
Evaluation of the Pathogenic-Mixed Biofilm Formation of Pseudomonas aeruginosa/Staphylococcus aureus and Treatment with Limonene on Three Different Materials by a Dynamic Model

 , ,
, ,  ,
,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains, Growth Conditions, Chemicals, and Test Materials
2.2. Susceptibility Test of Planktonic Cells
2.3. Biofilm Formation and Inhibition/Eradication with Limonene in CDC Biofilm Reactor® Model
2.4. qRT-PCR Analysis
2.5. Statistical Analysis
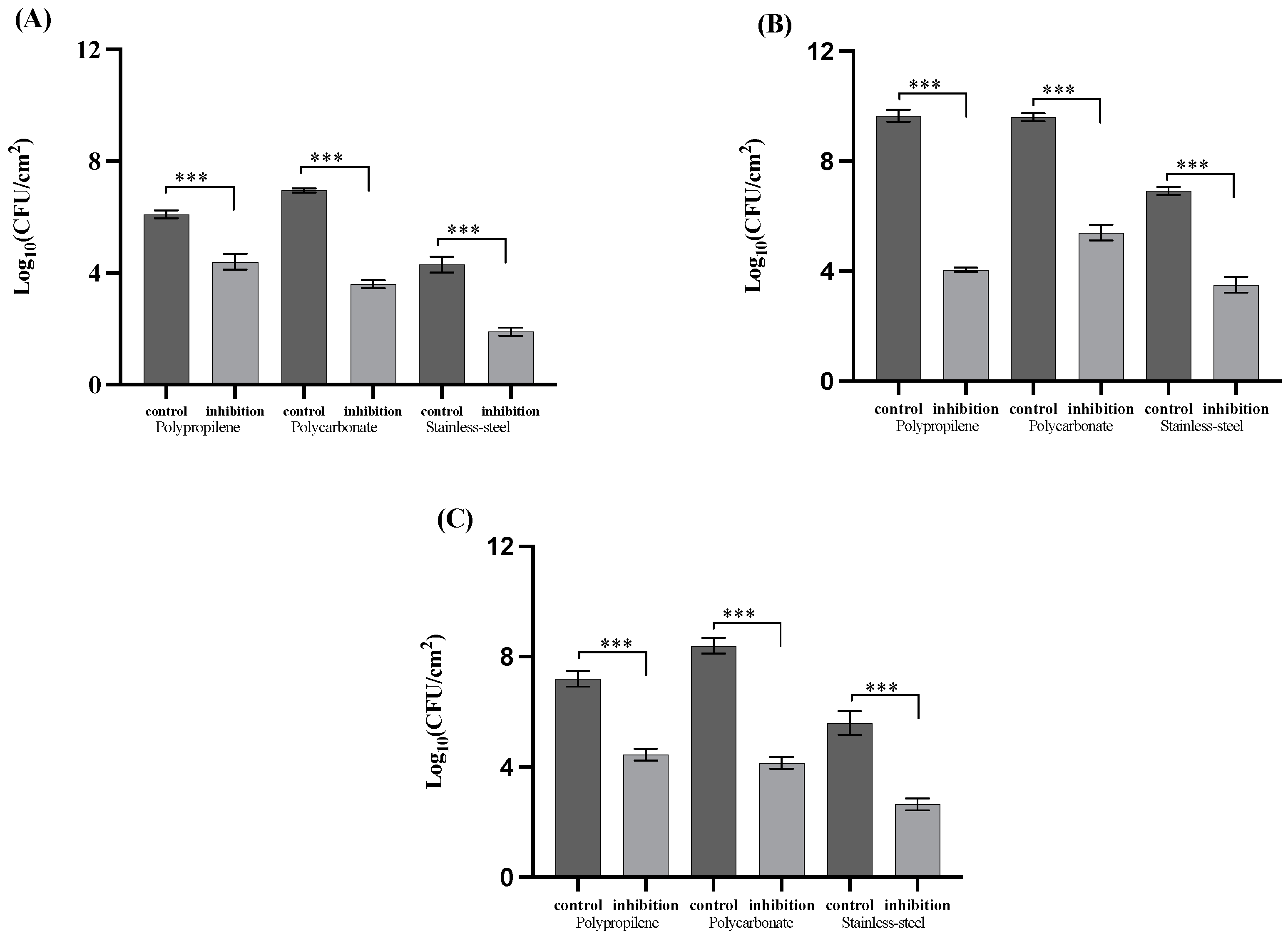
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Del Pozo, J.L. Biofilm-Related Disease. Expert Rev. Anti Infect. Ther. 2018, 16, 51–65. [Google Scholar] [CrossRef]
- Jamal, M.; Ahmad, W.; Andleeb, S.; Jalil, F.; Imran, M.; Nawaz, M.A.; Hussain, T.; Ali, M.; Rafiq, M.; Kamil, M.A. Bacterial biofilm and associated infections. J. Chin. Med. Assoc. JCMA 2018, 81, 7–11. [Google Scholar] [CrossRef] [PubMed]
- Azeredo, J.; Azevedo, N.F.; Briandet, R.; Cerca, N.; Coenye, T.; Costa, A.R.; Desvaux, M.; Di Bonaventura, G.; Hébraud, M.; Jaglic, Z.; et al. Critical Review on Biofilm Methods. Crit. Rev. Microbiol. 2017, 43, 313–351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wicaksono, W.A.; Erschen, S.; Krause, R.; Müller, H.; Cernava, T.; Berg, G. Enhanced Survival of Multi-Species Biofilms under Stress Is Promoted by Low-Abundant but Antimicrobial-Resistant Keystone Species. J. Hazard. Mater. 2022, 422, 126836. [Google Scholar] [CrossRef] [PubMed]
- Ibberson, C.B.; Whiteley, M. The Social Life of Microbes in Chronic Infection. Curr. Opin. Microbiol. 2020, 53, 44–50. [Google Scholar] [CrossRef]
- Hotterbeekx, A.; Kumar-Singh, S.; Goossens, H.; Malhotra-Kumar, S. In Vivo and In Vitro Interactions between Pseudomonas aeruginosa and Staphylococcus Spp. Front. Cell. Infect. Microbiol. 2017, 7, 106. [Google Scholar] [CrossRef] [Green Version]
- Woods, P.W.; Haynes, Z.M.; Mina, E.G.; Marques, C.N.H. Maintenance of S. aureus in Co-Culture with P. aeruginosa While Growing as Biofilms. Front. Microbiol. 2019, 9, 3291. [Google Scholar] [CrossRef] [Green Version]
- Briaud, P.; Camus, L.; Bastien, S.; Doléans-Jordheim, A.; Vandenesch, F.; Moreau, K. Coexistence with Pseudomonas aeruginosa Alters Staphylococcus aureus Transcriptome, Antibiotic Resistance and Internalization into Epithelial Cells. Sci. Rep. 2019, 9, 16564. [Google Scholar] [CrossRef] [Green Version]
