
Canada Goldenrod Invasion Regulates the Effects of Soil Moisture on Soil Respiration

 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Design
2.2. Soil, Autotrophic, and Heterotrophic Respiration Measurements
2.3. Soil and Vegetation Sample Collection and Preparation
2.4. Soil Characteristics
2.5. Statistical Analyses
3. Results
3.1. Effects of Treatments on Soil Total, Heterotrophic, and Autotrophic Respiration
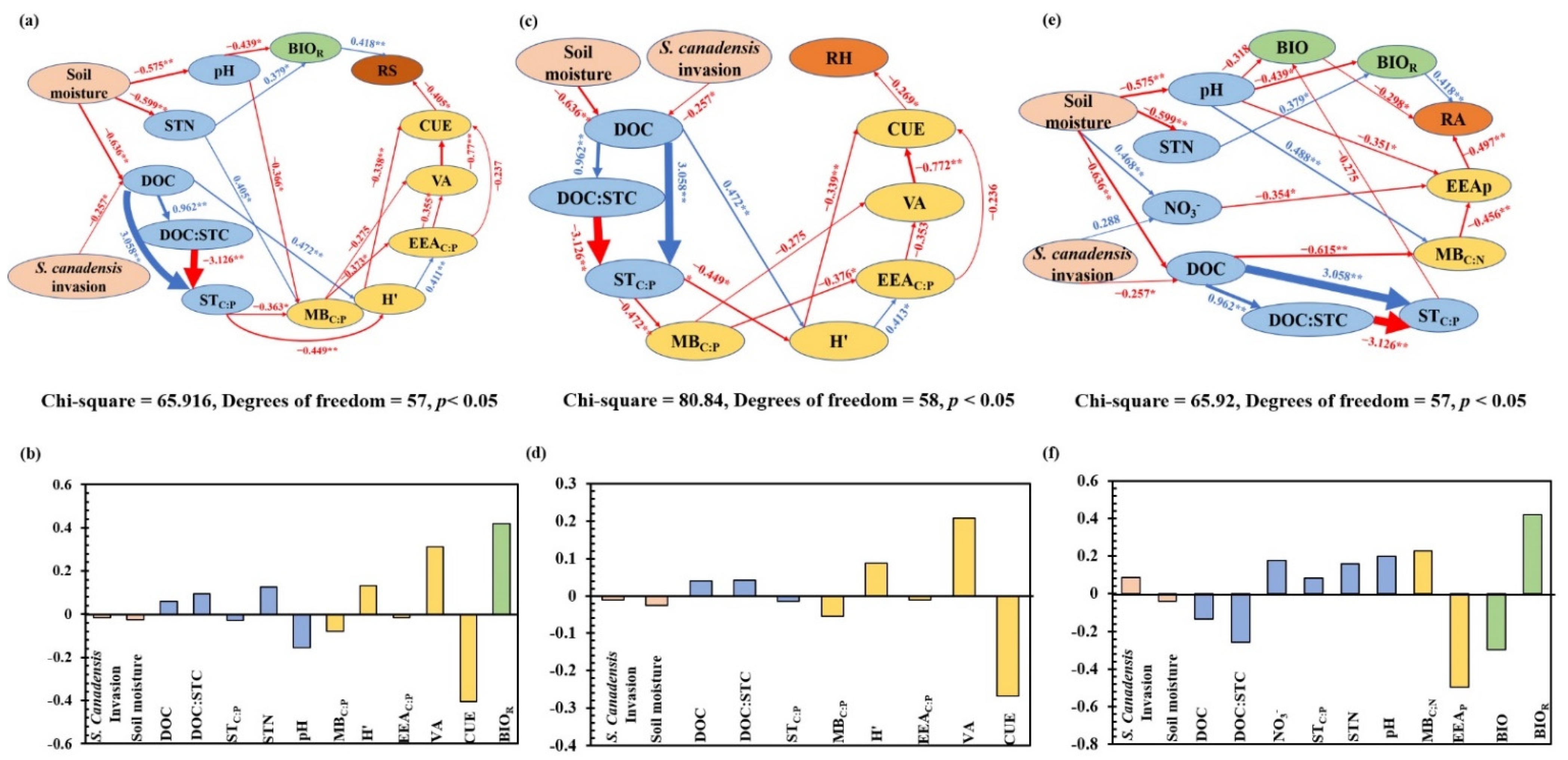
3.2. Correlations of Soil Total, Heterotrophic, and Autotrophic Respiration with Soil Characteristics and Plant Biomass
4. Discussion
4.1. Effects of S. Canadensis Invasion on Soil Total, Heterotrophic, and Autotrophic Respiration
4.2. Effects of Water Level on Soil Total, Heterotrophic, and Autotrophic Respiration
4.3. S. canadensis Invasion Affects Soil Respiration Responses to Soil Moisture
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Bielecka, A.; Borkowska, L.; Krolak, E. Environmental changes caused by the clonal invasive plant Solidago canadensis. Ann. Bot. Fenn. 2020, 57, 33–48. [Google Scholar] [CrossRef]
- Liao, M.; Xie, X.M.; Peng, Y.; Ma, A. Changes of soil microbiological characteristics after Solidago canadensis L. invasion. Ang. Sci. China. 2011, 10, 1064–1071. [Google Scholar] [CrossRef]
- Zhang, L.; Ma, X.C.; Wang, H.; Liu, S.W.; Siemann, E.; Zou, J.W. Soil respiration and litter decomposition increased following Perennial Forb invasion into an annual grassland. Pedosphere 2016, 26, 567–576. [Google Scholar] [CrossRef]
- Trigos-Peral, G.; Casacci, L.P.; Ślipiński, P.; Grześ, I.M.; Moroń, D.; Babik, H.; Witek, M. Ant communities and Solidago plant invasion: Environmental properties and food sources. Entomol. Sci. 2018, 21, 270–278. [Google Scholar] [CrossRef]
- Liao, M.; Xie, X.M.; Peng, Y.; Chai, J.J.; Chen, N. Characteristics of soil microbial community function and structure diversity with coverage of Solidago canadensis L. J. Cent. South Univ. 2013, 20, 749–756. [Google Scholar] [CrossRef]
- Zhang, C.B.; Wang, J.; Qian, B.Y.; Li, W.H. Effects of the invader Solidago canadensis on soil properties. Appl. Soil Ecol. 2009, 43, 163–169. [Google Scholar] [CrossRef]
- Chen, Z.Y.; Fu, C.C.; Wang, H.Y.; Li, B.; Wu, J.H.; Chen, J.K. Effects of Spartina alterniflora invasions on the benthic macro-invertebrates community at Dongtan of Chongming salt Marsh, the Yangtze River estuary. Wetland Sci. 2005, 3, 1–7. [Google Scholar]
- Zedler, J.B.; Kercher, S. Causes and consequences of invasive plants in wetlands: Opportunities, opportunists, and outcomes. Crit. Rev. Plant Sci. 2004, 23, 431–452. [Google Scholar] [CrossRef]
- Yusheng, Y.; Bin, D.; Jinsheng, X.; Guangshui, C.; Ren, G.; Ling, L.; Xiaogui, W.; Jianfen, G. Soil respiration of forest ecosystems and its respondence to global change. Acta. Ecol. Sin. 2004, 24, 583–591. [Google Scholar]
