
Seven Years Study of the Seasonal Dynamics of Zooplankton Communities in a Large Subtropical Floodplain Ecosystem: A Test of the PEG Model

Abstract
:1. Introduction
2. Materials and Methods
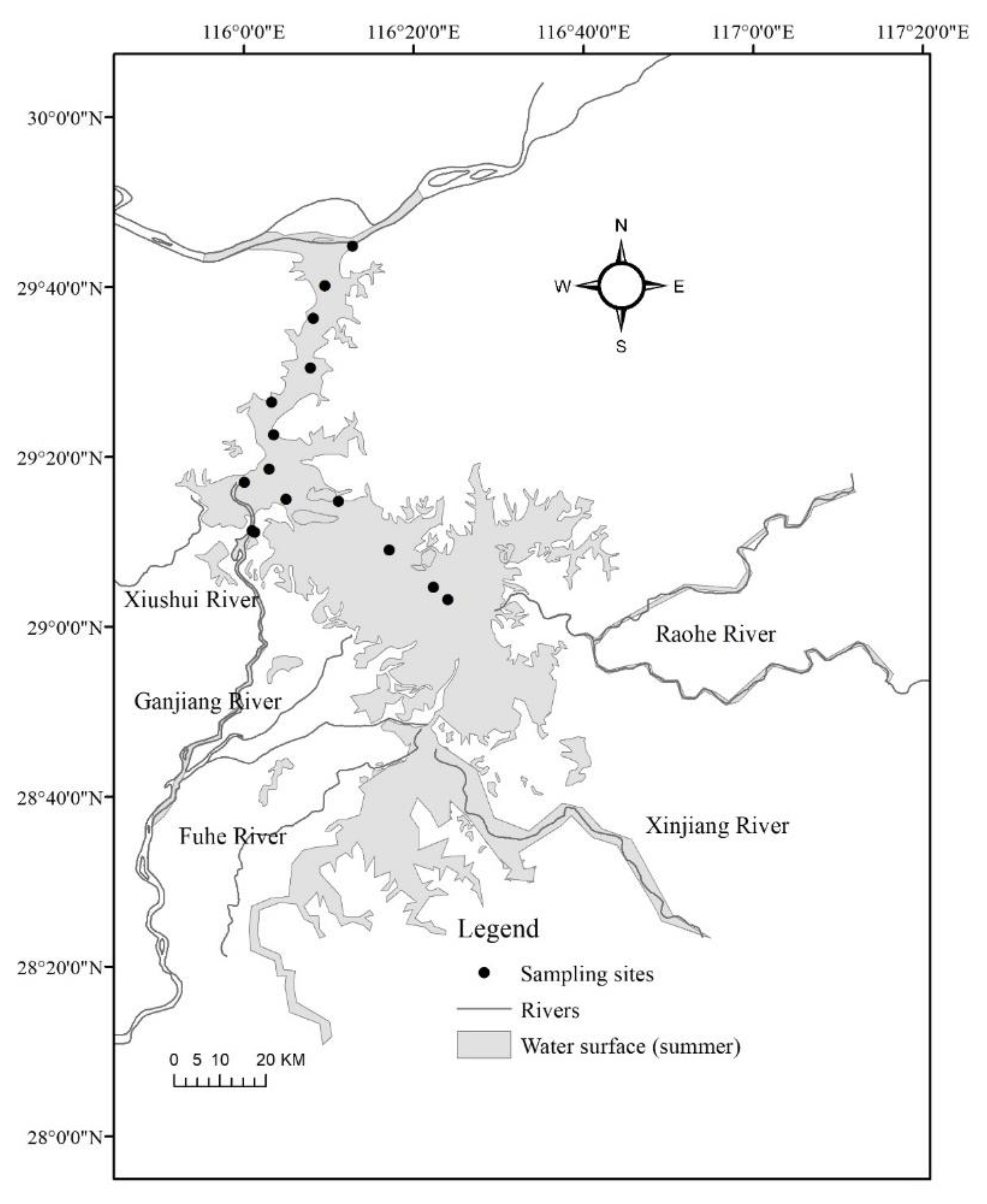
2.1. Study Area
2.2. Sampling
2.3. Statistical Analysis
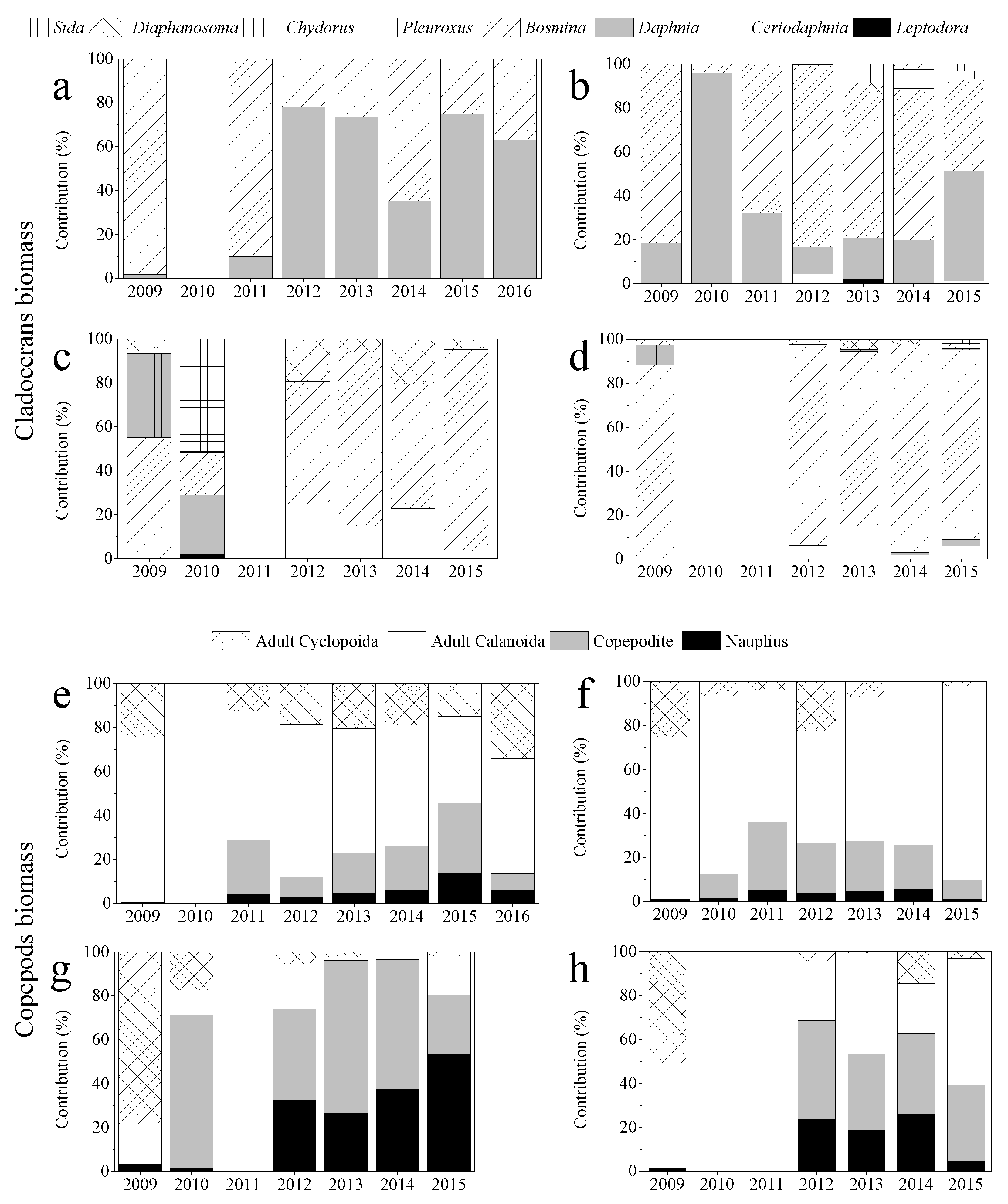
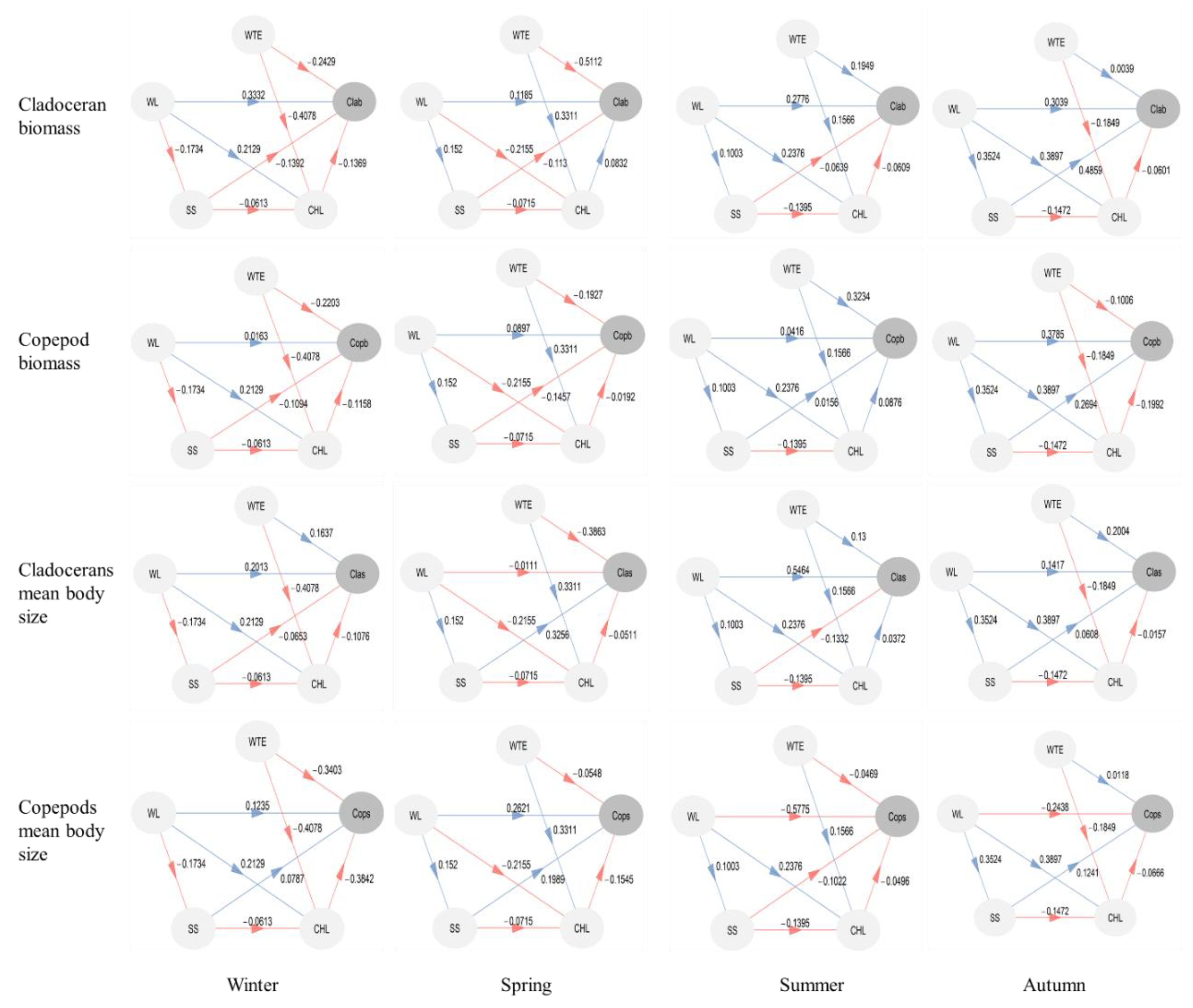
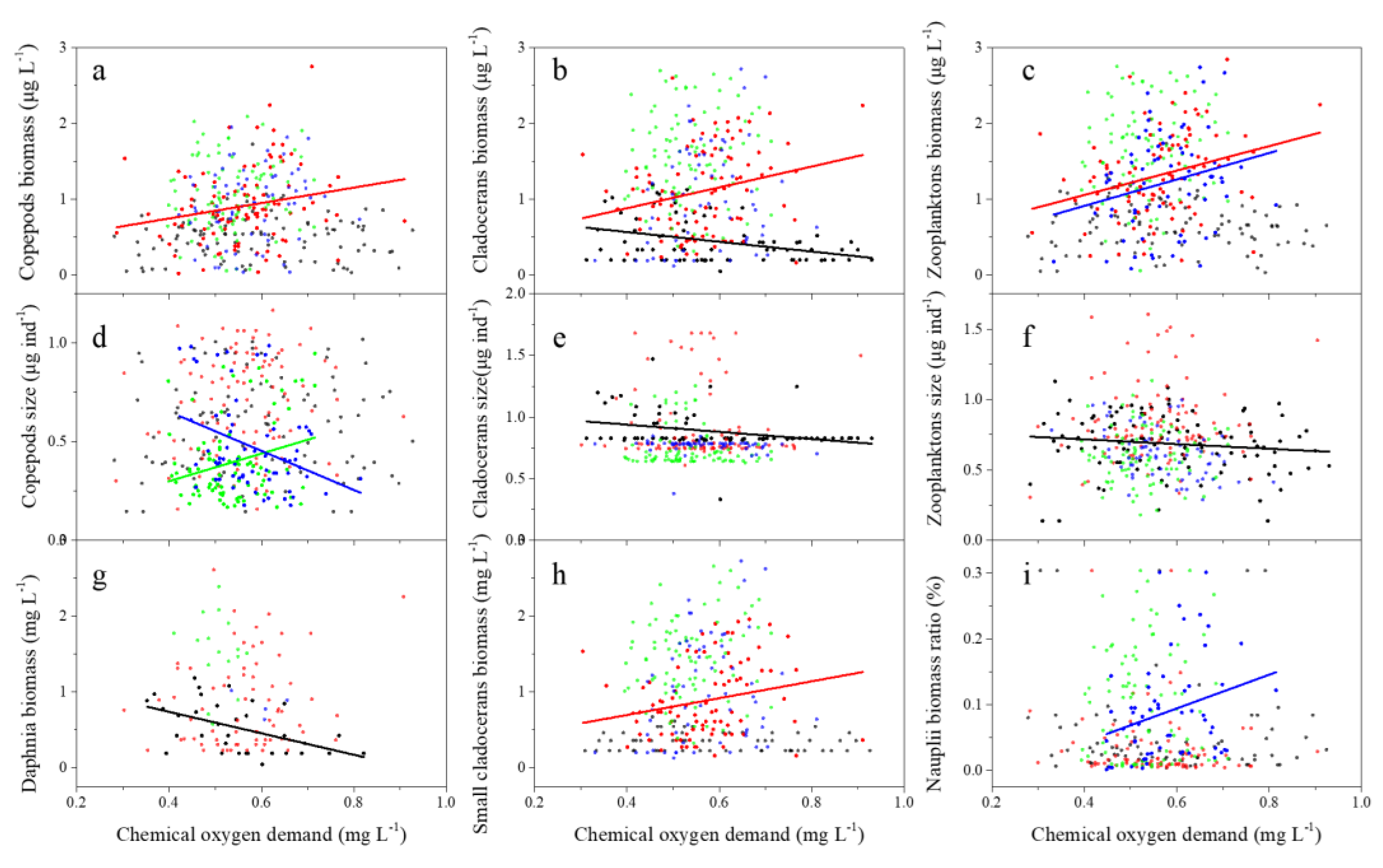
3. Results
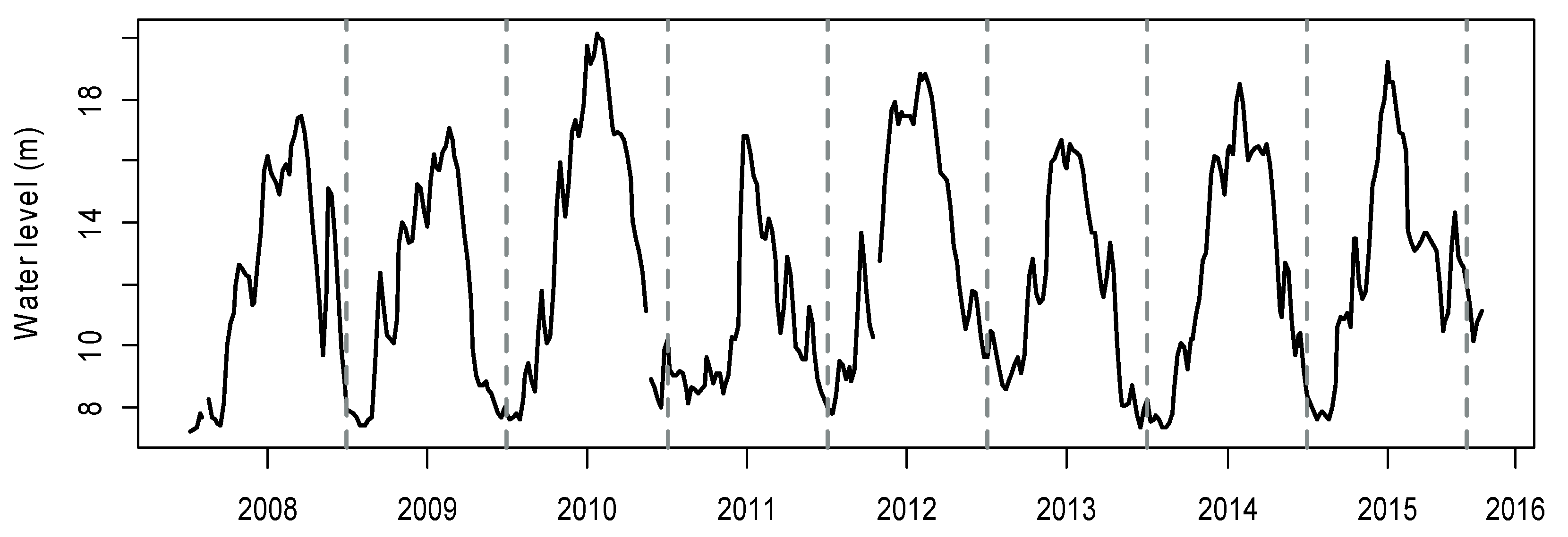
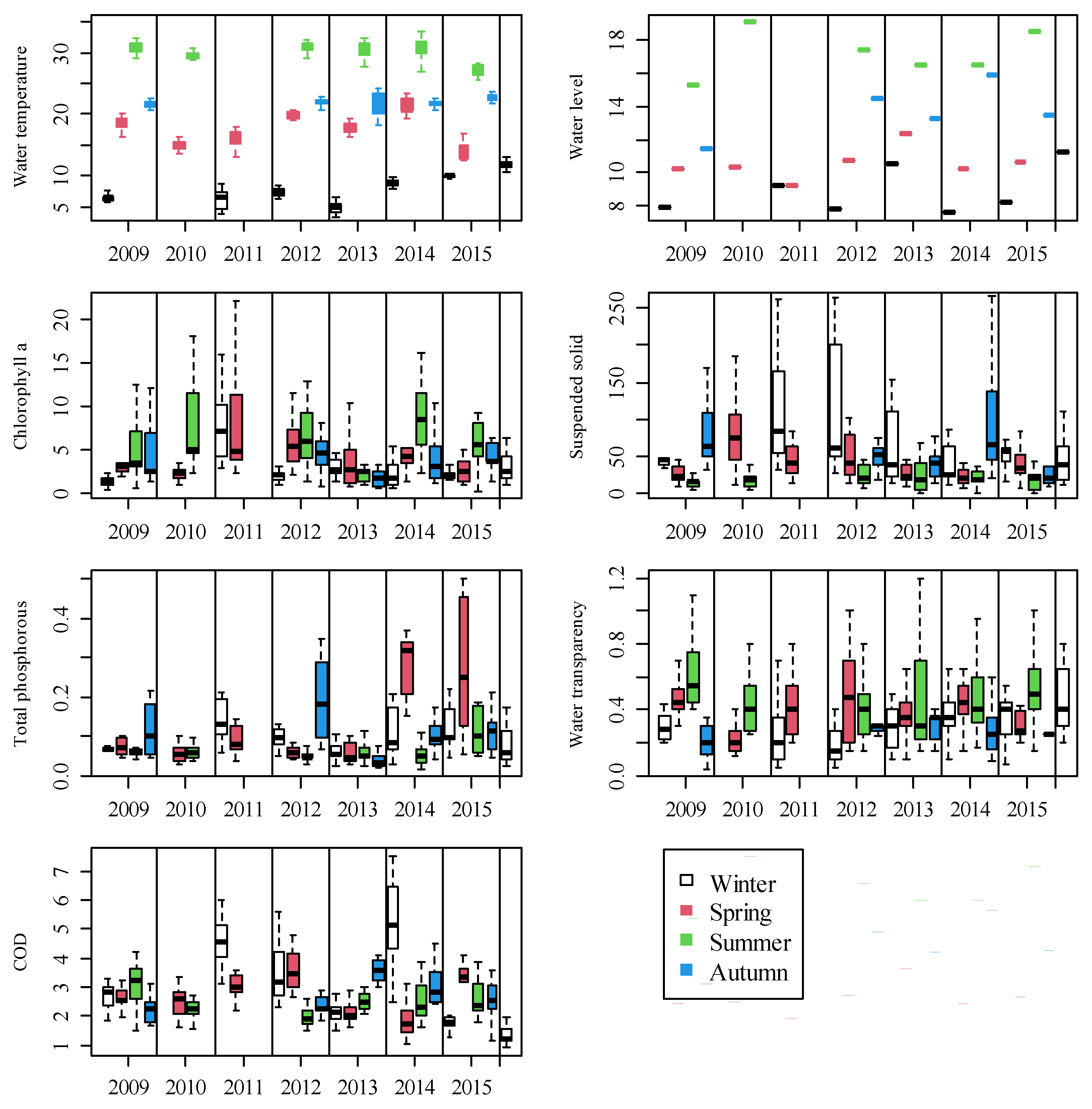
3.1. Inter- and Intra-Year Variations of Environmental Variables
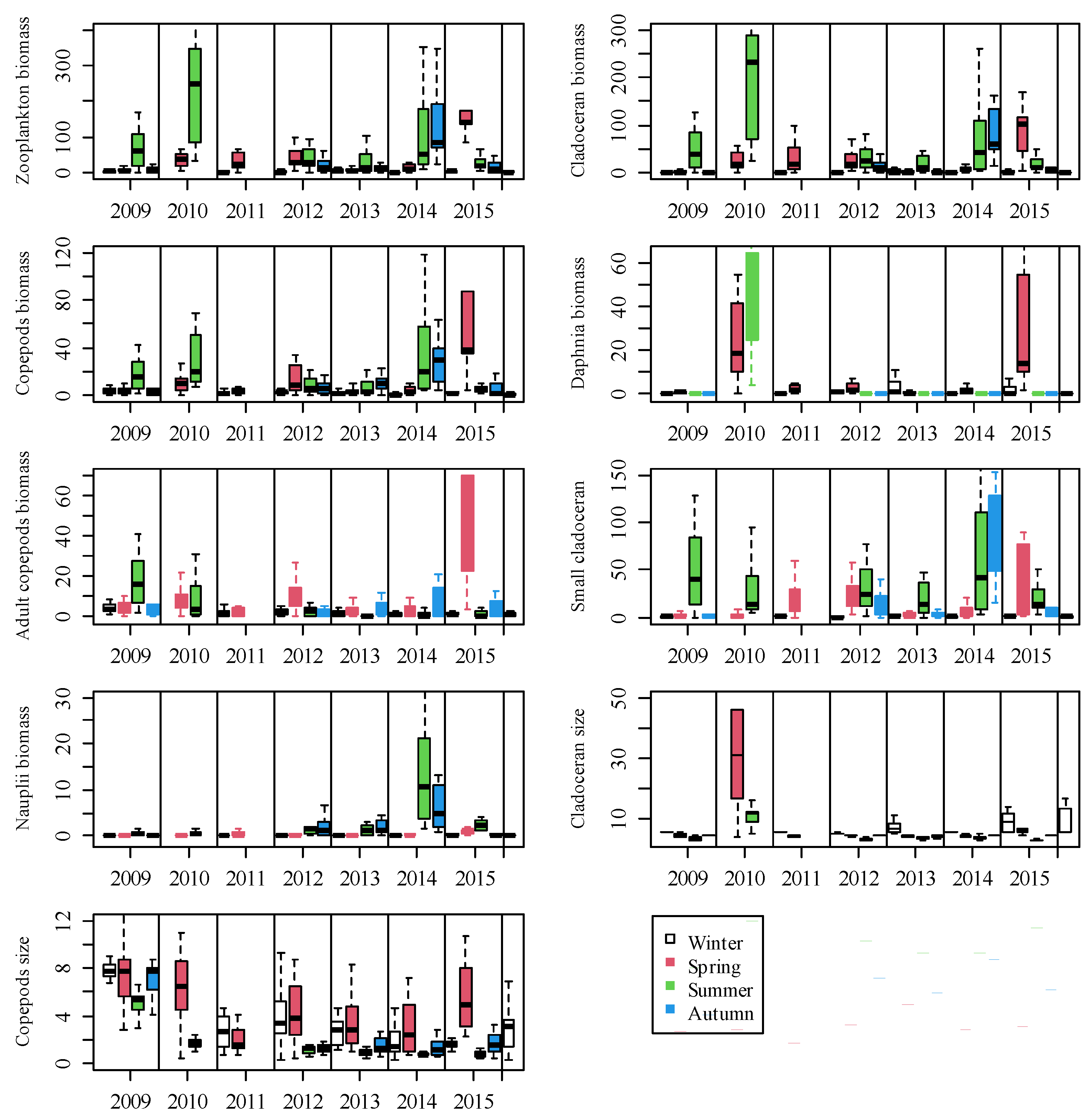
3.2. Zooplankton Biomass and Body Size
3.3. Zooplankton Community Structure
3.4. Direct and Indirect Effects of Water Temperature and Water Level
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

References
- Schoell, K.; Kiss, A.; Dinka, M.; Berczik, A. Flood-Pulse Effects on Zooplankton Assemblages in a River-Floodplain System (Gemenc Floodplain of the Danube, Hungary). Int. Rev. Hydrobiol. 2012, 97, 41–54. [Google Scholar] [CrossRef]
- Junk, W.J.; Bayley, P.B.; Sparks, R.E. The flood pulse concept in river-floodplain systems. Can. Spec. Publ. Fish. Aquat. Sci. 1989, 106, 110–127. [Google Scholar]
- Lansac-Tôha, F.; Bonecker, C.; Velho, L.; Simões, N.; Dias, J.; Alves, G.; Takahashi, E. Biodiversity of zooplankton communities in the Upper Paraná River floodplain: Interannual variation from long-term studies. Braz. J. Biol. 2009, 69, 539–549. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guida, R.J.; Remo, J.W.F.; Secchi, S. Tradeoffs of strategically reconnecting rivers to their floodplains: The case of the Lower Illinois River (USA). Sci. Total Environ. 2016, 572, 43–55. [Google Scholar] [CrossRef]
- Sommer, U.; Adrian, R.; De Senerpont Domis, L.; Elser, J.J.; Gaedke, U.; Ibelings, B.; Jeppesen, E.; Lürling, M.; Molinero, J.C.; Mooij, W.M. Beyond the Plankton Ecology Group (PEG) model: Mechanisms driving plankton succession. Ann. Rev. Ecol. Evol. Syst. 2012, 43, 429–448. [Google Scholar] [CrossRef]
