
The Effects of Ventilation, Humidity, and Temperature on Bacterial Growth and Bacterial Genera Distribution

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experiment
2.2. Equipment
2.3. Meat and Agar Preparation
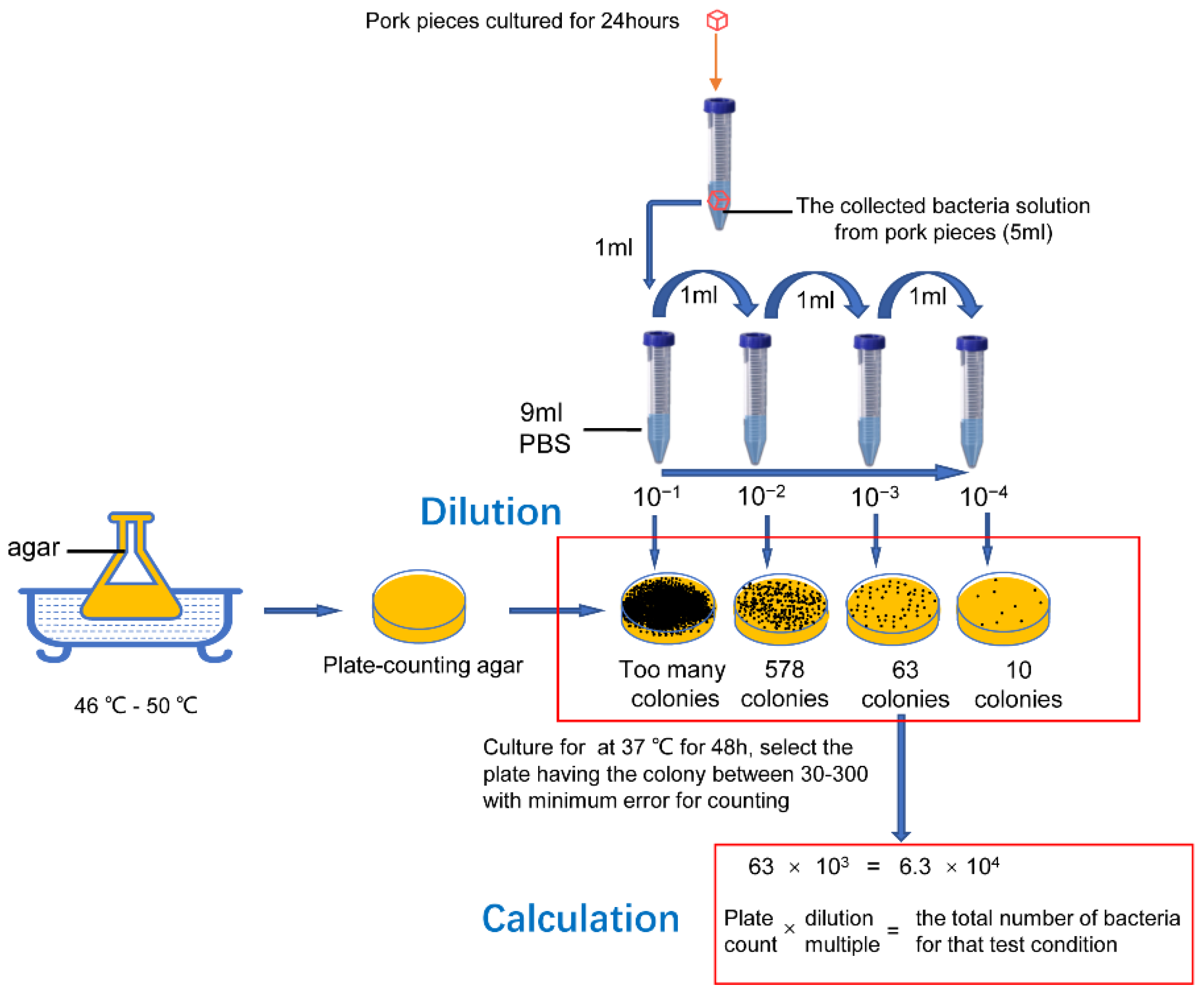
2.4. Bacteria Collection and Calculation
2.5. High-Throughput 16S rDNA Sequencing
2.6. Statistical Analysis
3. Results
3.1. Real-Time Temperature and Humidity under Different Working Conditions
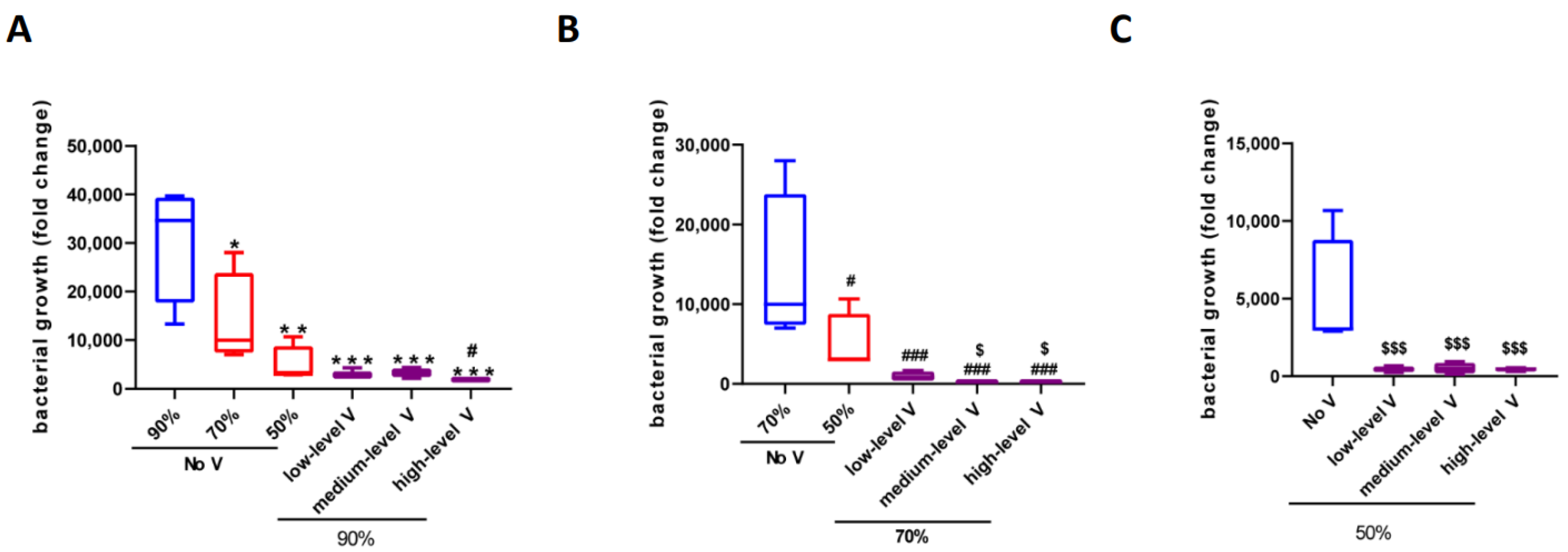
3.2. Effect of Humidity and Ventilation on Bacterial Growth at 26 °C
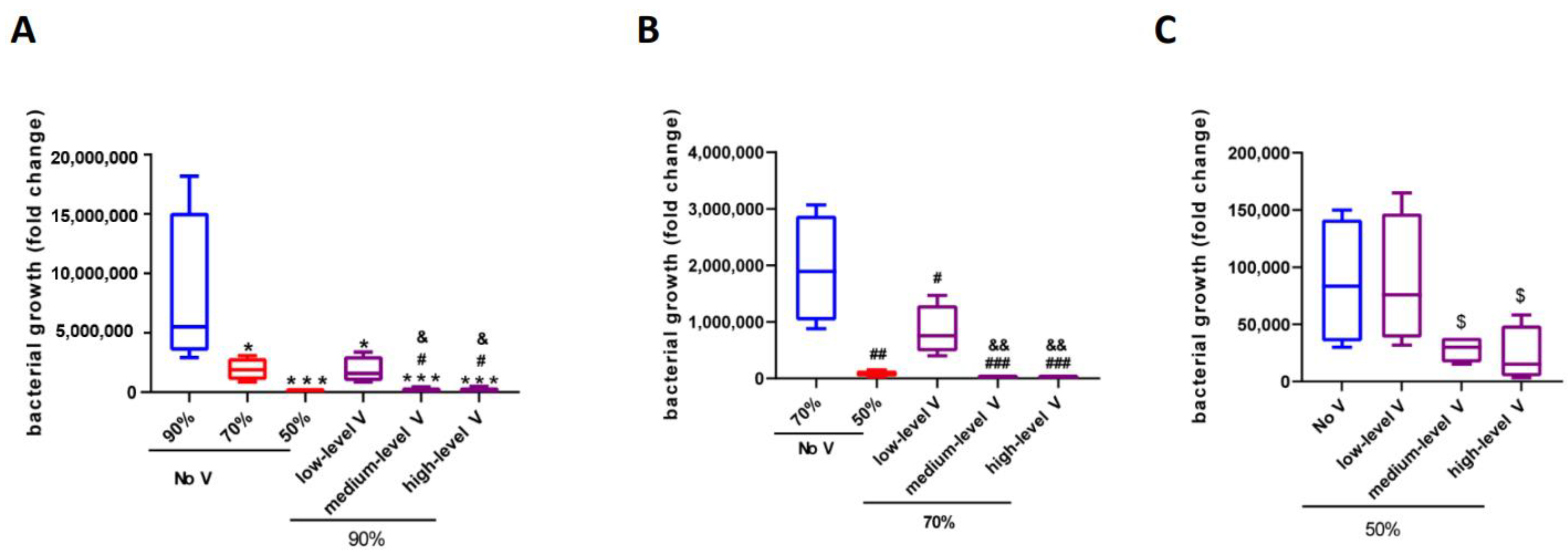
3.3. Effects of Humidity and Ventilation on Bacterial Growth at 34 °C
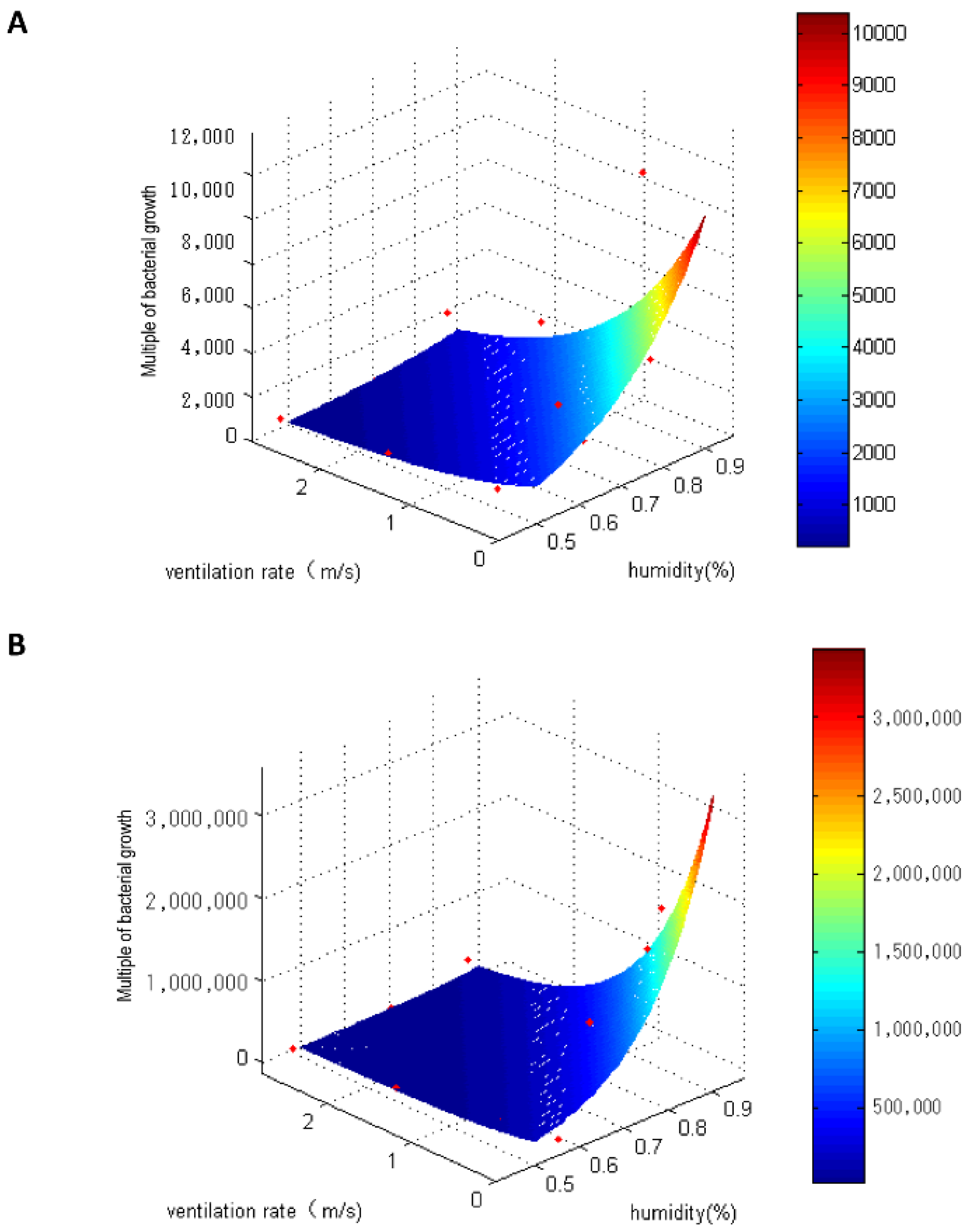
3.4. Bacterial Growth Model for Humidity and Ventilation
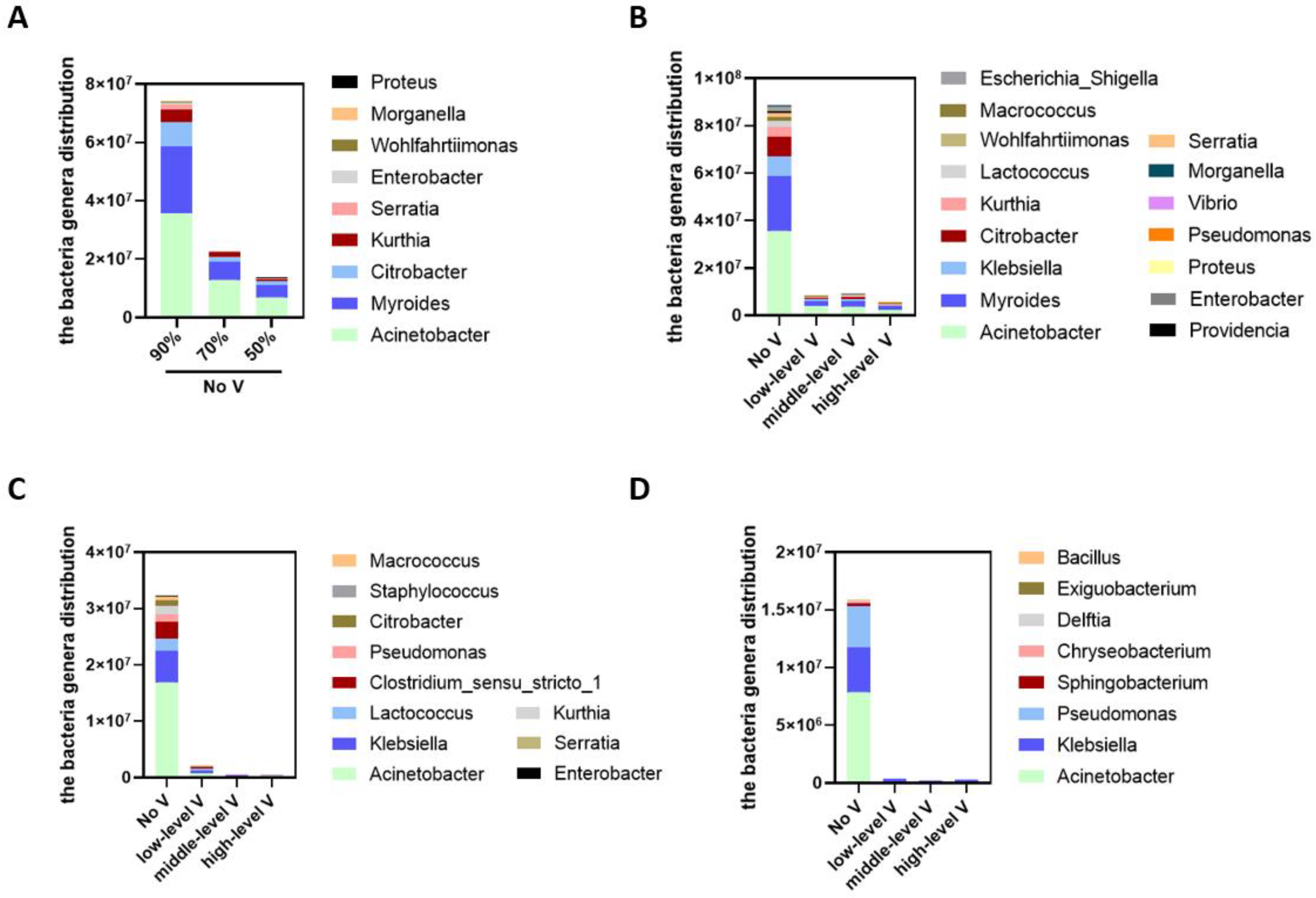
3.5. The Effect of Humidity and Ventilation on the Distribution of Bacterial Genera at 26 °C
3.6. The effect of Humidity and Ventilation on the Distribution of Bacteria Genera at 34 °C
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Igo, M.J.; Schaffner, D.W. Quantifying the Influence of Relative Humidity, Temperature, and Diluent on the Survival and Growth of Enterobacter aerogenes. J. Food Prot. 2019, 82, 2135–2147. [Google Scholar] [CrossRef]
- Kysela, D.T.; Brown, P.J.; Huang, K.C.; Brun, Y.V. Biological consequences and advantages of asymmetric bacterial growth. Annu. Rev. Microbiol. 2013, 67, 417–435. [Google Scholar] [CrossRef] [Green Version]