- Chang, S.X.; Shi, Z.; Thomas, B.R. Soil respiration and its temperature sensitivity in agricultural and afforested poplar plantation systems in northern Alberta. Biol. Fert. Soils 2016, 52, 629–641. [Google Scholar] [CrossRef]
- Yu, S.; Chen, Y.; Zhao, J.; Fu, S.; Li, Z.; Xia, H.; Zhou, L. Temperature sensitivity of total soil respiration and its heterotrophic and autotrophic components in six vegetation types of subtropical China. Sci. Total Environ. 2017, 607, 160–167. [Google Scholar] [CrossRef]
- Li, M.; Liu, T.; Duan, L.; Ma, L.; Wang, Y.X.; Zhou, Y.J.; Li, Y.K.; Zhao, X.Y.; Wang, X.Q.; Wang, G.Q.; et al. Hydrologic gradient changes of soil respiration in typical steppes of Eurasia. Sci. Total Environ. 2021, 794, 148684. [Google Scholar] [CrossRef] [PubMed]
- Marchante, E.; Kjøller, A.; Struwe, S.; Freitas, H. Short-and long-term impacts of Acacia longifolia invasion on the belowground processes of a Mediterranean coastal dune ecosystem. Appl. Soil Ecol. 2008, 40, 210–217. [Google Scholar] [CrossRef] [Green Version]
- Potts, D.L.; Harpole, W.S.; Goulden, M.L.; Suding, K.N. The impact of invasion and subsequent removal of an exotic thistle, Cynara cardunculus, on CO2 and H2O vapor exchange in a coastal California grassland. Biol. Invasions 2008, 10, 1073–1084. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; Wang, W.Q.; Sardans, J.; Ouyang, L.M.; Tong, C.; Asensio, D.; Gargallo, A.; Wiesmeier, M.; Penuelas, J. Higher fluxes of C, N and P in plant/soil cycles associated with plant invasion in a subtropical estuarine wetland in China. Sci. Total Environ. 2020, 730, 139124. [Google Scholar] [CrossRef] [PubMed]
- Gao, G.F.; Li, P.F.; Shen, Z.J.; Qin, Y.Y.; Zhang, X.M.; Ghoto, K.; Zhu, X.Y.; Zheng, H.L. Exotic Spartina alterniflora invasion increases CH4 while reduces CO2 emissions from mangrove wetland soils in southeastern China. Sci. Rep. 2018, 8, 9243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.; Zhang, Y.; Zou, J.; Siemann, E. Decomposition of Phragmites australis litter retarded by invasive Solidago canadensis in mixtures: An antagonistic non-additive effect. Sci. Rep. 2014, 4, 5488. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Wang, H.; Zou, J.; Rogers, W.E.; Siemann, E. Non-native plant litter enhances soil carbon dioxide emissions in an invaded annual grassland. PLoS ONE 2014, 9, e92301. [Google Scholar] [CrossRef] [Green Version]
- Cui, H.; Bai, J.; Du, S.; Wang, J.; Keculah, G.N.; Wang, W.; Zhang, G.L.; Jia, J. Interactive effects of groundwater level and salinity on soil respiration in coastal wetlands of a Chinese delta. Environ Pollut. 2021, 286, 117400. [Google Scholar] [CrossRef] [PubMed]
