
- Stewart, P.S.; Parker, A.E. Measuring Antimicrobial Efficacy against Biofilms: A Meta-analysis. Antimicrob. Agents Chemother. 2019, 63, 1–11. [Google Scholar] [CrossRef]
- Schwarzer, S.; James, G.A.; Goeres, D.; Bjarnsholt, T.; Vickery, K.; Percival, S.L.; Stoodley, P.; Schultz, G.; Jensen, S.O.; Malone, M. The efficacy of topical agents used in wounds for managing chronic biofilm infections: A systematic review. J. Infect. 2020, 80, 261–270. [Google Scholar] [CrossRef]
- Halstead, F.D.; Rauf, M.; Bamford, A.; Wearn, C.M.; Bishop, J.R.; Burt, R.; Fraise, A.P.; Moiemen, N.S.; Oppenheim, B.A.; Webber, M.A. Antimicrobial dressings: Comparison of the ability of a panel of dressings to prevent biofilm formation by key burn wound pathogens. Burns 2015, 41, 1683–1694. [Google Scholar] [CrossRef] [PubMed]
- Pourhajibagher, M.; Partoazar, A.; Alaeddini, M.; Etemad-Moghadam, S.; Bahador, A. Photodisinfection effects of silver sulfadiazine nanoliposomes doped-curcumin on Acinetobacter baumannii: A mouse model. Nanomedicine 2020, 15, 437–452. [Google Scholar] [CrossRef]
- Li, D.; Guo, Q.; Ding, L.; Zhang, W.; Cheng, L.; Wang, Y.; Xu, Z.; Wang, H.; Gao, L. Bimetallic CuCo 2 S 4 Nanozymes with Enhanced Peroxidase Activity at Neutral pH for Combating Burn Infections. ChemBioChem 2020, 21, 2620–2627. [Google Scholar] [CrossRef] [PubMed]
- Nozari, M.; Gholizadeh, M.; Oghani, F.Z.; Tahvildari, K. Studies on novel chitosan/alginate and chitosan/bentonite flexible films incorporated with ZnO nano particles for accelerating dermal burn healing: In vivo and in vitro evaluation. Int. J. Biol. Macromol. 2021, 184, 235–249. [Google Scholar] [CrossRef]
- Halstead, F.D.; Rauf, M.; Moiemen, N.S.; Bamford, A.; Wearn, C.M.; Fraise, A.P.; Lund, P.A.; Oppenheim, B.A.; Webber, M.A. The Antibacterial Activity of Acetic Acid against Biofilm-Producing Pathogens of Relevance to Burns Patients. PLoS ONE 2015, 10, e0136190. [Google Scholar] [CrossRef]
- Song, Z.; Sun, H.; Yang, Y.; Jing, H.; Yang, L.; Tong, Y.; Wei, C.; Wang, Z.; Zou, Q.; Zeng, H. Enhanced efficacy and anti-biofilm activity of novel nanoemulsions against skin burn wound multi-drug resistant MRSA infections. Nanomed. Nanotechnol. Biol. Med. 2016, 12, 1543–1555. [Google Scholar] [CrossRef] [PubMed]
- Tiwari, S.; Rajak, S.; Mondal, D.P.; Biswas, D. Sodium hypochlorite is more effective than 70% ethanol against biofilms of clinical isolates of Staphylococcus aureus. Am. J. Infect. Control 2018, 46, e37–e42. [Google Scholar] [CrossRef]
- Chhibber, T.; Gondil, V.S.; Sinha, V.R. Development of Chitosan-Based Hydrogel Containing Antibiofilm Agents for the Treatment of Staphylococcus aureus–Infected Burn Wound in Mice. AAPS Pharm. Sci. Tech. 2020, 21, 43. [Google Scholar] [CrossRef] [PubMed]
- Pourhajibagher, M.; Rahimi-esboei, B.; Ahmadi, H.; Bahador, A. The anti-biofilm capability of nano-emodin mediated sonodynamic therapy on multi-species biofilms produced by burn wound bacterial strains. Photodiagn. Photodyn. Ther. 2021, 24, 102288. [Google Scholar] [CrossRef]