- Dannemiller, K.C.; Weschler, C.J.; Peccia, J. Fungal and bacterial growth in floor dust at elevated relative humidity levels. Indoor Air 2017, 27, 354–363. [Google Scholar] [CrossRef]
- Sinclair, R.G.; Gerba, C.P. Microbial contamination in kitchens and bathrooms of rural Cambodian village households. Lett. Appl. Microbiol. 2011, 52, 144–149. [Google Scholar] [CrossRef]
- Flores, G.E.; Bates, S.T.; Caporaso, J.G.; Lauber, C.L.; Leff, J.W.; Knight, R.; Fierer, N. Diversity, distribution and sources of bacteria in residential kitchens. Environ. Microbiol. 2013, 15, 588–596. [Google Scholar] [CrossRef] [PubMed]
- Abney, S.E.; Ijaz, M.K.; McKinney, J.; Gerba, C.P. Laundry Hygiene and Odor Control: State of the Science. Appl. Environ. Microbiol. 2021, 87, e0300220. [Google Scholar] [CrossRef] [PubMed]
- Shan, Y.; Wu, W.; Fan, W.; Haahtela, T.; Zhang, G. House dust microbiome and human health risks. Int. Microbiol. 2019, 22, 297–304. [Google Scholar] [CrossRef]
- Chai, Q.; Li, Y.; Li, X.; Wu, W.; Peng, H.; Jia, R.; Sun, Q. Assessment of variation in paddy microbial communities under different storage temperatures and relative humidity by Illumina sequencing analysis. Food Res. Int. 2019, 126, 108581. [Google Scholar] [CrossRef] [PubMed]
- Bockmuhl, D.P. Laundry hygiene-how to get more than clean. J. Appl. Microbiol. 2017, 122, 1124–1133. [Google Scholar] [CrossRef] [Green Version]