- Mendes Colares, M.A.; Bonecker, C.C.; Simoes, N.R.; Alves, G.M.; Lansac-Toha, F.A. Structure of the zooplankton communities in macrophytes stand of a Neotropical floodplain (the Parana River, Brazil). Int. Rev. Hydrobiol. 2013, 98, 89–103. [Google Scholar] [CrossRef]
- Davidson, N.L., Jr.; Kelso, W.E.; Rutherford, D.A. Relationships between environmental variables and the abundance of cladocerans and copepods in the Atchafalaya River Basin. Hydrobiologia 1998, 379, 175–181. [Google Scholar] [CrossRef]
- Kiss, A.; Agoston-Szabo, E.; Dinka, M.; Schoell, K.; Berczik, A. Microcrustacean (Cladocera, Copepoda, Ostracoda) Diversity in Three Side Arms in the Gemenc Floodplain (Danube River, Hungary) in Different Hydrological Situations. Acta Zool. Bulg. 2014, 7, 135–141. [Google Scholar]
- Baranyi, C.; Hein, T.; Holarek, C.; Keckeis, S.; Schiemer, F. Zooplankton biomass and community structure in a Danube River floodplain system: Effects of hydrology. Freshw. Biol. 2002, 47, 473–482. [Google Scholar] [CrossRef]
- Lemke, M.J.; Paver, S.F.; Dungey, K.E.; Velho, L.F.M.; Kent, A.D.; Rodrigues, L.C.; Kellerhals, D.M.; Randle, M.R. Diversity and succession of pelagic microorganism communities in a newly restored Illinois River floodplain lake. Hydrobiologia 2017, 804, 35–58. [Google Scholar] [CrossRef] [Green Version]
- Wahl, D.H.; Goodrich, J.; Nannini, M.A.; Dettmers, J.M.; Soluk, D.A. Exploring riverine zooplankton in three habitats of the Illinois River ecosystem: Where do they come from? Limnol. Oceanogr. 2008, 53, 2583–2593. [Google Scholar] [CrossRef] [Green Version]
- G-Tóth, L.; Parpala, L.; Balogh, C.; Tàtrai, I.; Baranyai, E. Zooplankton community response to enhanced turbulence generated by water-level decrease in Lake Balaton, the largest shallow lake in Central Europe. Limnol. Oceanogr. 2011, 56, 2211–2222. [Google Scholar] [CrossRef]
- Levine, S.N.; Zehrer, R.F.; Burns, C.W. Impact of resuspended sediment on zooplankton feeding in Lake Waihola, New Zealand. Freshw. Biol. 2005, 50, 1515–1536. [Google Scholar] [CrossRef]
