
The Effects of Alcohol Drinking on Oral Microbiota in the Chinese Population

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Population and Saliva Sample Collection
2.2. DNA Extraction and 16S rRNA Gene Sequencing
2.3. Bioinformatic Analysis of 16S rRNA Sequencing
2.4. Statistical Analysis
3. Results
3.1. Characteristics of the Study Participants
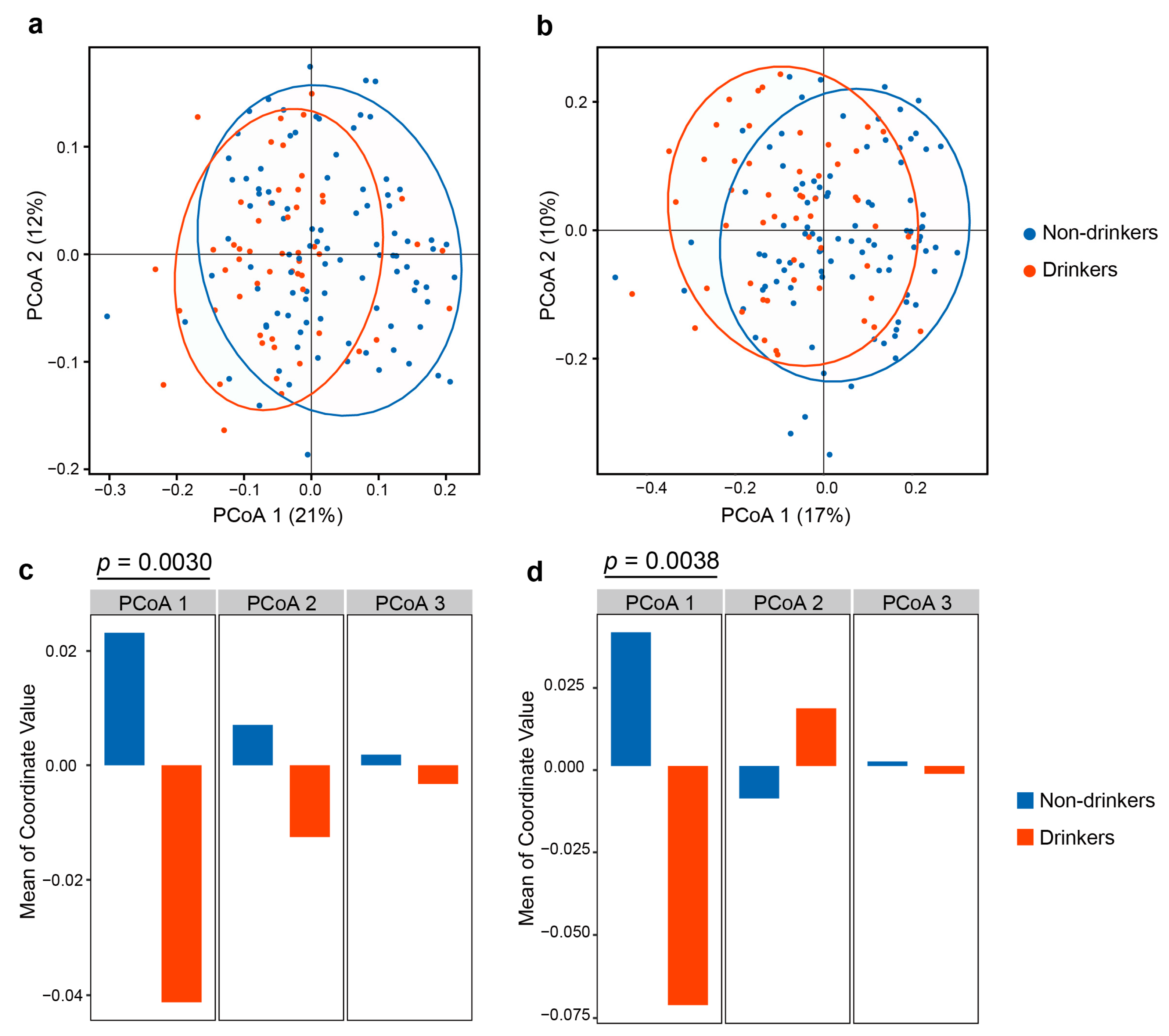
3.2. The Influence of Alcohol Drinking on Oral Microbial Diversity and Overall Composition
3.3. The Influence of Alcohol Drinking on Oral Microbial Taxa
3.4. Correlation between Oral Microbiota and Predictive Functional Pathways
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Koren, O.; Spor, A.; Felin, J.; Fåk, F.; Stombaugh, J.; Tremaroli, V.; Behre, C.J.; Knight, R.; Fagerberg, B.; Ley, R.E.; et al. Human oral, gut, and plaque microbiota in patients with atherosclerosis. Proc. Natl. Acad. Sci. USA 2011, 108 (Suppl. S1), 4592–4598. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galvão-Moreira, L.V.; da Cruz, M.C. Oral microbiome, periodontitis and risk of head and neck cancer. Oral Oncol. 2016, 53, 17–19. [Google Scholar] [CrossRef] [PubMed]
- Long, J.; Cai, Q.; Steinwandel, M.; Hargreaves, M.K.; Bordenstein, S.R.; Blot, W.J.; Zheng, W.; Shu, X.O. Association of oral microbiome with type 2 diabetes risk. J. Periodontal Res. 2017, 52, 636–643. [Google Scholar] [CrossRef] [PubMed]
- Poveda-Roda, R.; Jiménez, Y.; Carbonell, E.; Gavaldá, C.; Margaix-Muñoz, M.M.; Sarrión-Pérez, G. Bacteremia originating in the oral cavity. A review. Med. Oral Patol. Oral Cir. Bucal 2008, 13, E355–E362. [Google Scholar]
- Irfan, M.; Delgado, R.Z.R.; Frias-Lopez, J. The Oral Microbiome and Cancer. Front. Immunol. 2020, 11, 591088. [Google Scholar] [CrossRef]
- Takeuchi, T.; Miyauchi, E.; Kanaya, T.; Kato, T.; Nakanishi, Y.; Watanabe, T.; Kitami, T.; Taida, T.; Sasaki, T.; Negishi, H.; et al. Acetate differentially regulates IgA reactivity to commensal bacteria. Nature 2021, 595, 560–564. [Google Scholar] [CrossRef]
- Griswold, M.G.; Fullman, N.; Hawley, C.; Arian, N.; Zimsen, S.R.M.; Tymeson, H.D.; Venkateswaran, V.; Tapp, A.D.; Forouzanfar, M.H.; Salama, J.S.; et al. Alcohol use and burden for 195 countries and territories, 1990–2016: A systematic analysis for the Global Burden of Disease Study 2016. Lancet 2018, 392, 1015–1035. [Google Scholar] [CrossRef] [Green Version]
- Lages, E.J.; Costa, F.O.; Cortelli, S.C.; Cortelli, J.R.; Cota, L.O.; Cyrino, R.M.; Lages, E.M.; Nobre-Franco, G.C.; Brito, J.A.; Gomez, R.S. Alcohol Consumption and Periodontitis: Quantification of Periodontal Pathogens and Cytokines. J. Periodontol. 2015, 86, 1058–1068. [Google Scholar] [CrossRef]