- Ishiwata, N.; Tsunoi, Y.; Sarker, R.R.; Haruyama, Y.; Kawauachi, S.; Sekine, Y.; Onuma, C.; Tsuda, H.; Saitoh, D.; Nichidate, I.; et al. Control of burn wound infection by methylene blue-mediated photodynamic treatment with light-emitting diode array illumination in rats. Lasers Surg. Med. 2021, 53, 1238–1246. [Google Scholar] [CrossRef]
- Banar, M.; Emaneini, M.; Satarzadeh, M.; Abdellahi, N.; Beigverdi, R.; Leeuwen, W.B.; Jabalameli, F. Evaluation of Mannosidase and Trypsin Enzymes Effects on Biofilm Production of Pseudomonas aeruginosa Isolated from Burn Wound Infections. PLoS ONE 2016, 11, e0164622. [Google Scholar]
- Ghosh, C.; Manjunath, G.B.; Konai, M.M.; Uppu, D.S.; Paramanandham, K.; Shome, B.R.; Ravikumar, R.; Haldar, J. Aryl-alkyl-lysines: Membrane-Active Small Molecules Active against Murine Model of Burn Infection. ACS Infect. Dis. 2016, 2, 111–122. [Google Scholar] [CrossRef] [PubMed]
- Han, J.; Ma, Z.; Gao, P.; Lu, Z.; Liu, H.; Gao, L.; Lu, W.; Ju, X.; Lv, F.H.; Bie, X. The antibacterial activity of LI-F type peptide against methicillin-resistant Staphylococcus aureus (MRSA) in vitro and inhibition of infections in murine scalded epidermis. Appl. Microbiol. Biotechnol. 2018, 102, 2301–2311. [Google Scholar] [CrossRef]
- Memariani, H.; Shahbazzadeh, D.; Sabatier, J.M.; Memariani, M.; Karbalaeimahdi, A.; Bagheri, K.P. Mechanism of action and in vitro activity of short hybrid antimicrobial peptide PV3 against Pseudomonas aeruginosa. Biochem. Biophys. Res. Commun. 2016, 479, 103–108. [Google Scholar] [CrossRef]
- Pan, M.; Lu, C.; Zheng, M.; Zhou, W.; Song, F.; Chen, W.F.; Liu, D.; Cai, J. Unnatural Amino-Acid-Based Star-Shaped Poly(l-Ornithine)s as Emerging Long-Term and Biofilm-Disrupting Antimicrobial Peptides to Treat Pseudomonas aeruginosa-Infected Burn Wounds. Adv. Healthc. Mater. 2020, 9, e2000647. [Google Scholar] [CrossRef] [PubMed]
- Su, M.; Qiu, L.; Deng, Y.; Ruiz, C.H.; Rudolf, J.D.; Dong, L.B.; Feng, X.; Cameron, M.D.; Shen, B.; Duan, Y.; et al. Evaluation of Platensimycin and Platensimycin-Inspired Thioether Analogues against Methicillin-Resistant Staphylococcus aureus in Topical and Systemic Infection Mouse Models. Mol. Pharm. 2019, 16, 3065–3071. [Google Scholar] [CrossRef]
- Uusitalo, P.; Hagglund, U.; Rhoos, E.; Scherman, N.H.; Elofsson, M.; Sundin, C. The salicylidene acylhydrazide INP0341 attenuates Pseudomonas aeruginosa virulence in vitro and in vivo. J. Antibiot. 2017, 70, 937–943. [Google Scholar] [CrossRef]
- Wheeler, K.M.; Carcamo-Oyarce, G.; Turner, B.S.; Dellos-Nolan, S.; Co, J.Y.; Lehoux, S.; Cummings, R.D.; Wozniak, D.J.; Ribbeck, K. Mucin glycans attenuate the virulence of Pseudomonas aeruginosa in infection. Nat. Microbiol. 2019, 4, 2146–2154. [Google Scholar] [CrossRef] [PubMed]
- Lenzmeier, T.D.; Mudaliar, N.S.; Stanbro, J.A.; Watters, C.; Ahmad, A.; Simons, M.P.; Ventolini, G.; Zak, J.C.; Colmer-Hamood, J.A.; Hamood, A.N. Application of Lactobacillus gasseri 63 AM supernatant to Pseudomonas aeruginosa-infected wounds prevents sepsis in murine models of thermal injury and dorsal excision. J. Med. Microbiol. 2019, 68, 1560–1572. [Google Scholar] [CrossRef] [PubMed]