- Simoes, N.R.; Lansac-Toha, F.A.; Velho, L.F.M.; Bonecker, C.C. Intra and inter-annual structure of zooplankton communities in floodplain lakes: A long-term ecological research study. Rev. Biol. Trop. 2012, 60, 1819–1836. [Google Scholar] [CrossRef]
- Magalhaes Braghin, L.d.S.; Dias, J.D.; Simoes, N.R.; Bonecker, C.C. Food availability, depth, and turbidity drive zooplankton functional diversity over time in a Neotropical floodplain. Aquat. Sci. 2020, 83, 10. [Google Scholar] [CrossRef]
- Nogueira, M.G. Zooplankton composition, dominance and abundance as indicators of environmental compartmentalization in Jurumirim Reservoir (Paranapanema River), Sao Paulo, Brazil. Hydrobiologia 2001, 455, 1–18. [Google Scholar] [CrossRef]
- Bini, L.M.; Tundisi, J.G.; Matsumura-Tundisi, T.; Matheus, C.E. Spatial variation of zooplankton groups in a tropical reservoir (Broa Reservoir, São Paulo State-Brazil). Hydrobiologia 1997, 357, 89–98. [Google Scholar] [CrossRef]
- Bernot, R.J.; Dodds, W.K.; Quist, M.C.; Guy, C.S. Spatial and temporal variability of zooplankton in a great plains reservoir. Hydrobiologia 2004, 525, 101–112. [Google Scholar] [CrossRef] [Green Version]
- Brett, M.T.; Kainz, M.J.; Taipale, S.J.; Seshan, H. Phytoplankton, not allochthonous carbon, sustains herbivorous zooplankton production. Proc. Natl. Acad. Sci. USA 2009, 106, 21197–21201. [Google Scholar] [CrossRef] [Green Version]
- Cole, J.J.; Carpenter, S.R.; Kitchell, J.; Pace, M.L.; Solomon, C.T.; Weidel, B. Strong evidence for terrestrial support of zooplankton in small lakes based on stable isotopes of carbon, nitrogen, and hydrogen. Proc. Natl. Acad. Sci. USA 2011, 108, 1975–1980. [Google Scholar] [CrossRef] [Green Version]
- Hein, T.; Baranyi, C.; Herndl, G.J.; Wanek, W.; Schiemer, F. Allochthonous and autochthonous particulate organic matter in floodplains of the River Danube: The importance of hydrological connectivity. Freshw. Biol. 2003, 48, 220–232. [Google Scholar] [CrossRef]
- Stolte, W.; Lindström, T.; Granéli, E. Enhancement of poorly edible phytoplankton by allochthonous dissolved organic material: A modelling study. Aquat. Microb. Rcol. 2007, 46, 263–272. [Google Scholar] [CrossRef] [Green Version]