- Di Onofrio, V.; Gesuele, R.; Maione, A.; Liguori, G.; Liguori, R.; Guida, M.; Nigro, R.; Galdiero, E. Prevention of Pseudomonas aeruginosa Biofilm Formation on Soft Contact Lenses by Allium Sativum Fermented Extract (BGE) and Cannabinol Oil Extract (CBD). Antibiotics 2019, 8, 258. [Google Scholar] [CrossRef] [Green Version]
- Reichling, J. Anti-Biofilm and Virulence Factor-Reducing Activities of Essential Oils and Oil Components as a Possible Option for Bacterial Infection Control. Planta Med. 2020, 86, 520–537. [Google Scholar] [CrossRef]
- Chouhan, S.; Sharma, K.; Guleria, S. Antimicrobial Activity of Some Essential Oils—Present Status and Future Perspectives. Medicines 2017, 4, 58. [Google Scholar] [CrossRef] [Green Version]
- Gupta, P.; Pruthi, V.; Poluri, K.M. Mechanistic Insights into Candida Biofilm Eradication Potential of Eucalyptol. J. Appl. Microbiol. 2021, 131, 105–123. [Google Scholar] [CrossRef]
- Mittal, R.P.; Rana, A.; Jaitak, V. Essential Oils: An Impending Substitute of Synthetic Antimicrobial Agents to Overcome Antimicrobial Resistance. CDT 2019, 20, 605–624. [Google Scholar] [CrossRef]
- Sun, J. D-Limonene: Safety and Clinical Applications. Altern. Med. Rev. 2007, 12, 259–264. [Google Scholar]
- Vieira, A.J.; Beserra, F.P.; Souza, M.C.; Totti, B.M.; Rozza, A.L. Limonene: Aroma of Innovation in Health and Disease. Chem. Biol. Interact. 2018, 283, 97–106. [Google Scholar] [CrossRef] [Green Version]
- Galdiero, E.; Di Onofrio, V.; Maione, A.; Gambino, E.; Gesuele, R.; Menale, B.; Ciaravolo, M.; Carraturo, F.; Guida, M. Allium Ursinum and Allium Oschaninii against Klebsiella Pneumoniae and Candida Albicans Mono- and Polymicrobic Biofilms in In Vitro Static and Dynamic Models. Microorganisms 2020, 8, 336. [Google Scholar] [CrossRef] [Green Version]
- Williams, D.L.; Smith, S.R.; Peterson, B.R.; Allyn, G.; Cadenas, L.; Epperson, R.T.; Looper, R.E. Growth Substrate May Influence Biofilm Susceptibility to Antibiotics. PLoS ONE 2019, 14, e0206774. [Google Scholar] [CrossRef] [Green Version]
- Grinberg, M.; Orevi, T.; Kashtan, N. Bacterial Surface Colonization, Preferential Attachment and Fitness under Periodic Stress. PLoS Comput. Biol. 2019, 15, e1006815. [Google Scholar] [CrossRef]
- Gajewska, J.; Chajęcka-Wierzchowska, W. Biofilm Formation Ability and Presence of Adhesion Genes among Coagulase-Negative and Coagulase-Positive Staphylococci Isolates from Raw Cow’s Milk. Pathogens 2020, 9, 654. [Google Scholar] [CrossRef]
- Mirza, R.; Azeem, M.; Qaisar, U. Influence of Peganum Harmala Peptides on the Transcriptional Activity of Biofilm Related Genes in Sensitive and Resistant Strains of Pseudomonas aeruginosa and Staphylococcus aureus. Pak. J. Pharm. Sci. 2019, 32, 2341–2345. [Google Scholar]