- Wolkoff, P. Indoor air humidity, air quality, and health—An overview. Int. J. Hyg. Environ. Health 2018, 221, 376–390. [Google Scholar] [CrossRef]
- Hillman, P.; Gebremedhin, K.; Warner, R. Ventilation System to Minimize Airborne Bacteria, Dust, Humidity, and Ammonia in Calf Nurseries. J. Dairy Sci. 1992, 75, 1305–1312. [Google Scholar] [CrossRef]
- Doulgeraki, A.I.; Ercolini, D.; Villani, F.; Nychas, G.J. Spoilage microbiota associated to the storage of raw meat in different conditions. Int. J. Food Microbiol. 2012, 157, 130–141. [Google Scholar] [CrossRef] [PubMed]
- Jain, R.; Anjaiah, V.; Babbar, S.B. Guar gum: A cheap substitute for agar in microbial culture media. Lett. Appl. Microbiol. 2005, 41, 345–349. [Google Scholar] [CrossRef] [PubMed]
- Brugger, S.D.; Baumberger, C.; Jost, M.; Jenni, W.; Brugger, U.; Muhlemann, K. Automated counting of bacterial colony forming units on agar plates. PLoS ONE 2012, 7, e33695. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; Noor, S.; Huan, R.; Liu, C.; Li, J.; Shi, Q.; Zhang, Y.J.; Wu, C.; He, H. Comparison of the diversity of cultured and total bacterial communities in marine sediment using culture-dependent and sequencing methods. PeerJ 2020, 8, e10060. [Google Scholar] [CrossRef]
- GB 4789.2-2022 National Food Safety Standard; Microbiological Examination of Food, Detection of Total Bacterial Count 2022. National Health Commission of the People’s Republic of China: Beijing, China, 2022. (In Chinese)