- Matthews, B.; Mazumder, A. Habitat specialization and the exploitation of allochthonous carbon by zooplankton. Ecology 2006, 87, 2800–2812. [Google Scholar] [CrossRef]
- Karlsson, J.; Jonsson, A.; Meili, M.; Jansson, M. Control of zooplankton dependence on allochthonous organic carbon in humic and clear-water lakes in northern Sweden. Limnol. Oceanogr. 2003, 48, 269–276. [Google Scholar] [CrossRef]
- Ranta, E.; Bengtsson, J.; McManus, J. Growth, size and shape of Daphnia longispina, D.magna and D.pulex. Ann. Zool. Fennici 1993, 30, 299–311. [Google Scholar]
- Abrusan, G. Filamentous cyanobacteria, temperature and Daphnia growth: The role of fluid mechanics. Oecologia 2004, 141, 395–401. [Google Scholar] [CrossRef]
- Jeppesen, E.; Meerhoff, M.; Davidson, T.A.; Trolle, D.; Sondergaard, M.; Lauridsen, T.L.; Beklioglu, M.; Brucet, S.; Volta, P.; Gonzalez-Bergonzoni, I.; et al. Climate change impacts on lakes: An integrated ecological perspective based on a multi-faceted approach, with special focus on shallow lakes. J. Limnol. 2014, 73, 88–111. [Google Scholar] [CrossRef] [Green Version]
- Sommer, U.; Gliwicz, Z.M.; Lampert, W.; Duncan, A. The PEG-model of seasonal succession of planktonic events in fresh waters. Arch. Hydrobiol. 1986, 106, 433–471. [Google Scholar]
- Straile, D. Zooplankton biomass dynamics in oligotrophic versus eutrophic conditions: A test of the PEG model. Freshw. Biol. 2015, 60, 174–183. [Google Scholar] [CrossRef] [Green Version]
- Sommer, U.; Lewandowska, A. Climate change and the phytoplankton spring bloom: Warming and overwintering zooplankton have similar effects on phytoplankton. Glob. Chang. Biol. 2011, 17, 154–162. [Google Scholar] [CrossRef]
- Domis, L.N.D.S.; Elser, J.J.; Gsell, A.S.; Huszar, V.L.M.; Ibelings, B.W.; Jeppesen, E.; Kosten, S.; Mooij, W.M.; Roland, F.; Sommer, U.; et al. Plankton dynamics under different climatic conditions in space and time. Freshw. Biol. 2013, 58, 463–482. [Google Scholar] [CrossRef]
