
Biology, Systematics, Microbiome, Pathogen Transmission and Control of Chiggers (Acari: Trombiculidae, Leeuwenhoekiidae) with Emphasis on the United States



Abstract
1. Introduction
2. Chigger Biology
2.1. Life Cycle
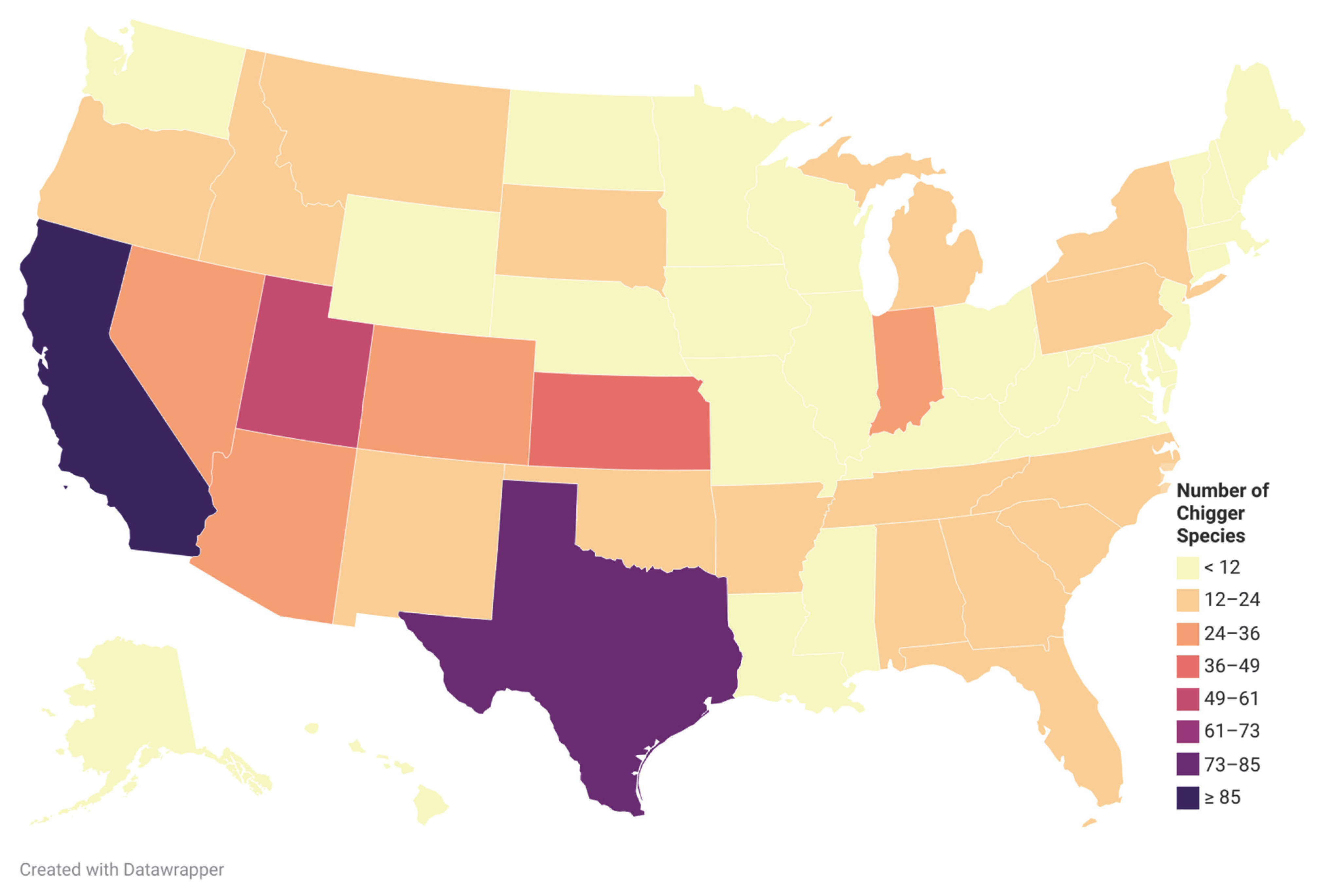
2.2. Hosts and Habitat
2.3. Collecting Methods
2.3.1. Trapping Animals
2.3.2. Plate Method
2.3.3. Berlese Funnel
2.3.4. Light Trapping
2.4. Chigger Lab Rearing
3. Systematics Status
3.1. Taxonomy
3.2. Identification
4. Chigger Microbiome
5. Do Chiggers Transmit Rickettsia and Orientia in the US?
5.1. Summary of Zoonotic Bacteria and Viruses Transmitted by Chiggers
5.2. Role of Endosymbionts in Orientia Infection
6. Possible Misdiagnosis of Tick-Borne Diseases and Allergies
7. Chigger Control
8. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Weitzel, T.; Silva-de la Fuente, M.C.; Martínez-Valdebenito, C.; Stekolnikov, A.A.; Pérez, C.; Pérez, R.; Vial, C.; Abarca, K.; Acosta-Jamett, G. Novel vector of scrub typhus in sub-Antarctic Chile: Evidence from human exposure. Clin. Infect. Dis. 2022, 74, 1862–1865. [Google Scholar] [CrossRef]
- Watt, G.; Parola, P. Scrub typhus and tropical rickettsioses. Curr. Opin. Infect. Dis. 2003, 16, 429–436. [Google Scholar] [CrossRef] [PubMed]
- Philip, C.B. Tsutsugamushi disease (scrub typhus) in World War II. J. Parasitol. 1948, 34, 169–191. [Google Scholar] [CrossRef] [PubMed]
- Kunz, S.E.; Price, M.A.; Graham, O.H. Biology and economic importance of the chigger Neoschongastia americana on Turkeys. J. Econ. Entomol. 1969, 62, 872–875. [Google Scholar] [CrossRef]
- Nielsen, D.H.; Robbins, R.G.; Rueda, L.M. Annotated World Checklist of the Trombiculidae and Leeuwenhoekiidae (1758–2021) (Acari: Trombiculoidea), with notes on nomenclature, taxonomy, and distribution. Zootaxa 2021, 4967, 1–243. [Google Scholar] [CrossRef]
- Pritt, S.; Cohen, K.; Sedlacek, H. Chapter 15—Parasitic diseases. In The Laboratory Rabbit, Guinea Pig, Hamster, and Other Rodents; Suckow, M.A., Stevens, K.A., Wilson, R.P., Eds.; American College of Laboratory Animal Medicine; Academic Press: Boston, MA, USA, 2012; pp. 415–446. ISBN 978-0-12-380920-9. [Google Scholar]
- Rashmir-Raven, A.M. Chapter 18—Disorders of the skin. In Equine Internal Medicine, 4th ed.; Reed, S.M., Bayly, W.M., Sellon, D.C., Saunders, W.B., Eds.; Elsevier Inc.: Amsterdam, Netherlands, 2018; pp. 1159–1216. ISBN 978-0-323-44329-6. [Google Scholar]
- Lee, I.-Y.; Yoon, S.-S.; Lee, W.-J.; Sin, H.-K.; Lee, W.-K. Observation of stylostome formation in the striped-field mouse (Apodemus agrarius Pallas) skin by chigger feeding. Korean J. Soil Zool. 2006, 11, 1–6. [Google Scholar]
- Shatrov, A.B. Stylostome formation in Trombiculid mites (Acariformes: Trombiculidae). Exp. Appl. Acarol. 2009, 49, 261. [Google Scholar] [CrossRef]
- Neal, T.J.; Barnett, H.C. The life cycle of the scrub typhus chigger mite Trombicula akamushi. Ann. Entomol. Soc. Am. 1961, 54, 196–203. [Google Scholar] [CrossRef]
- Everett, R.E.; Price, M.A.; Kunz, S.E. Biology of the chigger Neoschöngastia americana (Acarina: Trombiculidae). Ann. Entomol. Soc. Am. 1973, 66, 429–435. [Google Scholar] [CrossRef]
- Santibáñez, P.; Palomar, A.M.; Portillo, A.; Santibáñez, S.; Oteo, J.A. The role of chiggers as human pathogens. In An Overview of Tropical Diseases; Samie, A., Ed.; I-Tech Education and Publishing: Vienna, Austria, 2015; ISBN 978-953-51-2224-1. [Google Scholar]
- Lv, Y.; Guo, X.-G.; Jin, D.-C. Research progress on Leptotrombidium deliense. Korean J. Parasitol. 2018, 56, 313–324. [Google Scholar] [CrossRef]
- Shatrov, A.B.; Kudryashova, N.I. Taxonomy, life cycles and the origin of parasitism in Trombiculid mites. In Micromammals and Macroparasites: From Evolutionary Ecology to Management; Morand, S., Krasnov, B.R., Poulin, R., Eds.; Springer Japan: Tokyo, Japan, 2006; pp. 119–140. ISBN 978-4-431-36025-4. [Google Scholar]
- Wharton, G.W.; Fuller, H.S. A Manual of the Chiggers: A manual of the chiggers: The biology, classification, distribution and importance to man of the larvae of the family Trombiculidae (Acarina). Mem. Entomol. Soc. Wash. 1952, 4, 185. [Google Scholar]
- Lipovsky, L.J.; Byers, G.W.; Kardos, E.H. Spermatophores: The mode of insemination of chiggers (Acarina: Trombiculidae). J. Parasitol. 1957, 43, 256–262. [Google Scholar] [CrossRef] [PubMed]
- Walters, B.L.; Whitaker, J.O., Jr.; Gikas, N.S.; Wrenn, W.J. Host and Distribution Lists of Chiggers (Trombiculidae and Leeuwenhoekiidae), of North American Wild Vertebrates North of Mexico. Faculty Publications from the Harold W. Manter Laboratory of Parasitology No. 697, 2011. Available online: https://digitalcommons.unl.edu/parasitologyfacpubs/697/ (accessed on 19 May 2022).