- Woo, P.C.; Lau, S.K.; Teng, J.L.; Tse, H.; Yuen, K.Y. Then and now: Use of 16S rDNA gene sequencing for bacterial identification and discovery of novel bacteria in clinical microbiology laboratories. Clin. Microbiol. Infect. 2008, 14, 908–934. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moat, A.G.; Foster, J.W.; Spector, M.P. Microbial Physiology; John Wiley & Sons: Hoboken, NJ, USA, 2002. [Google Scholar]
- Beuchat, L.R.; Komitopoulou, E.; Beckers, H.; Betts, R.P.; Bourdichon, F.; Fanning, S.; Joosten, H.M.; Ter Kuile, B.H. Low-water activity foods: Increased concern as vehicles of foodborne pathogens. J. Food Prot. 2013, 76, 150–172. [Google Scholar] [CrossRef]
- Yannick, D.N.; Tremblay, S.H.; Jacques, M. Bacterial biofilms: Their importance in animal health and public health. Can. J. Vet. Res. 2014, 78, 110–116. [Google Scholar]
- Tang, W.; Kuehn, T.H.; Simcik, M.F. Effects of Temperature, Humidity and Air Flow on Fungal Growth Rate on Loaded Ventilation Filters. J. Occup. Environ. Hyg. 2015, 12, 525–537. [Google Scholar] [CrossRef]
- Esbelin, J.; Santos, T.; Hebraud, M. Desiccation: An environmental and food industry stress that bacteria commonly face. Food Microbiol. 2018, 69, 82–88. [Google Scholar] [CrossRef]
- Rabia, S. 2021 ASHRAE Handbook Fundamentals (I P); Section 9.3; ASHARE: Peachtree Corners, GA, USA, 2021. [Google Scholar]