- Enberg, N.; Alho, H.; Loimaranta, V.; Lenander-Lumikari, M. Saliva flow rate, amylase activity, and protein and electrolyte concentrations in saliva after acute alcohol consumption. Oral Surg. Oral Med. Oral Pathol. Oral Radiol. Endod. 2001, 92, 292–298. [Google Scholar] [CrossRef]
- Ingram, L.O. Ethanol tolerance in bacteria. Crit. Rev. Biotechnol. 1990, 9, 305–319. [Google Scholar] [CrossRef]
- Homann, N.; Tillonen, J.; Meurman, J.H.; Rintamäki, H.; Lindqvist, C.; Rautio, M.; Jousimies-Somer, H.; Salaspuro, M. Increased salivary acetaldehyde levels in heavy drinkers and smokers: A microbiological approach to oral cavity cancer. Carcinogenesis 2000, 21, 663–668. [Google Scholar] [CrossRef] [PubMed]
- Fan, X.; Peters, B.A.; Jacobs, E.J.; Gapstur, S.M.; Purdue, M.P.; Freedman, N.D.; Alekseyenko, A.V.; Wu, J.; Yang, L.; Pei, Z.; et al. Drinking alcohol is associated with variation in the human oral microbiome in a large study of American adults. Microbiome 2018, 6, 59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Signoretto, C.; Bianchi, F.; Burlacchini, G.; Sivieri, F.; Spratt, D.; Canepari, P. Drinking habits are associated with changes in the dental plaque microbial community. J. Clin. Microbiol. 2010, 48, 347–356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomas, A.M.; Gleber-Netto, F.O.; Fernandes, G.R.; Amorim, M.; Barbosa, L.F.; Francisco, A.L.; de Andrade, A.G.; Setubal, J.C.; Kowalski, L.P.; Nunes, D.N.; et al. Alcohol and tobacco consumption affects bacterial richness in oral cavity mucosa biofilms. BMC Microbiol. 2014, 14, 250. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez-Rabassa, M.; López, P.; Sánchez, R.; Hernández, C.; Rodríguez, C.; Rodríguez-Santiago, R.E.; Orengo, J.C.; Green, V.; Yamamura, Y.; Rivera-Amill, V. Inflammatory Biomarkers, Microbiome, Depression, and Executive Dysfunction in Alcohol Users. Int. J. Environ. Res. Public Health 2020, 17, 689. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Quinque, D.; Horz, H.P.; Li, M.; Rzhetskaya, M.; Raff, J.A.; Hayes, M.G.; Stoneking, M. Comparative analysis of the human saliva microbiome from different climate zones: Alaska, Germany, and Africa. BMC Microbiol. 2014, 14, 316. [Google Scholar] [CrossRef] [Green Version]
- He, Y.Q.; Liao, X.Y.; Xue, W.Q.; Xu, Y.F.; Xu, F.H.; Li, F.F.; Li, X.Z.; Zhang, J.B.; Wang, T.M.; Wang, F.; et al. Association Between Environmental Factors and Oral Epstein-Barr Virus DNA Loads: A Multicenter Cross-sectional Study in China. J. Infect. Dis. 2019, 219, 400–409. [Google Scholar] [CrossRef]