- Benke, A.C.; Chaubey, I.; Ward, G.M.; Dunn, E.L. Flood pulse dynamics of an unregulated river floodplain in the southeastern US coastal plain. Ecology 2000, 81, 2730–2741. [Google Scholar] [CrossRef]
- Zhao, K.; Song, K.; Pan, Y.; Wang, L.; Da, L.; Wang, Q. Metacommunity structure of zooplankton in river networks: Roles of environmental and spatial factors. Ecol. Indic. 2017, 73, 96–104. [Google Scholar] [CrossRef]
- Thomaz, S.M.; Bini, L.M.; Bozelli, R.L. Floods increase similarity among aquatic habitats in river-floodplain systems. Hydrobiologia 2007, 579, 1–13. [Google Scholar] [CrossRef]
- Zhang, Q.; Dong, X.; Chen, Y.; Yang, X.; Xu, M.; Davidson, T.A.; Jeppesen, E. Hydrological alterations as the major driver on environmental change in a floodplain Lake Poyang (China): Evidence from monitoring and sediment records. J. Great Lakes Res. 2018, 44, 377–387. [Google Scholar] [CrossRef]
- Li, B.; Yang, G.; Wan, R.; Hoermann, G. Dynamic water quality evaluation based on fuzzy matter-element model and functional data analysis, a case study in Poyang Lake. Environ. Sci. Pollut. Res. 2017, 24, 19138–19148. [Google Scholar] [CrossRef]
- Dai, L.; Wang, L.; Li, L.; Liang, T.; Zhang, Y.; Ma, C.; Xing, B. Multivariate geostatistical analysis and source identification of heavy metals in the sediment of Poyang Lake in China. Sci. Total Environ. 2018, 621, 1433–1444. [Google Scholar] [CrossRef]
- Shao, W.W.; Chen, X.D.; Zhou, Z.H.; Liu, J.H.; Yan, Z.Q.; Chen, S.L.; Wang, J.H. Analysis of river runoff in the Poyang Lake Basin of China: Long-term changes and influencing factors. Hydrol. Sci. J. 2017, 62, 575–587. [Google Scholar] [CrossRef]
- Li, Y.L.; Zhang, Q.; Werner, A.D.; Yao, J.; Ye, X.C. The influence of river-to-lake backflow on the hydrodynamics of a large floodplain lake system (Poyang Lake, China). Hydrol. Process. 2017, 31, 117–132. [Google Scholar] [CrossRef]
- Liu, B.; Liu, J.; Jeppesen, E.; Chen, Y.; Liu, X.; Zhang, W. Horizontal distribution of pelagic crustacean zooplankton biomass and body size in contrasting habitat types in Lake Poyang, China. Environ. Sci. Pollut. Res. 2019, 26, 2270–2280. [Google Scholar] [CrossRef] [PubMed]