- Duval, B.D.; Curtsinger, H.D.; Hands, A.; Martin, J.; McLaren, J.R.; Cadol, D.D. Greenhouse gas emissions and extracellular enzyme activity variability during decomposition of native versus invasive riparian tree litter. Plant Ecol. 2020, 221, 177–189. [Google Scholar] [CrossRef]
- Chen, Y.P.; Chen, G.C.; Ye, Y. Coastal vegetation invasion increases greenhouse gas emission from wetland soils but also increases soil carbon accumulation. Sci. Total Environ. 2015, 526, 19–28. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.; Jeelani, N.; Leng, X.; Cheng, X.; An, S. Spartina alterniflora invasion alters soil microbial community composition and microbial respiration following invasion chronosequence in a coastal wetland of China. Sci. Rep. 2016, 6, 26880. [Google Scholar] [CrossRef] [Green Version]
- Jones, G.L.; Scullion, J.; Allison, G.; Stott, H.; Johnson, D.; Owen, R.; Williams, G.; Gwynn-Jones, D. Reduced soil respiration beneath invasive Rhododendron ponticum persists after cutting and is related to substrate quality rather than microbial community. Soil Biol. Biochem. 2019, 138, 107577. [Google Scholar] [CrossRef]
- Zhang, L.; Wang, S.; Liu, S.; Liu, X.; Zou, J.; Siemann, E. Perennial forb invasions alter greenhouse gas balance between ecosystem and atmosphere in an annual grassland in China. Sci. Total Environ. 2018, 642, 781–788. [Google Scholar] [CrossRef]
- Ledger, K.J.; Pal, R.W.; Murphy, P.; Nagy, D.U.; Filep, R.; Callaway, R.M. Impact of an invader on species diversity is stronger in the non-native range than in the native range. Plant Ecol. 2015, 216, 1285–1295. [Google Scholar] [CrossRef]
- Maier, C.A.; Kress, L.W. Soil CO2 evolution and root respiration in 11 year-old loblolly pine (Pinus taeda) plantations as affected by moisture and nutrient availability. Can. J. For. Res. 2000, 30, 347–359. [Google Scholar] [CrossRef]
- O’neill, K.P.; Kasischke, E.S.; Richter, D.D. Environmental controls on soil CO2 flux following fire in black spruce, white spruce, and aspen stands of interior Alaska. Can. J. For. Res. 2002, 32, 1525–1541. [Google Scholar] [CrossRef]
- Rustad, L.E.; Huntington, T.G.; Boone, R.D. Controls on soil respiration: Implications for climate change. Biogeochemistry 2000, 48, 1–6. [Google Scholar] [CrossRef]
- Szymura, M.; Szymura, T.H. Soil preferences and morphological diversity of goldenrods (Solidago L.) from south-western Poland. Acta. Soc. Bot. Pol. 2013, 82, 107–115. [Google Scholar] [CrossRef] [Green Version]
- Park, J.S.; Choi, D.; Kim, Y. Potential distribution of goldenrod (Solidago altissima L.) during climate change in South Korea. Sustainability 2020, 12, 6710. [Google Scholar] [CrossRef]
- Jian-Zhong, L.U.; En-Sheng WE, N.G.; Xiao-Wen, W.U.; Weber, E.; Bin ZH, A.O.; Bo, L.I. Potential distribution of Solidago Canadensis in China. Acta Phytotaxonomica Sinica 2007, 45, 670–674. [Google Scholar]