- Sasa, M. Biology of chiggers. Annu. Rev. Entomol. 1961, 6, 221–244. [Google Scholar] [CrossRef]
- Zhan, Y.-Z.; Guo, X.-G.; Speakman, J.R.; Zuo, X.-H.; Wu, D.; Wang, Q.-H.; Yang, Z.-H. Abundances and host relationships of chigger mites in Yunnan Province, China. Med. Vet. Entomol. 2013, 27, 194–202. [Google Scholar] [CrossRef]
- Clopton, R.E.; Gold, R.E. Distribution and seasonal and diurnal activity patterns of Eutrombicula alfreddugesi (Acari: Trombiculidae) in a forest edge ecosystem. J. Med. Entomol. 1993, 30, 47–53. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Wang, D.; Chen, X. Trombiculid Mites of China: Studies on Vector and Pathogen of Tsutsugamushi Disease; Guangdong Science and Technology Publishing: Guangzhou, China, 1997; pp. 1–157. [Google Scholar]
- Durden, L.A.; Hu, R.; Oliver, J.H., Jr.; Cilek, J.E. Rodent ectoparasites from two locations in Northwestern Florida. J. Vector Ecol. 2000, 25, 222–228. [Google Scholar] [PubMed]
- Ponnusamy, L.; Garshong, R.; McLean, B.S.; Wasserberg, G.; Durden, L.A.; Crossley, D.; Apperson, C.S.; Roe, R.M. Rickettsia felis and other Rickettsia species in chigger mites collected from wild rodents in North Carolina, USA. Microorganisms 2022, 10, 1342. [Google Scholar] [CrossRef] [PubMed]
- Krantz, G.W.; Walter, D.E. A Manual of Acarology, 3rd ed.; Texas Tech University Press: Lubbock, TX, USA, 2009; ISBN 0-89672-620-7. [Google Scholar]
- Upham, R.W., Jr.; Hubert, A.A.; Phang, O.W.; Mat, Y.B.; Rapmund, G. Distribution of Leptotrombidium (Leptotrombidium) arenicola (Acarina: Trombiculidae) on the ground in West Malaysia. J. Med. Entomol. 1971, 8, 401–406. [Google Scholar] [CrossRef] [PubMed]
- Rohani, I.B.; Cromroy, H.L. Taxonomy and distribution of chiggers (Acarina: Trombiculidae) in Northcentral Florida. Fla. Entomol. 1979, 62, 363–376. [Google Scholar] [CrossRef]
- Nadchatram, M.; Dohany, A.L. Leptotrombidium (Leptotrombidium) umbricola, new species, a probable vector of scrub typhus in Peninsular Malaysia. Jpn. J. Med. Sci. Biol. 1980, 33, 277–282. [Google Scholar] [CrossRef]
- Tanskul, P.; Gingrich, J.B. Two new species of Leptotrombidium (Acari: Actinedida: Trombiculidae), probable vectors of scrub typhus in Thailand. J. Med. Entomol. 1986, 23, 661–665. [Google Scholar] [CrossRef] [PubMed]
- Jones, B.M. A method for studying the distribution and bionomics of Trombiculid mites (Acarina: Trombidiidae). Parasitology 1950, 40, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Nadchatram, M. A technique for rearing trombiculid mites (Acarina) developed in a tropical laboratory. J. Med. Entomol. 1968, 5, 465–469. [Google Scholar] [CrossRef] [PubMed]
- Frances, S.P.; Khlaimanee, N. Laboratory tests of arthropod repellents against Leptotrombidium Deliense noninfected and infected with Rickettsia tsutsugamushi and noninfected L. fletcheri (Acari: Trombiculidae). J. Med. Entomol. 1996, 33, 232–235. [Google Scholar] [CrossRef]
- Michener, C.D. A method of rearing chigger mites (Acarina, Trombiculinae). Am. J. Trop. Med. 1946, 26, 251–256. [Google Scholar] [CrossRef] [PubMed]
- Neal, T.J.; Lipvsky, L.J. Techniques for rearing the chigger mite, Trombicula Akamushi (Brumpt). J. Econ. Entomol. 1959, 52, 824–826. [Google Scholar] [CrossRef]
- Jenkins, D.W. A laboratory method of rearing chiggers affecting man (Acarina: Trombiculidae). Ann. Entomol. Soc. Am. 1947, 40, 56–68. [Google Scholar] [CrossRef]
- Audy, J.R.; Nadchatram, M. A Method of rearing individual Trombiculid mites in the field. Nature 1954, 174, 1021–1022. [Google Scholar] [CrossRef]
- Goff, M.L.; Loomis, R.B.; Welbourn, W.C.; Wrenn, W.J. A glossary of chigger terminology (Acari: Trombiculidae). J. Med. Entomol. 1982, 19, 221–238. [Google Scholar] [CrossRef]
- Fernandes, S.; Kulkarni, S.M. Studies on the Trombiculid Mite Fauna of India; Zoological Survey of India: Kolkata, India, 2003; ISBN 81-85874-99-9. [Google Scholar]
- Chaisiri, K.; Stekolnikov, A.A.; Makepeace, B.L.; Morand, S. A revised checklist of chigger mites (Acari: Trombiculidae) from Thailand, with the description of three new species. J. Med. Entomol. 2016, 53, 321–342. [Google Scholar] [CrossRef]
- Stekolnikov, A.A. A checklist of chigger mites (Acariformes: Trombiculidae) of Southeast Asia. Zootaxa 2021, 4913, 1–163. [Google Scholar] [CrossRef] [PubMed]
- Reeves, W.K.; Durden, L.A.; Wrenn, W.J. Ectoparasitic chiggers (Acari: Trombiculidae, Leeuwenhoekiidae), lice (Phthiraptera), and Hemiptera (Cimicidae and Reduviidae) from South Carolina, USA. Zootaxa 2004, 647, 1–20. [Google Scholar] [CrossRef]
- Wen, T.-H. New taxa and tentative rearrangement of Walchiidae Stat. n. with remarks on Trombiculoidea Nec Welbourn, 1991 (Acari: Acariformes) 1. Syst. Appl. Acarol. 1999, 4, 165–178. [Google Scholar] [CrossRef]
- Wen, T. Classification of the sand-mite Family Walchiidae (Acariformes: Trombiculoidea). Chin. J. Parasitol. Parasit. Dis. 2004, 22, 113–118. [Google Scholar]
- Zhang, Z.-Q.; Fan, Q.-H.; Pesic, V.; Smit, H.; Bochkov, A.V.; Khaustov, A.A.; Baker, A.; Wohltmann, A.; Wen, T.; Amrine, J.W.; et al. Order Trombidiformes Reuter, 1909. In Animal Biodiversity: An Outline of Higher-Level Classification and Survey of Taxonomic Richness; Magnolia Press: Auckland, New Zealand, 2011; pp. 129–138. ISBN 978-1-86977-850-7. [Google Scholar]
- Antonovskaia, A.A. Using DNA Markers in Studies of Chigger Mites (Acariformes, Trombiculidae). Entmol. Rev. 2018, 98, 1351–1368. [Google Scholar] [CrossRef]
- Crossley, D.A.; Clement, M.J. A new species of chigger (Acari: Trombiculidae) from Rafinesque’s Big-Eared bat (Chiroptera: Vespertilionidae) in Georgia, USA. J. Entomol. Sci 2015, 50, 248–251. [Google Scholar] [CrossRef]
- Bennett, S.G.; Crossley, D.A.; Durden, L.A.; Goff, M.L. Eutrombicula cinnabaris (Ewing, 1920) (Acari:Trombiculidae) is the common pest chigger mite of the Eastern United States. Entomol. Sci. 2014, 49, 413–414. [Google Scholar] [CrossRef]
- Loomis, R.B.; Wrenn, W.J. Systematics of the Pest chigger genus Eutrombicula (Acari: Trombiculidae). In Proceedings of the VI International Congress of Acarology; Griffiths, D.A., Bowman, C.E., Eds.; Ellis Horwood: Chichester, UK, 1984. [Google Scholar]
- Ewing, H.E. Notes on Trombiculid mites with descriptions of Walchiinae n. subf., Speotrombicula n. g., and Eutrombicula defecta n. sp. J. Parasitol. 1946, 32, 435–440. [Google Scholar] [CrossRef]