- APHA. Standard Methods for the Examination of Water and Wastewater; American Public Health Association (APHA): Washington, DC, USA, 2005. [Google Scholar]
- Wang, J.; Pan, F.; Soininen, J.; Heino, J.; Shen, J. Nutrient enrichment modifies temperature-biodiversity relationships in large-scale field experiments. Nat. Commun. 2016, 7, 13960. [Google Scholar] [CrossRef] [Green Version]
- Feitosa, I.B.; Huszar, V.L.M.; Domingues, C.D.; Appel, E.; Paranhos, R.; Almeida, R.M.; Branco, C.W.C.; Bastos, W.R.; Sarmento, H. Plankton community interactions in an Amazonian floodplain lake, from bacteria to zooplankton. Hydrobiologia 2019, 831, 55–70. [Google Scholar] [CrossRef]
- Moustaka-Gouni, M.; Michaloudi, E.; Sommer, U. Modifying the PEG model for Mediterranean lakes—No biological winter and strong fish predation. Freshw. Biol. 2014, 59, 1136–1144. [Google Scholar] [CrossRef] [Green Version]
- Taipale, S.J.; Galloway, A.W.; Aalto, S.L.; Kahilainen, K.K.; Strandberg, U.; Kankaala, P. Terrestrial carbohydrates support freshwater zooplankton during phytoplankton deficiency. Sci. Rep. 2016, 6, 30897. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taipale, S.; Kankaala, P.; Jones, R.I. Contributions of different organic carbon sources to Daphnia in the pelagic foodweb of a small polyhumic lake: Results from mesocosm (DIC)-C-13-additions. Ecosystems 2007, 10, 757–772. [Google Scholar] [CrossRef]
- Grey, J.; Jones, R.I.; Sleep, D. Stable isotope analysis of the origins of zooplankton carbon in lakes of differing trophic state. Oecologia 2000, 123, 232–240. [Google Scholar] [CrossRef] [PubMed]
- Karlsson, J.; Bergstrom, A.K.; Bystrom, P.; Gudasz, C.; Rodriguez, P.; Hein, C. Terrestrial organic matter input suppresses biomass production in lake ecosystems. Ecology 2015, 96, 2870–2876. [Google Scholar] [CrossRef] [Green Version]
- Berger, S.A.; Diehl, S.; Stibor, H.; Sebastian, P.; Scherz, A. Separating effects of climatic drivers and biotic feedbacks on seasonal plankton dynamics: No sign of trophic mismatch. Freshw. Biol. 2014, 59, 2204–2220. [Google Scholar] [CrossRef]