- Yang, S.; Geng, Q.; Xu, C.; Peng, F.; Zhang, M.; Xu, X. Effects of Solidago canadensis L. invasion on soil respiration in poplar plantations (Populus deltoides). J. Nanjing For. Univ. Nat. Sci. Ed. 2020, 44, 117. [Google Scholar]
- Hu, Z.; Li, J.; Shi, K.; Ren, G.; Dai, Z.; Sun, J.; Zheng, X.; Zhou, Y.; Zhang, J.; Li, G.; et al. Effects of Canada goldenrod invasion on soil extracellular enzyme activities and ecoenzymatic stoichiometry. Sustainability 2021, 13, 3768. [Google Scholar] [CrossRef]
- Hu, Z.; Zhang, J.; Du, Y.; Shi, K.; Ren, G.; Iqbal, B.; Dai, Z.; Li, J.; Li, G.; Du, D. Substrate availability regulates the suppressive effects of Canada goldenrod invasion on soil respiration. J. Plant Ecol. 2022, 15, 509–523. [Google Scholar] [CrossRef]
- Wang, S.; Wei, M.; Wu, B.; Cheng, H.; Wang, C. Combined nitrogen deposition and Cd stress antagonistically affect the allelopathy of invasive alien species Canada goldenrod on the cultivated crop lettuce. Sci. Hortic. 2020, 261, 108955. [Google Scholar] [CrossRef]
- Zhang, H.Y.; Goncalves, P.; Copeland, E.; Qi, S.S.; Dai, Z.C.; Li, G.L.; Wang, C.; Du, D.; Thomas, T. Invasion by the weed Conyza canadensis alters soil nutrient supply and shifts microbiota structure. Soil Biol. Biochem. 2020, 143, 107739. [Google Scholar] [CrossRef]
- Noh, N.J.; Lee, S.J.; Jo, W.; Han, S.; Yoon, T.K.; Chung, H.; Muraoka, H.; Son, Y. Effects of experimental warming on soil respiration and biomass in Quercus variabilis Blume and Pinus densiflora Sieb. et Zucc. seedlings. An. For. Sci. 2016, 73, 533–545. [Google Scholar] [CrossRef] [Green Version]
- Li, G.; Kim, S.; Han, S.H.; Chang, H.; Son, Y. Effect of soil moisture on the response of soil respiration to open-field experimental warming and precipitation manipulation. Forests 2017, 8, 56. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.; Liu, Y.; Gu, Y.; Guo, L.; Huang, Y.; Zhang, J.; Xu, Z.; Tan, B.; Zhang, L.; Chen, L.; et al. Ecoenzymatic stoichiometry and microbial nutrient limitations in rhizosphere soil along the Hailuogou Glacier forefield chronosequence. Sci. Total Environ. 2020, 704, 135413. [Google Scholar] [CrossRef] [PubMed]
- Miranda, K.M.; Espey, M.G.; Wink, D.A. A rapid, simple spectrophotometric method for simultaneous detection of nitrate and nitrite. Nitric Oxide 2001, 5, 62–71. [Google Scholar] [CrossRef]
- Doane, T.A.; Horwáth, W.R. Spectrophotometric determination of nitrate with a single reagent. Anal. Lett. 2003, 36, 2713–2722. [Google Scholar] [CrossRef]
- Olsen, S.R.; Sommers, L.E. Phosphorus in Methods of Soil Analysis, Part 2-Chemical and Microbiological Properties; Page, A.L., Ed.; ASA, Inc.: Hongkong, China, 1982; pp. 403–430. [Google Scholar]
- Murphy, J.; Riley, J.P. A modified single solution method for the determination of phosphate in natural waters. Anal. Chim. Acta 1962, 27, 31–36. [Google Scholar] [CrossRef]