- Fuller, H.S. The mite larvae of the family Trombiculidae in the Oudemans collection: Taxonomy and medical importance. Zool. Verh. 1952, 18, 1–261. [Google Scholar]
- Wright, S.M.; Wikel, S.K.; Wrenn, W.J. Host immune responsiveness to the chigger, Eutrombicula cinnabaris. Ann. Trop. Med. Parasitol. 1988, 82, 283–293. [Google Scholar] [CrossRef]
- Tuegel, M.A.; Wrenn, W.J. Sexual dimorphism in morphology and development of the pest chigger, Eutrombicula cinnabaris (Acari: Trombiculidae). Int. J. Acarol. 1998, 24, 199–211. [Google Scholar] [CrossRef]
- Mertins, J.W.; Torrence, S.M.; Sterner, M.C. Chiggers recently infesting Spea spp. in Texas, Usa, were Eutrombicula alfreddugesi, not Hannemania sp. J. Wildl. Dis. 2011, 47, 612–617. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Garvin, S.D.; Noden, B.H.; Dillwith, J.W.; Fox, S.F.; Payton, M.E.; Barker, R.W. Sylvatic infestation of Oklahoma reptiles with immature Ixodes scapularis (Acari: Ixodidae). J. Med. Entomol. 2015, 52, 873–878. [Google Scholar] [CrossRef] [PubMed]
- Bakkegard, K.A.; Patton, A.H.; Ray, C.H. Chigger mites (Hannemania Cf. Dunni) infest Northern Slimy Salamanders (Plethodon glutinosus) in Alabama, USA. Herpetol. Conserv. Biol. 2019, 14, 578–586. [Google Scholar]
- McAllister, C.T.; Durden, L.A.; Greiman, S.E. Euschoengastia pipistrelli (Acari: Trombiculidae) from American perimyotis, Perimyotis subflavus (Chiroptera: Vespertilionidae): Novel stereoscopic and scanning electron microscopy. J. Parasitol. 2021, 107, 125–128. [Google Scholar] [CrossRef] [PubMed]
- Durden, L.A.; Ellis, B.A.; Banks, C.W.; Crowe, J.D.; Oliver, J.H. Ectoparasites of gray squirrels in two different habitats and screening of selected ectoparasites for Bartonellae. J. Parasitol. 2004, 90, 485–489. [Google Scholar] [CrossRef]
- McAllister, C.T.; Bursey, C.R.; Robison, H.W.; Connior, M.B. Parasites of the Ozark zig-zag salamander, Plethodon angusticlavius (Caudata: Plethodontidae), from Northern Arkansas. Comp. Parasitol. 2013, 80, 69–79. [Google Scholar] [CrossRef]
- McAllister, C.T.; Bursey, C.R.; Connior, M.B.; Trauth, S.E.; Durden, L.A. New host and geographic distribution records for helminth and arthropod parasites of the southern toad, Anaxyrus terrestris (Anura: Bufonidae), from Florida. Southwest. Nat. 2015, 14, 641–649. [Google Scholar] [CrossRef]
- Anderson, L.E. Hoyer’s solution as a rapid permanent mounting medium for bryophytes. Bryologist 1954, 57, 242–244. [Google Scholar] [CrossRef]
- Swan, D.C. Berlese’s Fluid: Remarks upon its preparation and use as a mounting medium. Bull. Entomol. Res. 1936, 27, 389–391. [Google Scholar] [CrossRef]
- Stekolnikov, A.A. A new genus and species of chigger mites (Acariformes: Trombiculidae) from Madagascar. Int. J. Acarol. 2021, 47, 301–307. [Google Scholar] [CrossRef]
- Philip Samuel, P.; Govindarajan, R.; Krishnamoorthi, R.; Venkatesh, A. A rapid protocol for clearing, staining, and mounting of Arthropoda: Trombiculidae, Pediculidae and Pulicidae. North-West. J. Zool. 2021, 17, e201104. [Google Scholar]
- Nadchatram, M.; Dohany, A.L. A pictorial key to the subfamilies, genera and subgenera of Southeast Asian chiggers (Acari, Prostigmata, Trombiculidae). Bull. Inst. Med. Res. Malays. 1974, 16, 1–67. [Google Scholar]
- Brennan, J.M.; Goff, M.L. Keys to the genera of chiggers of the Western Hemisphere (Acarina: Trombiculidae). J. Parasitol. 1977, 63, 554–566. [Google Scholar] [CrossRef]
- Vercammen-Grandjean, P.H.; Langston, R.L. The Chigger Mites of the World:(Acarina): Trombiculidae & Leeuwenhoekiidae; George Williams Hooper Foundation, University of California: San Francisco, CA, USA, 1976; Volume 3. [Google Scholar]
- Stekolnikov, A.A. Leptotrombidium (Acari: Trombiculidae) of the world. Zootaxa 2013, 3728, 1–173. [Google Scholar] [CrossRef]
- Pearce, R.D.; O’Shea, T.J. Ectoparasites in an urban population of big brown bats (Eptesicus fuscus) in Colorado. J. Parasitol. 2007, 93, 518–530. [Google Scholar] [CrossRef]
- Alghamdi, S. The Vector Biology and Microbiome of Parasitic Mites and Other Ectoparasites of Rodents. Ph.D. Thesis, The University of Liverpool (United Kingdom), Liverpool, UK, 2019. [Google Scholar]
- Pepato, A.R.; Klimov, P.B. Origin and higher-level diversification of acariform mites—Evidence from nuclear ribosomal genes, extensive taxon sampling, and secondary structure alignment. BMC Evol. Biol. 2015, 15, 178. [Google Scholar] [CrossRef]
- Klimov, P.B.; OConnor, B.M.; Chetverikov, P.E.; Bolton, S.J.; Pepato, A.R.; Mortazavi, A.L.; Tolstikov, A.V.; Bauchan, G.R.; Ochoa, R. Comprehensive phylogeny of acariform mites (Acariformes) provides insights on the origin of the four-legged mites (Eriophyoidea), a long branch. Mol. Phylogenet. Evol. 2018, 119, 105–117. [Google Scholar] [CrossRef]
- Kumlert, R.; Chaisiri, K.; Anantatat, T.; Stekolnikov, A.A.; Morand, S.; Prasartvit, A.; Makepeace, B.L.; Sungvornyothin, S.; Paris, D.H. Autofluorescence microscopy for paired-matched morphological and molecular identification of individual chigger mites (Acari: Trombiculidae), the Vectors of Scrub Typhus. PLoS ONE 2018, 13, e0193163. [Google Scholar] [CrossRef]
- Dong, X.; Chaisiri, K.; Xia, D.; Armstrong, S.D.; Fang, Y.; Donnelly, M.J.; Kadowaki, T.; McGarry, J.W.; Darby, A.C.; Makepeace, B.L. Genomes of trombidid mites reveal novel predicted allergens and laterally transferred genes associated with secondary metabolism. GigaScience 2018, 7, giy127. [Google Scholar] [CrossRef]
- Zhou, Q.; Wang, Z.-X.; Tao, J.-M.; Qin, J.-P.; Lu, J.-P.; Lin, R.-Q.; Wang, L.-M.; Weng, Y.-B.; Tan, Z.-J. Characterization of Neoschoengastia gallinarum from subtropical China by rDNA and identification of two genotypes based on mitochondrial Cox1. Parasitol. Res. 2020, 119, 3339–3345. [Google Scholar] [CrossRef] [PubMed]
- Park, S.-W.; Ha, N.-Y.; Ryu, B.; Bang, J.H.; Song, H.; Kim, Y.; Kim, G.; Oh, M.; Cho, N.-H.; Lee, J. Urbanization of scrub typhus disease in South Korea. PLOS Negl. Trop. Dis. 2015, 9, e0003814. [Google Scholar] [CrossRef] [PubMed]
- Shao, R.; Mitani, H.; Barker, S.C.; Takahashi, M.; Fukunaga, M. Novel mitochondrial gene content and gene arrangement indicate illegitimate inter-mtDNA recombination in the chigger mite, Leptotrombidium pallidum. J. Mol. Evol. 2005, 60, 764–773. [Google Scholar] [CrossRef] [PubMed]