- Weinstein, M.P.; Patel, J.B. Methods for Dilution Antimicrobial Susceptibility Tests for Bacteria That Grow Aerobically: M07-A11, 11th ed.; Documents/Clinical and Laboratory Standards Institute; Committee for Clinical Laboratory Standards: Wayne, PA, USA, 2018. [Google Scholar]
- Goeres, D.M.; Loetterle, L.R.; Hamilton, M.A.; Murga, R.; Kirby, D.W.; Donlan, R.M. Statistical Assessment of a Laboratory Method for Growing Biofilms. Microbiology 2005, 151, 757–762. [Google Scholar] [CrossRef] [Green Version]
- Pfaffl, M.W. Relative Expression Software Tool (REST(C)) for Group-Wise Comparison and Statistical Analysis of Relative Expression Results in Real-Time PCR. Nucleic Acids Res. 2002, 30, e36. [Google Scholar] [CrossRef]
- Riesgo, A.; Pérez-Porro, A.R.; Carmona, S.; Leys, S.P.; Giribet, G. Optimization of Preservation and Storage Time of Sponge Tissues to Obtain Quality MRNA for Next-generation Sequencing. Mol. Ecol. Resour. 2012, 12, 312–322. [Google Scholar] [CrossRef]
- Savli, H.; Karadenizli, A.; Kolayli, F.; Gundes, S.; Ozbek, U.; Vahaboglu, H. Expression Stability of Six Housekeeping Genes: A Proposal for Resistance Gene Quantification Studies of Pseudomonas aeruginosa by Real-Time Quantitative RT-PCR. J. Med. Microbiol. 2003, 52, 403–408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ogonowska, P.; Nakonieczna, J. Validation of Stable Reference Genes in Staphylococcus aureus to Study Gene Expression under Photodynamic Treatment: A Case Study of SEB Virulence Factor Analysis. Sci. Rep. 2020, 10, 16354. [Google Scholar] [CrossRef] [PubMed]
- Alves, P.M.; Al-Badi, E.; Withycombe, C.; Jones, P.M.; Purdy, K.J.; Maddocks, S.E. Interaction between Staphylococcus aureus and Pseudomonas aeruginosa Is Beneficial for Colonisation and Pathogenicity in a Mixed Biofilm. Pathog. Dis. 2018, 76, fty003. [Google Scholar] [CrossRef] [PubMed]
- Lenhard, J.R.; Smith, N.M.; Quach, C.D.; Nguyen, T.Q.; Doan, L.H.; Chau, J. Bacterial Brothers in Arms: Cooperation of Staphylococcus aureus and Pseudomonas aeruginosa during Antimicrobial Exposure. J. Antimicrob. Chemother. 2019, 74, 2657–2665. [Google Scholar] [CrossRef]
- Chang, C.-Y. Surface Sensing for Biofilm Formation in Pseudomonas aeruginosa. Front. Microbiol. 2018, 8, 2671. [Google Scholar] [CrossRef]
- Zheng, Y.; He, L.; Asiamah, T.K.; Otto, M. Colonization of Medical Devices by Staphylococci: Colonization of Medical Devices by Staphylococci. Environ. Microbiol. 2018, 20, 3141–3153. [Google Scholar] [CrossRef] [Green Version]