- Brookes, P.C.; Landman, A.; Pruden, G.; Jenkinson, D.S. Chloroform fumigation and the release of soil nitrogen: A rapid direct extraction method to measure microbial biomass nitrogen in soil. Soil Biol Biochem. 1985, 17, 837–842. [Google Scholar] [CrossRef]
- Vance, E.D.; Brookes, P.C.; Jenkinson, D.S. An extraction method for measuring soil microbial biomass C. Soil Biol Biochem. 1987, 19, 703–707. [Google Scholar] [CrossRef]
- Guanlin, L.I.; Seongjun KI, M.; Minji PA, R.K.; Yowhan, S.O.N. Short-term effects of experimental warming and precipitation manipulation on soil microbial biomass C and N, community substrate utilization patterns and community composition. Pedosphere 2017, 27, 714–724. [Google Scholar]
- DeForest, J.L. The influence of time, storage temperature, and substrate age on potential soil enzyme activity in acidic forest soils using MUB-linked substrates and L-DOPA. Soil Biol Biochem. 2009, 41, 1180–1186. [Google Scholar] [CrossRef]
- Cui, Y.; Wang, X.; Zhang, X.; Ju, W.; Duan, C.; Guo, X.; Wang, Y.; Fang, L. Soil moisture mediates microbial carbon and phosphorus metabolism during vegetation succession in a semiarid region. Soil Biol Biochem. 2020, 147, 107814. [Google Scholar] [CrossRef]
- Wei, H.; Yan, W.; Quan, G.; Zhang, J.; Liang, K. Soil microbial carbon utilization, enzyme activities and nutrient availability responses to Bidens pilosa and a non-invasive congener under different irradiances. Sci. Rep. 2017, 7, 11309. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Cheng, H.; Wei, M.; Wu, B.; Wang, C. Litter decomposition process dramatically declines the allelopathy of Solidago canadensis L. on the seed germination and seedling growth of Lactuca sativa L. Int. J. Phytoremediat. 2020, 22, 1295–1303. [Google Scholar] [CrossRef]
- Zhang, D.; Ye, Y.; Li, J.; Dong, L. Allelopathic pathways, isolation and identification of an allelopathic substance from Solidago canadensis L. Allelopath. J. 2014, 33, 201. [Google Scholar]
- Zhang, Z.; Chen, L.; Wang, J.; Yao, J.; Li, J. Biochar preparation from Solidago canadensis and its alleviation of the inhibition of tomato seed germination by allelochemicals. RSC Adv. 2018, 8, 22370–22375. [Google Scholar] [CrossRef] [Green Version]
- Prevéy, J.S.; Germino, M.J.; Huntly, N.J.; Inouye, R.S. Exotic plants increase and native plants decrease with loss of foundation species in sagebrush steppe. Plant Ecol. 2010, 207, 39–51. [Google Scholar] [CrossRef]
- Lin, Z.; Zhang, R.; Tang, J.; Zhang, J. Effects of high soil water content and temperature on soil respiration. Soil Sci. 2011, 176, 150–155. [Google Scholar] [CrossRef]
- Chen, X.; Post, W.M.; Norby, R.J.; Classen, A.T. Modeling soil respiration and variations in source components using a multi-factor global climate change experiment. Clim. Chang. 2011, 107, 459–480. [Google Scholar] [CrossRef]