- Shao, R.; Barker, S.C.; Mitani, H.; Takahashi, M.; Fukunaga, M. Molecular mechanisms for the variation of mitochondrial gene content and gene arrangement among chigger mites of the genus Leptotrombidium (Acari: Acariformes). J. Mol. Evol. 2006, 63, 251–261. [Google Scholar] [CrossRef]
- Young, M.R.; Behan-Pelletier, V.M.; Hebert, P.D.N. Revealing the hyperdiverse mite fauna of subarctic Canada through DNA Barcoding. PLoS ONE 2012, 7, e48755. [Google Scholar] [CrossRef]
- de Castro Jacinavicius, F.; Bassini-Silva, R.; Muñoz-Leal, S.; Welbourn, C.; Ochoa, R.; Labruna, M.B.; Barros-Battesti, D.M. Molecular detection of Rickettsia genus in chigger mites (Trombidiformes: Trombiculidae) collected on small mammals in Southeastern Brazilian. Rev. Bras. Parasitol. Vet. 2019, 28, 563–568. [Google Scholar] [CrossRef] [PubMed]
- Ja, M.-R.; Dm, B.-B. A new species of pit mite (Trombidiformes: Harpirhynchidae) from the South American rattlesnake (Viperidae): Morphological and molecular analysis. Entomol. Ornithol. Herpetol. 2017, 6, 8. [Google Scholar]
- Moniuszko, H.; Zaleśny, G.; Mąkol, J. Host-associated differences in morphometric traits of parasitic larvae Hirsutiella zachvatkini (Actinotrichida: Trombiculidae). Exp. Appl. Acarol. 2015, 67, 123–133. [Google Scholar] [CrossRef]
- Moniuszko, H.; Felska, M.; Mąkol, J. Evidence for co-invasion events: Different chigger species (Actinotrichida, Trombidioidea: Trombiculidae) share a host. Exp. Appl. Acarol. 2018, 76, 29–39. [Google Scholar] [CrossRef]
- Söller, R.; Wohltmann, A.; Witte, H.; Blohm, D. Phylogenetic relationships within terrestrial mites (Acari: Prostigmata, Parasitengona) inferred from comparative DNA sequence analysis of the mitochondrial cytochrome oxidase subunit I gene. Mol. Phylogenet. Evol. 2001, 18, 47–53. [Google Scholar] [CrossRef]
- Kiene, F.; Andriatsitohaina, B.; Ramsay, M.S.; Rakotondramanana, H.; Rakotondravony, R.; Radespiel, U.; Strube, C. Forest edges affect ectoparasite infestation patterns of small mammalian hosts in fragmented forests in Madagascar. Int. J. Parasitol. 2020, 50, 299–313. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.I.; Shim, S.K.; Song, B.G.; Choi, E.N.; Hwang, K.J.; Park, M.Y.; Park, C.; Shin, E.-H. Detection of Orientia tsutsugamushi, the causative agent of scrub typhus, in a novel mite species, Eushoengastia koreaensis, in Korea. Vector Borne Zoonotic Dis. 2011, 11, 209–214. [Google Scholar] [CrossRef] [PubMed]
- Dohany, A.L.; Shirai, A.; Robinson, D.M.; Ram, S.; Huxsoll, D.L. Identification and antigenic typing of Rickettsia tsutsugamushi in naturally infected chiggers (Acarina: Trombiculidae) by direct immunofluorescence. Am. J. Trop. Med. Hyg. 1978, 27, 1261–1264. [Google Scholar] [CrossRef] [PubMed]
- Ree, H.I.; Lee, I.Y.; Cho, M.K. Determination of the vector species of Tsutsugamushi disease in Korea. Korean J. Parasitol. 1991, 29, 87–92. [Google Scholar] [CrossRef] [PubMed]
- Ponnusamy, L.; Willcox, A.C.; Roe, R.M.; Davidson, S.A.; Linsuwanon, P.; Schuster, A.L.; Richards, A.L.; Meshnick, S.R.; Apperson, C.S. Bacterial microbiome of the chigger mite Leptotrombidium imphalum varies by life stage and infection with the scrub typhus pathogen Orientia tsutsugamushi. PLoS ONE 2018, 13, e0208327. [Google Scholar] [CrossRef] [PubMed]
- Chaisiri, K.; Gill, A.C.; Stekolnikov, A.A.; Hinjoy, S.; McGarry, J.W.; Darby, A.C.; Morand, S.; Makepeace, B.L. Ecological and microbiological diversity of chigger mites, including vectors of scrub typhus, on small mammals across stratified habitats in Thailand. Anim. Microbiome 2019, 1, 18. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.; Richards, A.L. Scrub typhus: No longer restricted to the Tsutsugamushi Triangle. Trop. Med. Infect. Dis. 2018, 3, 11. [Google Scholar] [CrossRef] [PubMed]
- Elliott, I.; Pearson, I.; Dahal, P.; Thomas, N.V.; Roberts, T.; Newton, P.N. Scrub typhus ecology: A systematic review of Orientia in vectors and hosts. Parasit. Vectors 2019, 12, 513. [Google Scholar] [CrossRef]
- Xu, G.; Walker, D.H.; Jupiter, D.; Melby, P.C.; Arcari, C.M. A review of the global epidemiology of scrub typhus. PLOS Negl. Trop. Dis. 2017, 11, e0006062. [Google Scholar] [CrossRef]
- Phasomkusolsil, S.; Tanskul, P.; Ratanatham, S.; Watcharapichat, P.; Phulsuksombati, D.; Frances, S.P.; Lerdthusnee, K.; Linthicum, K.J. Transstadial and transovarial transmission of Orientia tsutsugamushi in Leptotrombidium imphalum and Leptotrombidium chiangraiensis (Acari: Trombiculidae). J. Med. Entomol. 2009, 46, 1442–1445. [Google Scholar] [CrossRef]
- Paris, D.H.; Shelite, T.R.; Day, N.P.; Walker, D.H. Unresolved problems related to scrub typhus: A seriously neglected life-threatening disease. Am. J. Trop. Med. Hyg. 2013, 89, 301–307. [Google Scholar] [CrossRef] [PubMed]
- Kelly, D.J.; Fuerst, P.A.; Ching, W.-M.; Richards, A.L. Scrub typhus: The geographic distribution of phenotypic and denotypic variants of Orientia tsutsugamushi. Clin. Infect. Dis. 2009, 48, S203–S230. [Google Scholar] [CrossRef] [PubMed]
- Izzard, L.; Fuller, A.; Blacksell, S.D.; Paris, D.H.; Richards, A.L.; Aukkanit, N.; Nguyen, C.; Jiang, J.; Fenwick, S.; Day, N.P. Isolation of a novel Orientia species (O. chuto sp. nov.) from a patient infected in Dubai. J. Clin. Microbiol. 2010, 48, 4404–4409. [Google Scholar] [CrossRef] [PubMed]
- Balcells, M.E.; Rabagliati, R.; García, P.; Poggi, H.; Oddó, D.; Concha, M.; Abarca, K.; Jiang, J.; Kelly, D.J.; Richards, A.L. Endemic scrub typhus–like illness, Chile. Emerg. Infect. Dis. 2011, 17, 1659. [Google Scholar] [CrossRef]
- Weitzel, T.; Dittrich, S.; López, J.; Phuklia, W.; Martinez-Valdebenito, C.; Velásquez, K.; Blacksell, S.D.; Paris, D.H.; Abarca, K. Endemic scrub typhus in South America. N. Engl. J. Med. 2016, 375, 954–961. [Google Scholar] [CrossRef]
- Kocher, C.; Jiang, J.; Morrison, A.C.; Castillo, R.; Leguia, M.; Loyola, S.; Ampuero, J.S.; Cespedes, M.; Halsey, E.S.; Bausch, D.G. Serologic evidence of scrub typhus in the Peruvian Amazon. Emerg. Infect. Dis. 2017, 23, 1389. [Google Scholar] [CrossRef]
- Cosson, J.F.; Galan, M.; Bard, E.; Razzauti, M.; Bernard, M.; Morand, S.; Brouat, C.; Dalecky, A.; Bâ, K.; Charbonnel, N.; et al. Detection of Orientia sp. DNA in rodents from Asia, West Africa and Europe. Parasit. Vectors 2015, 8, 172. [Google Scholar] [CrossRef]