- Ssekatawa, K.; Byarugaba, D.K.; Kato, C.D.; Wampande, E.M.; Ejobi, F.; Tweyongyere, R.; Nakavuma, J.L. A review of phage mediated antibacterial applications. Alex. J. Med. 2020, 57, 1–20. [Google Scholar] [CrossRef]
- Alves, D.R.; Booth, S.P.; Scavone, P.; Schellenberger, P.; Salvage, J.; Dedi, C.; Thet, N.T.; Jenkins, A.T.A.; Waters, R.; Ng, K.W.; et al. Development of a High-Throughput ex-Vivo Burn Wound Model Using Porcine Skin, and Its Application to Evaluate New Approaches to Control Wound Infection. Front. Cell Infect. Microbiol. 2018, 8, 196. [Google Scholar] [CrossRef] [PubMed]
- Ho, Y.H.; Tseng, C.C.; Wang, L.S.; Chen, Y.T.; Ho, G.J.; Lin, T.Y.; Wang, L.Y.; Chen, L.K. Application of Bacteriophage-containing Aerosol against Nosocomial Transmission of Carbapenem-Resistant Acinetobacter baumannii in an Intensive Care Unit. PLoS ONE. 2016, 11, e0168380. [Google Scholar] [CrossRef] [PubMed]
- Holguin, A.V.; Rangel, G.; Clavijo, V.; Prada, C.; Mantilla, M.; Gomez, M.C.; Kutter, E.; Taylor, C.; Fineran, P.C.; Barrios, A.F.; et al. Phage PHIPan70, a Putative Temperate Phage, Controls Pseudomonas aeruginosa in Planktonic, Biofilm and Burn Mouse Model Assays. Viruses 2015, 7, 4602–4623. [Google Scholar] [CrossRef] [PubMed]
- O’Flaherty, S.; Ross, R.P.; Meaney, W.; Fitzgerald, G.F.; Elbreki, M.F.; Coffey, A. Potential of the polyvalent anti-Staphylococcus bacteriophage K for control of antibiotic-resistant staphylococci from hospitals. Appl. Environ. Microbiol. 2005, 71, 1836–1842. [Google Scholar] [CrossRef]
- Pallavali, R.R.; Degati, V.L.; Narala, V.R.; Velpula, K.K.; Yenugu, S.; Durbaka, V.R.P. Lytic bacteriophages against bacterial biofilms formed by multidrug-resistant Pseudomonas aeruginosa, Escherichia coli, Klebsiella pneumoniae, and Staphylococcus aureus isolated form burn wounds. PHAGE Ther. Appl. Res. 2021, 2, 120–130. [Google Scholar]
- Jones, L.D.; Mana, T.S.C.; Cadnum, J.L.; Jencson, A.L.; Silva, S.Y.; Wilson, B.M.; Donskey, C.J. Effectiveness of foam disinfectants in reducing sink-drain gram-negative bacterial colonization. Infect Control Hosp. Epidemiol. 2020, 41, 280–285. [Google Scholar] [CrossRef]
- Mitchell, B.G.; Fasugba, O.; Russo, P.L. Where is the strength of evidence? A review of infection prevention and control guidelines. J. Hosp. Inf. 2020, 105, 242–252. [Google Scholar] [CrossRef]
- Akinbobola, A.B.; Sherry, L.; Mckay, W.G.; Ramage, G.; Williams, C. Tolerance of Pseudomonas aeruginosa in in-vitro biofilms to high-level peracetic acid disinfection. J. Hosp. Infect. 2017, 97, 162–168. [Google Scholar] [CrossRef]
- Luo, Y.; Yang, Q.; Li, B.; Yao, Y. Establishment of a quality control circle to reduce biofilm formation in flexible endoscopes by improvement of qualified cleaning rate. J. Intern. Med. Res. 2020, 48, 300060520952983. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, M.M.; Graziano, K.U.; Olson, N.; Franca, R.; Alfa, M.J. The polytetrafluoroethylene (PTFE) channel model of cyclic-buildup biofilm and traditional biofilm: The impact of friction, and detergent on cleaning and subsequent high-level disinfection. Infect Control Hosp. Epidemiol. 2020, 41, 172–180. [Google Scholar] [CrossRef]