- Doran, P.M. Application of Plant Tissue Cultures in Phytoremediation Research: Incentives and Limitations. Biotechnol. Bioeng. 2009, 103, 60–76. [Google Scholar] [CrossRef]
- Solórzano-Santos, F.; Miranda-Novales, M.G. Essential Oils from Aromatic Herbs as Antimicrobial Agents. Curr. Opin. Biotechnol. 2012, 23, 136–141. [Google Scholar] [CrossRef]
- Korenblum, E.; Regina de Vasconcelos Goulart, F.; de Almeida Rodrigues, I.; Abreu, F.; Lins, U.; Alves, P.; Blank, A.; Valoni, É.; Sebastián, G.V.; Alviano, D.; et al. Antimicrobial Action and Anti-Corrosion Effect against Sulfate Reducing Bacteria by Lemongrass (Cymbopogon Citratus) Essential Oil and Its Major Component, the Citral. AMB Express 2013, 3, 44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Bryan, C.A.; Crandall, P.G.; Chalova, V.I.; Ricke, S.C. Orange Essential Oils Antimicrobial Activities against Salmonella Spp. J. Food Sci. 2008, 73, M264–M267. [Google Scholar] [CrossRef]
- Ambrosio, C.M.S.; Ikeda, N.Y.; Miano, A.C.; Saldaña, E.; Moreno, A.M.; Stashenko, E.; Contreras-Castillo, C.J.; Da Gloria, E.M. Unraveling the Selective Antibacterial Activity and Chemical Composition of Citrus Essential Oils. Sci. Rep. 2019, 9, 17719. [Google Scholar] [CrossRef] [Green Version]
- Bhandari, D.P.; Poudel, D.K.; Satyal, P.; Khadayat, K.; Dhami, S.; Aryal, D.; Chaudhary, P.; Ghimire, A.; Parajuli, N. Volatile Compounds and Antioxidant and Antimicrobial Activities of Selected Citrus Essential Oils Originated from Nepal. Molecules 2021, 26, 6683. [Google Scholar] [CrossRef]
- Vázquez-Sánchez, D.; Galvão, J.A.; Mazine, M.R.; Gloria, E.M.; Oetterer, M. Control of Staphylococcus aureus Biofilms by the Application of Single and Combined Treatments Based in Plant Essential Oils. Int. J. Food Microbiol. 2018, 286, 128–138. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Names | Acronym | Primer Name | Sequence (5′→3′) | References |
|---|---|---|---|---|
| pilin gene | pilA | P. aeuriginosa_pilA_F | GATCGAACTGATGATCGTGGT | Mirza et al., 2019 [20] |
| P. aeuriginosa_pilA_R | GACATATGTTTCGGTCGCAGT | |||
| flagellar gene | flgK | P. aeuriginosa_flgK_F | CGATACCGTCAACAAGCAACT | Mirza et al., 2019 [20] |
| P. aeuriginosa_flgK_R | CTTGCTGGTATCGGTGATGTT | |||
| rpoS | rpoS | P. aeuriginosa_rpoS_F | CTCCCCGGGCAACTCCAAAAG | Savli et al. 2003 [25] |
| P. aeuriginosa_rpoS_R | CGATCATCCGCTTCCGACCAG | |||
| laminin-binding protein | eno | S. aureus_eno_F | ACGTGCAGCAGCTGACT | Mirza et al., 2019 [20] |
| S. aureus_eno_R | CAACAGCATCTTCAGTACCTTC | |||
| intercellular adhesion gene | icaA | S. aureus_icaA_F | CCAGAAACATTGGGAGGTCTT | Mirza et al., 2019 [20] |