- Masakhwe, C.; Linsuwanon, P.; Kimita, G.; Mutai, B.; Leepitakrat, S.; Yalwala, S.; Abuom, D.; Auysawasi, N.; Gilbreath, T.; Wanja, E.; et al. Identification and characterization of Orientia chuto in Trombiculid chigger mites collected from wild rodents in Kenya. J. Clin. Microbiol. 2018, 56, e01124-18. [Google Scholar] [CrossRef]
- Tilak, R.; Kunwar, R.; Wankhade, U.B.; Tilak, V.W. Emergence of Schoengastiella ligula as the vector of scrub typhus outbreak in Darjeeling: Has Leptotrombidium deliense been replaced? Indian J. Public Health 2011, 55, 92. [Google Scholar] [CrossRef]
- Peters, C.J.; Simpson, G.L.; Levy, H. Spectrum of Hantavirus Infection: Hemorrhagic Fever with Renal Syndrome and Hantavirus Pulmonary Syndrome. Annu. Rev. Med. 1999, 50, 531–545. [Google Scholar] [CrossRef]
- CDC Hantavirus Pulmonary Syndrome (HPS)—Hantavirus. Available online: https://www.cdc.gov/hantavirus/hps/index.html (accessed on 11 April 2022).
- Wu, G.; Zhang, Y.; Guo, H.; Jiang, K.; Zhang, J.; Gan, Y. The role of Leptotrombidium scutellare in the transmission of human diseases. Chin. Med. J. 1996, 109, 670–673. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Tesh, R.B. The role of mites in the transmission and maintenance of Hantaan Virus (Hantavirus: Bunyaviridae). J. Infect. Dis. 2014, 210, 1693–1699. [Google Scholar] [CrossRef] [PubMed]
- Houck, M.A.; Qin, H.; Roberts, H.R. Hantavirus transmission: Potential role of ectoparasites. Vector Borne Zoonotic Dis. 2001, 1, 75–79. [Google Scholar] [CrossRef] [PubMed]
- Kosoy, M.; Morway, C.; Sheff, K.W.; Bai, Y.; Colborn, J.; Chalcraft, L.; Dowell, S.F.; Peruski, L.F.; Maloney, S.A.; Baggett, H. Bartonella Tamiae Sp. Nov., a newly recognized pathogen isolated from three human patients from Thailand. J. Clin. Microbiol. 2008, 46, 772–775. [Google Scholar] [CrossRef] [PubMed]
- Kabeya, H.; Colborn, J.M.; Bai, Y.; Lerdthusnee, K.; Richardson, J.H.; Maruyama, S.; Kosoy, M.Y. Detection of Bartonella tamiae DNA in ectoparasites from rodents in Thailand and their sequence similarity with bacterial cultures from Thai patients. Vector Borne Zoonotic Dis. 2010, 10, 429–434. [Google Scholar] [CrossRef] [PubMed]
- Tsui, P.-Y.; Tsai, K.-H.; Weng, M.-H.; Hung, Y.-W.; Liu, Y.-T.; Hu, K.-Y.; Lien, J.-C.; Lin, P.-R.; Shaio, M.-F.; Wang, H.-C. Molecular detection and characterization of spotted fever group Rickettsiae in Taiwan. Am. J. Trop. Med. Hyg. 2007, 77, 883–890. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.-J.; Lee, E.-M.; Park, J.-M.; Lee, K.-M.; Han, S.-H.; Kim, J.-K.; Lee, S.-H.; Song, H.-J.; Choi, M.-S.; Kim, I.-S.; et al. Molecular detection of various Rickettsiae in mites (Acari: Trombiculidae) in Southern Jeolla Province, Korea. Microbiol. Immunol. 2007, 51, 307–312. [Google Scholar] [CrossRef]
- Huang, Y.; Zhao, L.; Zhang, Z.; Liu, M.; Xue, Z.; Ma, D.; Sun, X.; Sun, Y.; Zhou, C.; Qin, X.; et al. Detection of a novel Rickettsia from Leptotrombidium scutellare mites (Acari: Trombiculidae) from Shandong of China. J. Med. Entomol. 2017, 54, 544–549. [Google Scholar] [CrossRef]
- Ogawa, M.; Takahashi, M.; Matsutani, M.; Takada, N.; Noda, S.; Saijo, M. Obligate intracellular bacteria diversity in unfed Leptotrombidium scutellare larvae highlights novel bacterial endosymbionts of mites. Microbiol. Immunol. 2020, 64, 1–9. [Google Scholar] [CrossRef]
- Kampen, H.; Schöler, A.; Metzen, M.; Oehme, R.; Hartelt, K.; Kimmig, P.; Maier, W.A. Neotrombicula autumnalis (Acari, Trombiculidae) as a vector for Borrelia burgdorferi sensu lato? Exp. Appl. Acarol. 2004, 33, 93–102. [Google Scholar] [CrossRef]
- Literak, I.; Stekolnikov, A.A.; Sychra, O.; Dubska, L.; Taragelova, V. Larvae of chigger mites Neotrombicula spp. (Acari: Trombiculidae) exhibited Borrelia but no Anaplasma infections: A Field Study Including Birds from the Czech Carpathians as Hosts of Chiggers. Exp. Appl. Acarol. 2008, 44, 307–314. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, M.; Urakami, H.; Yoshida, Y.; Furuya, Y.; Misumi, H.; Hori, E.; Kawamura, A.; Tanaka, H. Occurrence of high ratio of males after introduction of minocycline in a colony of Leptotrombidium fletcheri infected with Orientia tsutsugamushi. Eur. J. Epidemiol. 1997, 13, 19–23. [Google Scholar] [CrossRef] [PubMed]
- Duron, O.; Bouchon, D.; Boutin, S.; Bellamy, L.; Zhou, L.; Engelstädter, J.; Hurst, G.D. The diversity of reproductive parasites among arthropods: Wolbachia do not walk alone. BMC Biol. 2008, 6, 27. [Google Scholar] [CrossRef] [PubMed]
- Phasomkusolsil, S.; Tanskul, P.; Ratanatham, S.; Watcharapichat, P.; Phulsuksombati, D.; Frances, S.P.; Lerdthusnee, K.; Linthicum, K.J. Influence of Orientia tsutsugamushi infection on the developmental biology of Leptotrombidium imphalum and Leptotrombidium chiangraiensis (Acari: Trombiculidae). J. Med. Entomol. 2012, 49, 1270–1275. [Google Scholar] [CrossRef] [PubMed]
- Rodino, K.G.; Theel, E.S.; Pritt, B.S. Tick-borne diseases in the United States. Clin. Chem. 2020, 66, 537–548. [Google Scholar] [CrossRef] [PubMed]
- Robinson, M.T.; Satjanadumrong, J.; Hughes, T.; Stenos, J.; Blacksell, S.D. Diagnosis of spotted fever group Rickettsia infections: The Asian perspective. Epidemiol. Infect. 2019, 147, e286. [Google Scholar] [CrossRef]
- Zhou, C.; Woods, P.; Abouzeid, A.; Brooks, M.N. Case Report: Rocky Mountain Spotted Fever with adrenalectomy and a gard-to-find tick. Am. J. Case Rep. 2022, 23, e934505-1–e934505-6. [Google Scholar] [CrossRef]
- Shapiro, M.R.; Fritz, C.L.; Tait, K.; Paddock, C.D.; Nicholson, W.L.; Abramowicz, K.F.; Karpathy, S.E.; Dasch, G.A.; Sumner, J.W.; Adem, P.V.; et al. Rickettsia 364D: A newly recognized cause of eschar-Associated Illness in California. Clin. Infect. Dis. 2010, 50, 541–548. [Google Scholar] [CrossRef]
- Kuo, C.-C.; Lee, P.-L.; Wang, H.-C. Molecular identification of Rickettsia spp. in chigger mites in Taiwan. Med. Vet. Entomol. 2021, 223–229. [Google Scholar] [CrossRef]
- Zhioua, E.; Rodhain, F.; Binet, P.h.; Perez-Eid, C. Prevalence of antibodies to Borrelia burgdorferi in forestry workers of Ile de France, France. Eur. J. Epidemiol. 1997, 13, 959–962. [Google Scholar] [CrossRef]