- Bhatt, S.; Mehta, P.; Chen, C.; Schneider, C.L.; White, L.N.; Chen, H.L.; Kong, M.G. Efficacy of low-temperature plasma-activated gas disinfection against biofilm on contaminated GI endoscope channels. Gastrointest Endosc. 2019, 89, 105–114. [Google Scholar] [CrossRef] [PubMed]
- Rashid, T.; Poblete, K.; Amadio, J.; Hasan, I.; Begum, K.; Alam, M.J.; Garey, K.W. Evaluation of a shoe sole UVC device to reduce pathogen colonization on floors, surfaces and patients. J. Hosp. Infect. 2018, 98, 96–101. [Google Scholar] [CrossRef]
- Hooker, E.A.; Allen, S.; Gray, L.; Kaufman, C. A randomized trial to evaluate a launderable bed protection system for hospital beds. Antimicrob. Resist. Infect Control 2012, 1, 1–27. [Google Scholar] [CrossRef]
- Yuen, J.W.M.; Chung, T.W.K.; Loke, A.Y. Methicillin-resistant Staphylococcus aureus (MRSA) contamination in bedside surfaces of a hospital ward and the potential effectiveness of enhanced disinfection with an antimicrobial polymer surfactant. Int. J. Environ. Res. Public Health 2015, 12, 3026–3041. [Google Scholar] [CrossRef]
- Hooker, E.A.; Bochan, M.; Reiff, T.T.; Blackwell, C.; Webb, K.W.; Hart, K.W. Decreasing Clostridium difficile health care-associated infections through use of a launderable mattress cover. Am. J. Infect Control 2015, 43, 1326–1330. [Google Scholar] [CrossRef]
- Burns, S.; Tulpinski, J.; Som, A.; Taylor, T. Pushing our buttons: How many cycles does it take to remove an active disinfecting compound from a keypad? Am. J. Infect Control 2014, 42, S39–S40. [Google Scholar] [CrossRef]
- Gostine, A.; Gostine, D.; Donohue, C.; Carlstrom, L. Evaluating the effectiveness of ultraviolet-C lamps for reducing keyboard contamination in the intensive care unit: A longitudinal analysis. Am. J. Infect Control 2016, 44, 1089–1094. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Curtin, A.M.; Buckley, H.L. Biofouling detection methods that are widely applicable and useful across disciplines: A mini-review. Biofouling 2021, 37, 494–505. [Google Scholar] [CrossRef]
- Astada, A.; Nakagami, G.; Minematsu, T.; Kitamura, A.; Mugita, Y.; Sanada, H. Concurrent validity of biofilm detection by wound blotting on hard-to-heal wounds. J. Wound Care 2021, 30, S4–S13. [Google Scholar] [CrossRef]
- Cieslinski, J.; Ribeiro, V.S.T.; Kraft, L.; Suss, P.H.; Rosa, E.; Morello, L.G.; Pillonetto, M.; Tuon, F.F. Direct detection of microorganisms in sonicated orthopedic devices after in vitro biofilm production and different processing conditions. Eur. J. Orthop. Surg. Traumatol. 2021, 31, 1113–1120. [Google Scholar] [CrossRef] [PubMed]
- Locke, A.; Fitzgerald, S.; Mahadevan-Jansen, A. Advances in optical detection of human-associated pathogenic bacteria. Molecules 2020, 25, 5256. [Google Scholar] [CrossRef] [PubMed]
- Nutman, A.; Marchaim, D. How to: Molecular investigation of a hospital outbreak. Clin. Microbiol. Infect. 2019, 25, 688–695. [Google Scholar] [CrossRef] [PubMed]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).