| S. aureus_icaA_R | CCTTTTCGTTTTCATTGTGCT | |||
| 16S Rrna | 16S rRNA | S. aureus_16SrRNA_F | TATGGAGGAACACCAGTGGCGAAG | Ogonowska and Nakonieczn, 2020 [26] |
| S. aureus_16SrRNA_R | TCATCGTTTACGGCGTGGACTACC |
| MIC | MBC | MBC/MIC | ||
|---|---|---|---|---|
| mL/L | ratio | |||
| S. aureus | 20 | >40 | - | batteriostatic |
| P. aeruginosa | 40 | >40 | - | batteriostatic |
| Gene | Staphylococcus aureus + Pseudomonas aeruginosa | ||
|---|---|---|---|
| Polypropylene | Polycarbonate | Steel | |
| pilA | 5.91 | 7.99 | 4.21 |
| flgK | 2.16 | 4.18 | 2.27 |
| eno | 0.50 | −0.65 | −6.98 |
| icaA | 0.49 | −0.70 | −7.50 |
| Pseudomonas aeruginosa | |||
| Polypropylene | Polycarbonate | Steel | |
| pilA | −3.13 | 1.88 | −3.13 |
| flgK | −2.72 | 0.96 | −2.72 |
| Staphylococcus aureus | |||
| Polypropylene | Polycarbonate | Steel | |
| eno | −0.16 | 15.41 | 20.65 |
| icaA | −2.38 | 10.58 | 22.84 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gambino, E.; Maione, A.; Guida, M.; Albarano, L.; Carraturo, F.; Galdiero, E.; Di Onofrio, V. Evaluation of the Pathogenic-Mixed Biofilm Formation of Pseudomonas aeruginosa/Staphylococcus aureus and Treatment with Limonene on Three Different Materials by a Dynamic Model. Int. J. Environ. Res. Public Health 2022, 19, 3741. https://doi.org/10.3390/ijerph19063741
Gambino E, Maione A, Guida M, Albarano L, Carraturo F, Galdiero E, Di Onofrio V. Evaluation of the Pathogenic-Mixed Biofilm Formation of Pseudomonas aeruginosa/Staphylococcus aureus and Treatment with Limonene on Three Different Materials by a Dynamic Model. International Journal of Environmental Research and Public Health. 2022; 19(6):3741. https://doi.org/10.3390/ijerph19063741
Chicago/Turabian StyleGambino, Edvige, Angela Maione, Marco Guida, Luisa Albarano, Federica Carraturo, Emilia Galdiero, and Valeria Di Onofrio. 2022. "Evaluation of the Pathogenic-Mixed Biofilm Formation of Pseudomonas aeruginosa/Staphylococcus aureus and Treatment with Limonene on Three Different Materials by a Dynamic Model" International Journal of Environmental Research and Public Health 19, no. 6: 3741. https://doi.org/10.3390/ijerph19063741
APA StyleGambino, E., Maione, A., Guida, M., Albarano, L., Carraturo, F., Galdiero, E., & Di Onofrio, V. (2022). Evaluation of the Pathogenic-Mixed Biofilm Formation of Pseudomonas aeruginosa/Staphylococcus aureus and Treatment with Limonene on Three Different Materials by a Dynamic Model. International Journal of Environmental Research and Public Health, 19(6), 3741. https://doi.org/10.3390/ijerph19063741