- Jia, Y.J.; Liao, Y.; He, Y.Q.; Zheng, M.Q.; Tong, X.T.; Xue, W.Q.; Zhang, J.B.; Yuan, L.L.; Zhang, W.L.; Jia, W.H. Association Between Oral Microbiota and Cigarette Smoking in the Chinese Population. Front. Cell. Infect. Microbiol. 2021, 11, 658203. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Lauber, C.L.; Walters, W.A.; Berg-Lyons, D.; Lozupone, C.A.; Turnbaugh, P.J.; Fierer, N.; Knight, R. Global patterns of 16S rRNA diversity at a depth of millions of sequences per sample. Proc. Natl. Acad. Sci. USA 2011, 108 (Suppl. S1), 4516–4522. [Google Scholar] [CrossRef] [Green Version]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McDonald, D.; Price, M.N.; Goodrich, J.; Nawrocki, E.P.; DeSantis, T.Z.; Probst, A.; Andersen, G.L.; Knight, R.; Hugenholtz, P. An improved Greengenes taxonomy with explicit ranks for ecological and evolutionary analyses of bacteria and archaea. ISME J. 2012, 6, 610–618. [Google Scholar] [CrossRef] [PubMed]
- Lozupone, C.A.; Knight, R. Species divergence and the measurement of microbial diversity. FEMS Microbiol. Rev. 2008, 32, 557–578. [Google Scholar] [CrossRef] [PubMed]
- Lozupone, C.A.; Hamady, M.; Kelley, S.T.; Knight, R. Quantitative and qualitative beta diversity measures lead to different insights into factors that structure microbial communities. Appl. Environ. Microbiol. 2007, 73, 1576–1585. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Langille, M.G.; Zaneveld, J.; Caporaso, J.G.; McDonald, D.; Knights, D.; Reyes, J.A.; Clemente, J.C.; Burkepile, D.E.; Vega Thurber, R.L.; Knight, R.; et al. Predictive functional profiling of microbial communities using 16S rRNA marker gene sequences. Nat. Biotechnol. 2013, 31, 814–821. [Google Scholar] [CrossRef]
- Segata, N.; Izard, J.; Waldron, L.; Gevers, D.; Miropolsky, L.; Garrett, W.S.; Huttenhower, C. Metagenomic biomarker discovery and explanation. Genome Biol. 2011, 12, R60. [Google Scholar] [CrossRef] [Green Version]
- Nearing, J.T.; DeClercq, V.; Van Limbergen, J.; Langille, M.G.I. Assessing the Variation within the Oral Microbiome of Healthy Adults. mSphere 2020, 5, e00451-20. [Google Scholar] [CrossRef]
- Jansson, L. Association between alcohol consumption and dental health. J. Clin. Periodontol. 2008, 35, 379–384. [Google Scholar] [CrossRef]
- Wang, J.; Lv, J.; Wang, W.; Jiang, X. Alcohol consumption and risk of periodontitis: A meta-analysis. J. Clin. Periodontol. 2016, 43, 572–583. [Google Scholar] [CrossRef]
- Barb, J.J.; Maki, K.A.; Kazmi, N.; Meeks, B.K.; Krumlauf, M.; Tuason, R.T.; Brooks, A.T.; Ames, N.J.; Goldman, D.; Wallen, G.R. The oral microbiome in alcohol use disorder: A longitudinal analysis during inpatient treatment. J. Oral Microbiol. 2022, 14, 2004790. [Google Scholar] [CrossRef] [PubMed]
- Marsh, P.D.; Do, T.; Beighton, D.; Devine, D.A. Influence of saliva on the oral microbiota. Periodontology 2000 2016, 70, 80–92. [Google Scholar] [CrossRef] [PubMed]
- Bluemel, S.; Wang, L.; Kuelbs, C.; Moncera, K.; Torralba, M.; Singh, H.; Fouts, D.E.; Schnabl, B. Intestinal and hepatic microbiota changes associated with chronic ethanol administration in mice. Gut Microbes 2020, 11, 265–275. [Google Scholar] [CrossRef] [PubMed]