- Crispell, G.; Commins, S.P.; Archer-Hartman, S.A.; Choudhary, S.; Dharmarajan, G.; Azadi, P.; Karim, S. Discovery of Alpha-Gal-containing antigens in North American tick species believed to induce red meat allergy. Front. Immunol. 2019, 10, 1056. [Google Scholar] [CrossRef] [PubMed]
- Commins, S.P.; Platts-Mills, T.A.E. Tick bites and red meat allergy. Curr. Opin. Allergy Clin. Immunol. 2013, 13, 354–359. [Google Scholar] [CrossRef] [PubMed]
- Stoltz, L.P.; Cristiano, L.M.; Dowling, A.P.G.; Wilson, J.M.; Platts-Mills, T.A.E.; Traister, R.S. Could chiggers be contributing to the prevalence of galactose-alpha-1,3-galactose sensitization and mammalian meat allergy? J. Allergy Clin. Immunol. Pract. 2019, 7, 664–666. [Google Scholar] [CrossRef]
- Chiggers. Available online: https://hortnews.extension.iastate.edu/chiggers (accessed on 7 April 2022).
- Houseman, R.M. Chiggers. Available online: https://extension.missouri.edu/publications/g7398 (accessed on 13 April 2022).
- Anderson, R.R.; Oi, F.M. Chiggers. Available online: https://ssl.acesag.auburn.edu/pubs/docs/A/ANR-1109/ANR-1109-archive.pdf (accessed on 13 April 2022).
- Moore, G.C.; Merchant, M.E. Chiggers. Available online: https://agrilifeextension.tamu.edu/library/landscaping/chiggers/ (accessed on 7 April 2022).
- Ewing, H.E.; Hartzell, A. The chigger-mites affecting man and domestic animals. J. Econ. Entomol. 1918, 11, 256–264. [Google Scholar] [CrossRef][Green Version]
- Ewing, H.E. Our only common North American chigger, its distribution and nomenclature. J. Agric. Res. 1923, 26, 401–403. [Google Scholar]
- Ewing, H.E. A contribution to our knowledge of the taxonomy of chiggers, including the descriptions of a new genus, six new species and a new variety. Am. J. Trop. Med. 1925, 5, 251–265. [Google Scholar] [CrossRef]
- Ewing, H.E. The adult of our common North American chigger. Proceedings of the Biological Society of Washington 1925. [Google Scholar]
- Ewing, H.E. Birds as hosts for the common chigger. Am. Nat. 1929, 63, 94–96. [Google Scholar] [CrossRef]
- Ewing, H.E. A catalogue of the Trombiculinae, or chigger mites, of the new world, with new genera and species and a key to the genera. Proc. United States Natl. Mus. 1931, 80. [Google Scholar] [CrossRef]
- Ewing, H.E. A key to the genera of chiggers (mite larvae of the subfamily Trombiculinae) with descriptions of new genera and species. J. Wash. Acad. Sci. 1938, 28, 288–295. [Google Scholar]
- Ewing, H.E. Remarks on the taxonomy of some American chiggers (Trombiculinae), including the descriptions of New Genera and species. J. Parasitol. 1942, 28, 485–493. [Google Scholar] [CrossRef]
- Ewing, H.E. The American chiggers (larvae of the Trombiculinae) of the genus Acariscus, new genus. Proc. Entomol. Soc. Wash. 1943, 45, 57–66. [Google Scholar]
- Brennan, J.M. A new genus and species of chigger, Chatia setosa (Trombiculidae, Acarina) from Northwestern United States. J. Parasitol. 1946, 32, 132–135. [Google Scholar] [CrossRef] [PubMed]
- Doetschman, W.; Furman, D. A tropical chigger, Eutrombicula batatas (Linn) attacking man in California. Am. J. Trop. Med. 1949, 29, 605–608. [Google Scholar] [CrossRef] [PubMed]
- Jenkins, D.W. Trombiculid mites affecting man. III. Trombicula (Eutrombicula) splendens Ewing in North America. J. Parasitol. 1949, 35, 201–204. [Google Scholar] [CrossRef]
- Brennan, J.M.; Wharton, G.W. Studies on North American chiggers. No. 3. the subgenus Neotrombicula. Am. Midl. Nat. 1950, 44, 153–197. [Google Scholar] [CrossRef]
- Lipovsky, L.J.; Loomis, R.B. A new chigger mite (genus Euschöngastia) from the Central United States. J. Parasitol. 1954, 40, 407–410. [Google Scholar] [CrossRef]
- Crossley, D.A. A new species of chigger mite from Colorado (Acarina, Trombiculidae). J. Parasitol. 1955, 41, 289–291. [Google Scholar] [CrossRef]
- Hyland, K.E. A new species of chigger mite, Hannemania hegeneri (Acarina; Trombiculidae). J. Parasitol. 1956, 42, 176–179. [Google Scholar] [CrossRef]
- Brennan, J.M.; White, J.S. New records and descriptions of chiggers (Acarina: Trombiculidae) on Bats in Alabama. J. Parasitol. 1960, 46, 346–350. [Google Scholar] [CrossRef]
- Johnston, D.E.; DeGiusti, D.L. Ecological and systematic notes on some chigger mites of the genera Gahrliepia, Trombicula, and Euschongastia (Acarina: Trombiculidae) in Michigan and Ontario. J. Parasitol. 1961, 47, 11–12. [Google Scholar] [CrossRef]
- Powder, W.A.; Loomis, R.B. A new species and new records of chiggers (Acarina, Trombiculidae) from reptiles of Southern California. J. Parasitol. 1962, 48, 204–208. [Google Scholar] [CrossRef] [PubMed]
- Loomis, R.B. The discovery of chiggers (Acarina, Trombiculidae) in the nasal passages of cricetid rodents from California, with descriptions of two new species. J. Parasitol. 1963, 49, 330–333. [Google Scholar] [CrossRef]
- Brennan, J.M. Five new chiggers from Southwestern United States (Acarina: Trombiculidae). J. Parasitol. 1965, 51, 108–113. [Google Scholar] [CrossRef] [PubMed]
- Brennan, J.M. New records of chiggers (Acarina: Trombiculidae) from Baja California and islands of the gulf of California. J. Parasitol. 1966, 52, 772–775. [Google Scholar] [CrossRef]
- Johnston, D.E.; Wharton, G.W.; Stoll, R.J. On the occurrence of Fonsecia palmella Brennan and Loomis (Acari: Trombiculidae), a chigger mite of reptiles, in Ohio. J. Parasitol. 1966, 52, 966. [Google Scholar] [CrossRef]
- Webb, J.P., Jr.; Loomis, R.B. A new subgenus of intranasal chiggers of the genus Microtrombicula from North America and Korea. J. Med. Entomol. 1970, 7, 655–663. [Google Scholar] [CrossRef] [PubMed]
- Crossley, D.A.; Atyeo, W.T. A new species of chigger, Blankaartia pauli (Acarina: Trombiculidae), from the Southeastern United States. J. Med. Entomol. 1972, 9, 253–254. [Google Scholar] [CrossRef]
- Everett, R.E.; Price, M.A.; Kunz, S.E. New host records of the chigger Neoschoengastia americana from Texas (Acarina: Trombiculidae). J. Med. Entomol. 1972, 9, 109–110. [Google Scholar] [CrossRef] [PubMed]
- Easton, E.R. Ectoparasites in two diverse habitats in Western Oregon. J. Med. Entomol. 1975, 12, 295–298. [Google Scholar] [CrossRef]
- Goff, M.L.; Judd, F.W. The first record of a chigger from the Texas tortoise, Gopherus berlandieri. Southwest. Nat. 1981, 26, 83–84. [Google Scholar] [CrossRef]