- Balogh, J. Dependence of soil respiration on soil moisture, clay content, soil organic matter, and CO2 uptake in dry grasslands. Soil Biol Biochem. 2014, 78, 341. [Google Scholar] [CrossRef]
- Jia, B.; Zhou, G.; Wang, Y.; Wang, F.; Wang, X. Effects of temperature and soil water-content on soil respiration of grazed and ungrazed Leymus chinensis steppes, Inner Mongolia. J. Arid Environ. 2006, 67, 60–76. [Google Scholar] [CrossRef]
- Ge, Y.; Wang, Q.; Wang, L.; Liu, W.; Liu, X.; Huang, Y.; Christie, P. Response of soil enzymes and microbial communities to root extracts of the alien Alternanthera philoxeroides. Arch. Agron. Soil Sci. 2018, 64, 708–717. [Google Scholar] [CrossRef]
- Tang, X.; Pei, X.; Lei, N.; Luo, X.; Liu, L.; Shi, L.; Chen, G.; Liang, J. Global patterns of soil autotrophic respiration and its relation to climate, soil and vegetation characteristics. Geoderma 2020, 369, 114339. [Google Scholar] [CrossRef]
- Liu, Y.; Liu, S.; Wan, S.; Wang, J.; Luan, J.; Wang, H. Differential responses of soil respiration to soil warming and experimental throughfall reduction in a transitional oak forest in central China. Agric. For. Meteorol. 2016, 226–227, 186–198. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| S. canadensis Invasion | Water Level | Interactive Effect | |||||||
|---|---|---|---|---|---|---|---|---|---|
| df | F | p | df | F | p | df | F | p | |
| RS | 5 | 38.16 | ** 1 | 2 | 3.52 | * | 10 | 0.52 | 0.87 |
| RA | 5 | 40.85 | ** | 2 | 3.97 | * | 10 | 0.66 | 0.76 |
| RH | 5 | 23.33 | ** | 2 | 1.48 | 0.23 | 10 | 0.44 | 0.92 |
| S. canadensis Invasion | Water Level | S. canadensis Invasion * Soil Moisture | |||||||
|---|---|---|---|---|---|---|---|---|---|
| df | F | p | df | F | p | df | F | p | |
| RS | 3 | 0.26 | 0.62 | 3 | 11.92 | ** 1 | 9 | 7.8 | ** |
| RA | 3 | 1.88 | 0.18 | 3 | 30.22 | ** | 9 | 2.55 | 0.12 |
| RH | 3 | 2.86 | 0.09 | 3 | 19.16 | ** | 9 | 3.11 | 0.08 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, S.; Li, K.; Li, G.; Hu, Z.; Zhang, J.; Iqbal, B.; Du, D. Canada Goldenrod Invasion Regulates the Effects of Soil Moisture on Soil Respiration. Int. J. Environ. Res. Public Health 2022, 19, 15446. https://doi.org/10.3390/ijerph192315446
Xu S, Li K, Li G, Hu Z, Zhang J, Iqbal B, Du D. Canada Goldenrod Invasion Regulates the Effects of Soil Moisture on Soil Respiration. International Journal of Environmental Research and Public Health. 2022; 19(23):15446. https://doi.org/10.3390/ijerph192315446
Chicago/Turabian StyleXu, Sixuan, Kexin Li, Guanlin Li, Zhiyuan Hu, Jiaqi Zhang, Babar Iqbal, and Daolin Du. 2022. "Canada Goldenrod Invasion Regulates the Effects of Soil Moisture on Soil Respiration" International Journal of Environmental Research and Public Health 19, no. 23: 15446. https://doi.org/10.3390/ijerph192315446