- Berggren, M.; Ziegler, S.E.; St-Gelais, N.F.; Beisner, B.E.; del Giorgio, P.A. Contrasting patterns of allochthony among three major groups of crustacean zooplankton in boreal and temperate lakes. Ecology 2014, 95, 1947–1959. [Google Scholar] [CrossRef]
- Zhang, H.; Yu, X.B.; Wang, Y.Y.; Xu, J. The importance of terrestrial carbon in supporting molluscs in the wetlands of Poyang Lake. Chin. J. Oceanol. Limnol. 2017, 35, 825–832. [Google Scholar] [CrossRef]
- Shurin, J.B.; Winder, M.; Adrian, R.; Keller, W.; Matthews, B.; Paterson, A.M.; Paterson, M.J.; Pinel-Alloul, B.; Rusak, J.A.; Yan, N.D. Environmental stability and lake zooplankton diversity—Contrasting effects of chemical and thermal variability. Ecol. Lett. 2010, 13, 453–463. [Google Scholar] [CrossRef]
- Liu, X.; Li, Y.-l.; Liu, B.-G.; Qian, K.-M.; Chen, Y.-W.; Gao, J.-F. Cyanobacteria in the complex river-connected Poyang Lake: Horizontal distribution and transport. Hydrobiologia 2016, 768, 95–110. [Google Scholar] [CrossRef]
- Thorp, J.H.; Mantovani, S. Zooplankton of turbid and hydrologically dynamic prairie rivers. Freshw. Biol. 2005, 50, 1474–1491. [Google Scholar] [CrossRef]
- Thorp, J.H.; Black, A.R.; Haag, K.H.; Wehr, J.D. Zooplankton Assemblages in the Ohio River: Seasonal, tributary, and navigation dam effects. Can. J. Fish. Aquat. Sci. 1994, 51, 1634–1643. [Google Scholar] [CrossRef]
- Jeppesen, E.; Jensen, J.P.; Sondergaard, M.; Lauridsen, T.; Pedersen, L.J.; Jensen, L. Top-down control in freshwater lakes: The role of nutrient state, submerged macrophytes and water depth. Hydrobiologia 1997, 342, 151–164. [Google Scholar] [CrossRef]
- Ślusarczyk, M. Impact of Fish Predation on a Small-Bodied Cladoceran: Limitation or Stimulation? Developments in Hydrobiology Book Series; Kluwer Academic Publishers: Brussels, Belgium, 1997; pp. 215–221. [Google Scholar]
- Brooks, J.L.; Dodson, S.I. Predation, body size, and composition of plankton. Science 1965, 150, 28–35. [Google Scholar] [CrossRef] [PubMed]