- Visca, P.; Seifert, H.; Towner, K.J. Acinetobacter infection--an emerging threat to human health. IUBMB Life 2011, 63, 1048–1054. [Google Scholar] [CrossRef] [PubMed]
- Wyres, K.L.; Lam, M.M.C.; Holt, K.E. Population genomics of Klebsiella pneumoniae. Nat. Rev. Microbiol. 2020, 18, 344–359. [Google Scholar] [CrossRef] [PubMed]
- Lambiase, A.; Del Pezzo, M.; Raia, V.; Sepe, A.; Ferri, P.; Rossano, F. Chryseobacterium respiratory tract infections in patients with cystic fibrosis. J. Infect. 2007, 55, 518–523. [Google Scholar] [CrossRef] [PubMed]
- Acosta-Ochoa, M.I.; Rodrigo-Parra, A.; Rodriguez-Martin, F.; Molina-Miguel, A. Urinary infection due to Chryseobacterium indologenes. Nefrologia 2013, 33, 620. [Google Scholar] [CrossRef]
- Deveci, A.; Coban, A.Y. Optimum management of Citrobacter koseri infection. Expert. Rev. Anti-Infect. Ther. 2014, 12, 1137–1142. [Google Scholar] [CrossRef] [PubMed]
- Hu, S.H.; Yuan, S.X.; Qu, H.; Jiang, T.; Zhou, Y.J.; Wang, M.X.; Ming, D.S. Antibiotic resistance mechanisms of Myroides sp. J. Zhejiang Univ. Sci. B 2016, 17, 188–199. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Wang, L.; Zhou, J.; Wu, H.; Li, D.; Cui, Y.; Lu, B. Exiguobacterium sp. A1b/GX59 isolated from a patient with community-acquired pneumonia and bacteremia: Genomic characterization and literature review. BMC Infect. Dis. 2017, 17, 508. [Google Scholar] [CrossRef]
- Lund, B.M. Foodborne disease due to Bacillus and Clostridium species. Lancet 1990, 336, 982–986. [Google Scholar] [CrossRef]