- Kwan, S.Y.; Jiao, J.; Joon, A.; Wei, P.; Petty, L.E.; Below, J.E.; Daniel, C.R.; Wu, X.; Zhang, J.; Jenq, R.R.; et al. Gut microbiome features associated with liver fibrosis in Hispanics, a population at high risk for fatty liver disease. Hepatology 2021, 75, 955–967. [Google Scholar] [CrossRef] [PubMed]
- Scher, J.U.; Sczesnak, A.; Longman, R.S.; Segata, N.; Ubeda, C.; Bielski, C.; Rostron, T.; Cerundolo, V.; Pamer, E.G.; Abramson, S.B.; et al. Expansion of intestinal Prevotella copri correlates with enhanced susceptibility to arthritis. eLife 2013, 2, e01202. [Google Scholar] [CrossRef] [PubMed]
- Zhao-Fleming, H.H.; Barake, S.R.S.; Hand, A.; Wilkinson, J.E.; Sanford, N.; Winn, R.; Dissanaike, S. Traditional culture methods fail to detect principle pathogens in necrotising soft tissue infection: A case report. J. Wound Care 2018, 27, S24–S28. [Google Scholar] [CrossRef] [PubMed]
- Bein, T.; Brem, J.; Schüsselbauer, T. Bacteremia and sepsis due to Prevotella oris from dentoalveolar abscesses. Intensive Care Med. 2003, 29, 856. [Google Scholar] [CrossRef]
- Teanpaisan, R.; Douglas, C.W.; Eley, A.R.; Walsh, T.F. Clonality of Porphyromonas gingivalis, Prevotella intermedia and Prevotella nigrescens isolated from periodontally diseased and healthy sites. J. Periodontal Res. 1996, 31, 423–432. [Google Scholar] [CrossRef]
- Baumgartner, J.C.; Watkins, B.J.; Bae, K.S.; Xia, T. Association of black-pigmented bacteria with endodontic infections. J. Endod. 1999, 25, 413–415. [Google Scholar] [CrossRef]
- Si, J.; You, H.J.; Yu, J.; Sung, J.; Ko, G. Prevotella as a Hub for Vaginal Microbiota under the Influence of Host Genetics and Their Association with Obesity. Cell Host Microbe 2017, 21, 97–105. [Google Scholar] [CrossRef] [Green Version]
- Mager, D.L.; Haffajee, A.D.; Devlin, P.M.; Norris, C.M.; Posner, M.R.; Goodson, J.M. The salivary microbiota as a diagnostic indicator of oral cancer: A descriptive, non-randomized study of cancer-free and oral squamous cell carcinoma subjects. J. Transl. Med. 2005, 3, 27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karpiński, T.M. Role of Oral Microbiota in Cancer Development. Microorganisms 2019, 7, 20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tett, A.; Pasolli, E.; Masetti, G.; Ercolini, D.; Segata, N. Prevotella diversity, niches and interactions with the human host. Nat. Rev. Microbiol. 2021, 19, 585–599. [Google Scholar] [CrossRef] [PubMed]
- Larsen, J.M. The immune response to Prevotella bacteria in chronic inflammatory disease. Immunology 2017, 151, 363–374. [Google Scholar] [CrossRef] [Green Version]