- Luecke, C.; Vanni, M.J.; Magnuson, J.J.; Kitchell, J.F.; Jacobson, P.T. Seasonal regulation of Daphnia populations by planktivorous fish: Implications for the spring clear-water phase. Limnol. Oceanogr. 1990, 35, 1718–1733. [Google Scholar] [CrossRef]
- Feniova, I.; Dawidowicz, P.; Gladyshev, M.I.; Kostrzewska-Szlakowska, I.; Rzepecki, M.; Razlutskij, V.; Sushchik, N.N.; Majsak, N.; Dzialowski, A.R. Experimental effects of large-bodied Daphnia, fish and zebra mussels on cladoceran community and size structure. J. Plankton Res. 2015, 37, 611–625. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| As in PEG Model | Differences from PEG Model and Additions | |
|---|---|---|
| Winter |
|
|
| Spring |
|
|
| Summer |
|
|
| Autumn | Not clear |
|
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, B.; Wu, J.; Hu, Y.; Wang, G.; Chen, Y. Seven Years Study of the Seasonal Dynamics of Zooplankton Communities in a Large Subtropical Floodplain Ecosystem: A Test of the PEG Model. Int. J. Environ. Res. Public Health 2022, 19, 956. https://doi.org/10.3390/ijerph19020956
Liu B, Wu J, Hu Y, Wang G, Chen Y. Seven Years Study of the Seasonal Dynamics of Zooplankton Communities in a Large Subtropical Floodplain Ecosystem: A Test of the PEG Model. International Journal of Environmental Research and Public Health. 2022; 19(2):956. https://doi.org/10.3390/ijerph19020956
Chicago/Turabian StyleLiu, Baogui, Jiayi Wu, Yang Hu, Guoxiang Wang, and Yuwei Chen. 2022. "Seven Years Study of the Seasonal Dynamics of Zooplankton Communities in a Large Subtropical Floodplain Ecosystem: A Test of the PEG Model" International Journal of Environmental Research and Public Health 19, no. 2: 956. https://doi.org/10.3390/ijerph19020956
APA StyleLiu, B., Wu, J., Hu, Y., Wang, G., & Chen, Y. (2022). Seven Years Study of the Seasonal Dynamics of Zooplankton Communities in a Large Subtropical Floodplain Ecosystem: A Test of the PEG Model. International Journal of Environmental Research and Public Health, 19(2), 956. https://doi.org/10.3390/ijerph19020956