- Jovanovic, J.; Ornelis, V.F.M.; Madder, A.; Rajkovic, A. Bacillus cereus food intoxication and toxicoinfection. Compr. Rev. Food Sci. Food Saf. 2021, 20, 3719–3761. [Google Scholar] [CrossRef]
- Frieri, M.; Kumar, K.; Boutin, A. Antibiotic resistance. J. Infect. Public Health 2017, 10, 369–378. [Google Scholar] [CrossRef] [Green Version]
- Zagaliotis, P.; Michalik-Provasek, J.; Gill, J.J.; Walsh, T.J. Therapeutic Bacteriophages for Gram-Negative Bacterial Infections in Animals and Humans. Pathog. Immun. 2022, 7, 1–45. [Google Scholar] [CrossRef] [PubMed]
- Pendleton, J.N.; Gorman, S.P.; Gilmore, B.F. Clinical relevance of the ESKAPE pathogens. Expert. Rev. Anti-Infect. Ther. 2013, 11, 297–308. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 26 °C | No V | Low-Level V | Middle-Level V | High-Level V |
|---|---|---|---|---|
| 90% | 3.06 ± 0.61 × 104 | 3.20 ± 0.56 × 103 | 3.30 ± 0.47 × 103 | 1.87 ± 0.08 × 103 |
| 70% | 1.38 ± 0.48 × 104 | 9.31 ± 3.18 × 102 | 1.93 ± 0.33 × 102 | 2.33 ± 0.47 × 102 |
| 50% | 4.91 ± 1.92 × 103 | 4.45 ± 0.95 × 102 | 5.21 ± 1.77 × 102 | 4.51 ± 0.49 × 102 |
| 34 °C | No V | Low-Level V | Middle-Level V | High-Level V |
|---|---|---|---|---|
| 90% | 8.03 ± 3.44 × 106 | 1.85 ± 0.56 × 106 | 2.01 ± 0.78 × 105 | 2.11 ± 0.97 × 105 |
| 70% | 1.93 ± 0.48 × 106 | 8.46 ± 2.24 × 105 | 1.14 ± 0.13 × 104 | 1.16 ± 0.20 × 104 |