- Pomeroy, L.V.; Loomis, R.B. A new genus of Trombiculine chiggers (Acari: Trombiculidae) from Western North America. J. Med. Entomol. 1984, 21, 268–273. [Google Scholar] [CrossRef]
- Ludwig, D.F.; Crossley, D.A., Jr.; Hayes, M.J.; Mallow, D.; Wicht, M.C., Jr. Pest chigger, Eutrombicula alfreddugesi infestation of small mammals in piedmont habitats of Georgia (Acarina: Trombiculidae). J. Entomol. Sci. 1985, 20, 1–8. [Google Scholar] [CrossRef]
- Durden, L.A.; Wilson, N. Parasitic and phoretic arthropods of sylvatic and commensal white-footed mice (Peromyscus leucopus) in Central Tennessee, with notes on Lyme Disease. J. Parasitol. 1991, 77, 219–223. [Google Scholar] [CrossRef] [PubMed]
- Wilson, N.A.; Telford, S.R., Jr.; Forrester, D.J. Ectoparasites of a population of urban gray squirrels in northern florida. J. Med. Entomol. 1991, 28, 461–464. [Google Scholar] [CrossRef] [PubMed]
- Durden, L.A. Parasitic arthropods of sympatric meadow voles and white-footed mice at Fort Detrick, Maryland. J. Med. Entomol. 1992, 29, 761–766. [Google Scholar] [CrossRef] [PubMed]
- Goldberg, S.R.; Bursey, C.R. Duration of attachment of the chigger, Eutrombicula lipovskyana (Trombiculidae) in mite pockets of Yarrow’s Spiny Lizard, Sceloporus jarrovii (Phrynosomatidae) from Arizona. J. Wildl. Dis. 1993, 29, 142–144. [Google Scholar] [CrossRef] [PubMed]
- Anthony, C.D.; Mendelson, J.R.; Simons, R.R. Differential parasitism by sex on Plethodontid Salamanders and histological evidence for structural damage to the nasolabial groove. Am. Midl. Nat. 1994, 132, 302–307. [Google Scholar] [CrossRef]
- Goldberg, S.R.; Bursey, C.R. Prevalence of ectoparasite infestation in neonate Yarrow’s spiny lizards, Sceloporus jarrovii (Phrynosomatidae), from Arizona. Great Basin Nat. 1994, 54, 189–190. [Google Scholar] [CrossRef]
- Pung, O.J.; Durden, L.A.; Banks, C.W.; Jones, D.N. Ectoparasites of opossums and raccoons in Southeastern Georgia. J. Med. Entomol. 1994, 31, 915–919. [Google Scholar] [CrossRef]
- Forrester, D.J.; Mclaughlin, G.S.; Telford, S.R., Jr.; Foster, G.W.; Mccown, J.W. Ectoparasites (Acari, Mallophaga, Anoplura, Diptera) of white-tailed deer, Odocoileus virginianus from Southern Florida. J. Med. Entomol. 1996, 33, 96–101. [Google Scholar] [CrossRef]
- Durden, L.A.; Banks, C.W.; Clark, K.L.; Belbey, B.V.; Oliver, J.H. Ectoparasite fauna of the eastern woodrat, Neotoma floridana: composition, origin, and comparison with ectoparasite faunas of Western woodrat species. J. Parasitol. 1997, 83, 374–381. [Google Scholar] [CrossRef] [PubMed]
- Goff, M.L.; Mckown, R.D. The genus Hexidionis (Acari: Trombiculidae) with the description of a new species from Texas. J. Med. Entomol. 1997, 34, 438–440. [Google Scholar] [CrossRef] [PubMed]
- Little, S.E.; Carmichael, K.P.; Rakich, P.M. Trombidiosis-induced dermatitis in white-tailed deer (Odocoileus virginianus). Vet. Pathol. 1997, 34, 350–352. [Google Scholar] [CrossRef] [PubMed]
- Spalding, M.G.; Wrenn, W.J.; Schwikert, S.T.; Schmidt, J.A. Dermatitis in young Florida Sandhill cranes (Grus canadensis Pratensis) due to infestation by the chigger, Blankaartia sinnamaryi. J. Parasitol. 1997, 83, 768–771. [Google Scholar] [CrossRef]
- Pung, O.J.; Durden, L.A.; Patrick, M.J.; Conyers, T.; Mitchell, L.R. Ectoparasites and gastrointestinal helminths of Southern flying squirrels in Southeast Georgia. J. Parasitol. 2000, 86, 1051–1055. [Google Scholar] [CrossRef]
- Sladky, K.K.; Norton, T.M.; Loomis, M.R. Trombiculid mites (Hannemania sp.) in canyon tree frogs (Hyla arenicolor). J. Zoo Wildl. Med. 2000, 31, 570–575. [Google Scholar] [CrossRef]
- Williams, C.K.; Davidson, W.R.; Lutz, R.S.; Applegate, R.D. Health status of northern bobwhite quail (Colinus virginianus) in Eastern Kansas. Avian Dis. 2000, 44, 953–956. [Google Scholar] [CrossRef]
- Cunningham, M.W.; Phillips, L.A.; Welbourn, C. Trombiculiasis in the Florida black bear. J. Wildl. Dis. 2001, 37, 634–639. [Google Scholar] [CrossRef][Green Version]
- Klukowski, M. Seasonal changes in abundance of host-seeking chiggers (Acari: Trombiculidae) and infestations on fence lizards, Sceloporus undulatus. J. Herpetol. 2004, 38, 141–144. [Google Scholar] [CrossRef]
- Malone, J.H.; Paredes-Leon, R. Characteristics of chigger mite (Hannemania sp.) parasitism on Eleutherodactylus marnockii (Amphibia: Leptodactylidae). Tex. J. Sci. 2005, 57, 345–358. [Google Scholar]
- Kurta, A.; Whitaker, J.O., Jr.; Wrenn, W.J.; Soto-centeno, J.A. Ectoparasitic assemblages on mormoopid bats (Chiroptera: Mormoopidae) from Puerto Rico. J. Med. Entomol. 2007, 44, 953–958. [Google Scholar] [CrossRef]
- Torrence, S.M.; Smith, L.M.; McMurry, S.T. Larval Hannemania sp. infestations of Spea spp. in the Southern High Plains, Texas, USA. J. Wildl. Dis. 2007, 43, 742–746. [Google Scholar] [CrossRef][Green Version]
- Nims, T.N.; Durden, L.A.; Chandler, C.R.; Pung, O.J. Parasitic and phoretic arthropods of the oldfield mouse (Peromyscus polionotus) from burned habitats with additional ectoparasite records from the eastern harvest mouse (Reithrodontomys humulis) and southern short-tailed shrew (Blarina carolinensis). Comp. Parasitol. 2008, 75, 102–106. [Google Scholar] [CrossRef]
- Westfall, M.C.; Cecala, K.K.; Price, S.J.; Dorcas, M.E. Patterns of Trombiculid mite (Hannemania dunni) parasitism among plethodontid salamanders in the western piedmont of North Carolina. J. Parasitol. 2008, 94, 631–634. [Google Scholar] [CrossRef]
- Bulté, G.; Plummer, A.C.; Thibaudeau, A.; Blouin-Demers, G. Infection of Yarrow’s spiny lizards (Sceloporus jarrovii) by chiggers and malaria in the Chiricahua Mountains, Arizona. Southwest. Nat. 2009, 54, 204–207. [Google Scholar] [CrossRef]
- Mertins, J.W.; Hanson, B.A.; Corn, J.L. Whartonacarus floridensis sp. nov. (Acari: Trombiculidae), with a taxonomic review and the first record of Whartonacarus chiggers in the continental United States. J. Med. Entomol. 2009, 46, 1260–1268. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Corn, J.L.; Mertins, J.W.; Hanson, B.; Snow, S. First reports of ectoparasites collected from wild-caught exotic reptiles in Florida. J. Med. Entomol. 2011, 48, 94–100. [Google Scholar] [CrossRef] [PubMed]
- McCoy, E.D.; Styga, J.M.; Rizkalla, C.E.; Mushinsky, H.R. Time since fire affects ectoparasite prevalence on lizards in the Florida scrub ecosystem. Fire Ecol. 2012, 8, 32–40. [Google Scholar] [CrossRef]