- Kjær Hansen, S.; Løfberg, S.V.; Nielsen, D.K.; Kobberø, H.; Justesen, U.S. Bacteraemia with Moryella indoligenes and Fastidiosipila sanguinis: A case report. Access Microbiol. 2020, 2, acmi000108. [Google Scholar] [CrossRef]
- Tsementzi, D.; Pena-Gonzalez, A.; Bai, J.; Hu, Y.J.; Patel, P.; Shelton, J.; Dolan, M.; Arluck, J.; Khanna, N.; Conrad, L.; et al. Comparison of vaginal microbiota in gynecologic cancer patients pre- and post-radiation therapy and healthy women. Cancer Med. 2020, 9, 3714–3724. [Google Scholar] [CrossRef] [Green Version]
- Al-Hebshi, N.N.; Nasher, A.T.; Maryoud, M.Y.; Homeida, H.E.; Chen, T.; Idris, A.M.; Johnson, N.W. Inflammatory bacteriome featuring Fusobacterium nucleatum and Pseudomonas aeruginosa identified in association with oral squamous cell carcinoma. Sci. Rep. 2017, 7, 1834. [Google Scholar] [CrossRef] [Green Version]
- Baraniya, D.; Jain, V.; Lucarelli, R.; Tam, V.; Vanderveer, L.; Puri, S.; Yang, M.; Al-Hebshi, N.N. Screening of Health-Associated Oral Bacteria for Anticancer Properties in vitro. Front. Cell. Infect. Microbiol. 2020, 10, 575656. [Google Scholar] [CrossRef]
- Valles-Colomer, M.; Bacigalupe, R.; Vieira-Silva, S.; Suzuki, S.; Darzi, Y.; Tito, R.Y.; Yamada, T.; Segata, N.; Raes, J.; Falony, G. Variation and transmission of the human gut microbiota across multiple familial generations. Nat. Microbiol. 2022, 7, 87–96. [Google Scholar] [CrossRef]
- Adeva-Andany, M.; López-Ojén, M.; Funcasta-Calderón, R.; Ameneiros-Rodríguez, E.; Donapetry-García, C.; Vila-Altesor, M.; Rodríguez-Seijas, J. Comprehensive review on lactate metabolism in human health. Mitochondrion 2014, 17, 76–100. [Google Scholar] [CrossRef]
- Lamont, R.J.; Koo, H.; Hajishengallis, G. The oral microbiota: Dynamic communities and host interactions. Nat. Rev. Microbiol. 2018, 16, 745–759. [Google Scholar] [CrossRef] [PubMed]
- Jing, X.; Yang, F.; Shao, C.; Wei, K.; Xie, M.; Shen, H.; Shu, Y. Role of hypoxia in cancer therapy by regulating the tumor microenvironment. Mol. Cancer 2019, 18, 157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sedghi, L.; DiMassa, V.; Harrington, A.; Lynch, S.V.; Kapila, Y.L. The oral microbiome: Role of key organisms and complex networks in oral health and disease. Periodontology 2000 2021, 87, 107–131. [Google Scholar] [CrossRef] [PubMed]
- Ames, N.J.; Barb, J.J.; Schuebel, K.; Mudra, S.; Meeks, B.K.; Tuason, R.T.S.; Brooks, A.T.; Kazmi, N.; Yang, S.; Ratteree, K.; et al. Longitudinal gut microbiome changes in alcohol use disorder are influenced by abstinence and drinking quantity. Gut Microbes 2020, 11, 1608–1631. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
| Non-Drinkers (N = 96) | Drinkers (N = 54) | p-Value | |
|---|---|---|---|
| Age, mean (s.d.) | 45.63 (9.73) | 49.24 (11.04) | 0.05 1 |
| Sex, n (%) | <0.01 2 | ||