| 50% | 8.67 ± 2.81 × 104 | 8.71 ± 2.89 × 104 | 2.83 ± 0.59 × 104 | 2.29 ± 1.24 × 104 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qiu, Y.; Zhou, Y.; Chang, Y.; Liang, X.; Zhang, H.; Lin, X.; Qing, K.; Zhou, X.; Luo, Z. The Effects of Ventilation, Humidity, and Temperature on Bacterial Growth and Bacterial Genera Distribution. Int. J. Environ. Res. Public Health 2022, 19, 15345. https://doi.org/10.3390/ijerph192215345
Qiu Y, Zhou Y, Chang Y, Liang X, Zhang H, Lin X, Qing K, Zhou X, Luo Z. The Effects of Ventilation, Humidity, and Temperature on Bacterial Growth and Bacterial Genera Distribution. International Journal of Environmental Research and Public Health. 2022; 19(22):15345. https://doi.org/10.3390/ijerph192215345
Chicago/Turabian StyleQiu, Yujia, Yan Zhou, Yanfen Chang, Xinyue Liang, Hui Zhang, Xiaorui Lin, Ke Qing, Xiaojie Zhou, and Ziqiang Luo. 2022. "The Effects of Ventilation, Humidity, and Temperature on Bacterial Growth and Bacterial Genera Distribution" International Journal of Environmental Research and Public Health 19, no. 22: 15345. https://doi.org/10.3390/ijerph192215345
APA StyleQiu, Y., Zhou, Y., Chang, Y., Liang, X., Zhang, H., Lin, X., Qing, K., Zhou, X., & Luo, Z. (2022). The Effects of Ventilation, Humidity, and Temperature on Bacterial Growth and Bacterial Genera Distribution. International Journal of Environmental Research and Public Health, 19(22), 15345. https://doi.org/10.3390/ijerph192215345