- Ott-Conn, C.N.; Woods, L.W.; Clifford, D.L.; Branston, T.; Foley, J. Histopathology and risk factors associated with Neotrombicula microti infestation in the endangered Amargosa vole (Microtus californicus scirpensis). J. Wildl. Dis. 2015, 51, 680–687. [Google Scholar] [CrossRef]
- McAllister, C.T.; Bursey, C.R.; Connior, M.B. New Host and Distributional records for parasites (Apicomplexa, Trematoda, Nematoda, Acanthocephala, Acarina) of Texas herpetofauna. Comp. Parasitol. 2017, 84, 42–50. [Google Scholar] [CrossRef]
- Milley, C.; Dryden, M.; Rosenkrantz, W.; Griffin, J.; Reeder, C. Comparison of parasitic mite retrieval methods in a population of community cats. J. Feline Med. Surg. 2017, 19, 657–664. [Google Scholar] [CrossRef] [PubMed]
- Harrity, E.J.; Conway, C.J. Novel ectoparasite infestation on Yuma Ridgway’s rails (Rallus obsoletus yumanensis). Wilson J. Ornithol. 2019, 131, 139–146. [Google Scholar] [CrossRef]
- Bassini-Silva, R.; Jacinavicius, F.D.C.; Welbourn, C.; Ochoa, R.; Barros-battesti, D.M. Synonymy of the genus Delmohius Brennan and Goff, 1978 with Carebareia Goff and Brennan, 1977 (Trombidiformes: Trombiculidae). Syst. Appl. Acarol. 2020, 25, 1188–1196. [Google Scholar] [CrossRef]
- Orton, R.W.; Kinsey, C.T.; McBrayer, L.D. Mite load predicts the quality of sexual color and locomotor performance in a sexually dichromatic lizard. Ecol. Evol. 2020, 10, 3152–3163. [Google Scholar] [CrossRef]
- McInnis, A.; Root, K.; Rizer, C.; Bradley, D. Diagnosing a crisis: The conundrum of alpha-gal. J. Nurse Pract. 2021, 17, 1081–1084. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Country (Continent) | Different Chigger Species in GenBank | % * | Peer Reviewed Publications | Chigger Species Reported | Target Genes * |
|---|---|---|---|---|---|
| Laos (Asia) | 11 | 30.56 | [71] | Ascoschoengastia indica | COI |
| Blankaartia acuscutellaris | COI | ||||
| Leptotrombidium deliense | 18S, COI, whole genome shotgun | ||||
| Microtrombicula chamlongi | COI | ||||
| Schoengastia kanhaensis | COI | ||||
| Walchia alpestris | COI | ||||
| Walchia ewingi ewingi | COI | ||||
| Walchia ewingi lupella | COI | ||||
| Walchia kritochaeta | COI | ||||
| Walchia micropelta | COI | ||||
| Walchia minuscuta | COI | ||||
| Thailand (Asia) | 1 | 2.78 | [72] | Leptotrombidium deliense | Mitochondrial (mt) DNA complete genome |
| China (Asia) | 1 | 2.78 | [73] | Neoschoengastia gallinarum | small subunit rRNA, large subunit rRNA, 5.8S, internal transcribed spacer 2, large subunit rRNA, COI |
| Korea (Asia) | 2 | 5.56 | [74] | Helenicula miyagawai | 18S |
| Leptotrombidium scutellare | mt DNA complete genome, COI, 18S, 5.8S | ||||
| Japan (Asia) | 1 | 2.78 | [75] | Leptotrombidium pallidum | COI, mt DNA complete genome, 5.8S, whole genome shotgun |
| Malaysia (Asia) | 1 | 2.78 | [76] | Leptotrombidium fletcheri | mt DNA complete genome |
| Canada (North America) | 1 | 2.78 | [77] | Neotrombicula microti | COI |
| US (North America) | 1 | 2.78 | [69,70] | Eutrombicula splendens | large subunit rRNA, small subunit rRNA, 28S, 18S, Ef1-α, Srp54, Hsp70, COI |
| Mexico (North America) | 1 | 2.78 | [70] | Acomatacarus arizonensis | 28S, 18S, COI, Ef1-α, Srp54, Hsp70 |
| Brazil (South America) | 12 | 32.33 | [78,79] | Herpetacarus(Arisocerus) hertigi | small subunit rRNA |
| Blankaartia sinnamaryi | small subunit rRNA | ||||
| Eutrombicula daemoni | small subunit rRNA | ||||
| Eutrombicula goeldii | small subunit rRNA | ||||
| Eutrombicula tinami | small subunit rRNA | ||||
| Fonsecia ewingi | 18S | ||||
| Hannemania hepatica | 18S | ||||
| Hannemania yungicola | 18S | ||||
| Quadraseta brasiliensis | small subunit rRNA, 18S | ||||
| Quadraseta pazca | small subunit rRNA | ||||
| Quadraseta trapezoides | small subunit rRNA | ||||
| Trombewingia bakeri | small subunit rRNA | ||||
| Poland (Europe) | 3 | 8.33 | [80,81] | Hirsutiella zachvatkini | COI |
| Miyatrombicula muris | COI | ||||
| Neotrombicula inopinata | COI | ||||
| Spain (Europe) | 1 | 2.78 | [82] | Morelacarus sp. | COI |
| Madagascar (Africa) | 1 | 2.78 | [83] | Schoutedenichia microcebi | small subunit rRNA |
| Scenarios | Methods of Avoiding Chigger Bites |
|---|---|
| When venturing into chigger habitat | Wear permethrin-treated clothing |
| Apply repellents like DEET | |
| Wear tightly woven long sleeves, long pants, and closed-toe shoes | |
| Tuck shirts into pants, tuck pants into socks | |
| Stay in the middle of the trail, avoid high bushes and tall grass | |
| Avoid sitting directly on the ground | |
| After you come inside | Remove clothing immediately and launder it in hot water |
| Wipe down shoes and boots | |
| Take a hot and soapy shower with a vigorous skin massage | |
| Yard | Try to keep small mammals out of the yard by putting up a fence and keeping trash can lids secured |
| keep turf short and vegetation trimmed |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, K.; Roe, R.M.; Ponnusamy, L. Biology, Systematics, Microbiome, Pathogen Transmission and Control of Chiggers (Acari: Trombiculidae, Leeuwenhoekiidae) with Emphasis on the United States. Int. J. Environ. Res. Public Health 2022, 19, 15147. https://doi.org/10.3390/ijerph192215147
Chen K, Roe RM, Ponnusamy L. Biology, Systematics, Microbiome, Pathogen Transmission and Control of Chiggers (Acari: Trombiculidae, Leeuwenhoekiidae) with Emphasis on the United States. International Journal of Environmental Research and Public Health. 2022; 19(22):15147. https://doi.org/10.3390/ijerph192215147
Chicago/Turabian StyleChen, Kaiying, R. Michael Roe, and Loganathan Ponnusamy. 2022. "Biology, Systematics, Microbiome, Pathogen Transmission and Control of Chiggers (Acari: Trombiculidae, Leeuwenhoekiidae) with Emphasis on the United States" International Journal of Environmental Research and Public Health 19, no. 22: 15147. https://doi.org/10.3390/ijerph192215147
APA StyleChen, K., Roe, R. M., & Ponnusamy, L. (2022). Biology, Systematics, Microbiome, Pathogen Transmission and Control of Chiggers (Acari: Trombiculidae, Leeuwenhoekiidae) with Emphasis on the United States. International Journal of Environmental Research and Public Health, 19(22), 15147. https://doi.org/10.3390/ijerph192215147