| Male | 58 (60.42) | 48 (88.89) | |
| Female | 38 (39.58) | 6 (11.11) | |
| Education, n (%) | 0.68 2 | ||
| <High school | 77 (80.21) | 41 (75.93) | |
| ≥High school | 19 (19.79) | 13 (24.07) | |
| Smoking status, n (%) | <0.001 2 | ||
| Non-current | 61 (63.54) | 15 (27.78) | |
| Current | 35 (36.46) | 39 (72.22) | |
| Teeth loss after age 20, n (%) | 0.13 2 | ||
| Yes | 45 (46.88) | 33 (61.11) | |
| No | 51 (53.12) | 21 (38.89) | |
| The number of teeth lost after age 20, mean(s.d.) | 3.33 (7.08) | 3.54 (6.15) | 0.85 1 |
| Tooth brushing frequency, n (%) | 0.84 2 | ||
| ≤1 time a day | 61 (63.54) | 36 (66.67) | |
| ≥2 times a day | 35 (36.46) | 18 (33.33) |
| Taxonomy | Mean Count 1 | log2Fc (95% CI) 3 | p-Value 2 | |
|---|---|---|---|---|
| Non-Drinkers (N = 96) | Drinkers (N = 54) | |||
| Genus level | ||||
| Prevotella | 1255.23 | 1810.96 | 0.47 (0.16, 0.78) | 0.0033 |
| Moryella | 17.05 | 32.07 | 0.58 (0.02, 1.14) | 0.042 |
| Lautropia | 107.90 | 65.59 | −0.82 (−1.37, −0.26) | 0.0039 |
| Haemophilus | 780.65 | 598.50 | −0.31 (−0.54, −0.08) | 0.0093 |
| Porphyromonas | 557.63 | 488.76 | −0.29 (−0.56, −0.02) | 0.036 |
| Species level | ||||
| Prevotella melaninogenica | 447.50 | 805.52 | 0.68 (0.26, 1.10) | 0.0016 |
| Prevotella tannerae | 53.61 | 109.00 | 0.87 (0.31, 1.43) | 0.0024 |
| Haemophilus parainfluenzae | 726.85 | 567.28 | −0.29 (−0.54, −0.04) | 0.021 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liao, Y.; Tong, X.-T.; Jia, Y.-J.; Liu, Q.-Y.; Wu, Y.-X.; Xue, W.-Q.; He, Y.-Q.; Wang, T.-M.; Zheng, X.-H.; Zheng, M.-Q.; et al. The Effects of Alcohol Drinking on Oral Microbiota in the Chinese Population. Int. J. Environ. Res. Public Health 2022, 19, 5729. https://doi.org/10.3390/ijerph19095729
Liao Y, Tong X-T, Jia Y-J, Liu Q-Y, Wu Y-X, Xue W-Q, He Y-Q, Wang T-M, Zheng X-H, Zheng M-Q, et al. The Effects of Alcohol Drinking on Oral Microbiota in the Chinese Population. International Journal of Environmental Research and Public Health. 2022; 19(9):5729. https://doi.org/10.3390/ijerph19095729
Chicago/Turabian StyleLiao, Ying, Xia-Ting Tong, Yi-Jing Jia, Qiao-Yun Liu, Yan-Xia Wu, Wen-Qiong Xue, Yong-Qiao He, Tong-Min Wang, Xiao-Hui Zheng, Mei-Qi Zheng, and et al. 2022. "The Effects of Alcohol Drinking on Oral Microbiota in the Chinese Population" International Journal of Environmental Research and Public Health 19, no. 9: 5729. https://doi.org/10.3390/ijerph19095729
APA StyleLiao, Y., Tong, X.-T., Jia, Y.-J., Liu, Q.-Y., Wu, Y.-X., Xue, W.-Q., He, Y.-Q., Wang, T.-M., Zheng, X.-H., Zheng, M.-Q., & Jia, W.-H. (2022). The Effects of Alcohol Drinking on Oral Microbiota in the Chinese Population. International Journal of Environmental Research and Public Health, 19(9), 5729. https://doi.org/10.3390/ijerph19095729