
Potential Role of Glyphosate, Glyphosate-Based Herbicides, and AMPA in Breast Cancer Development: A Review of Human and Human Cell-Based Studies


Abstract
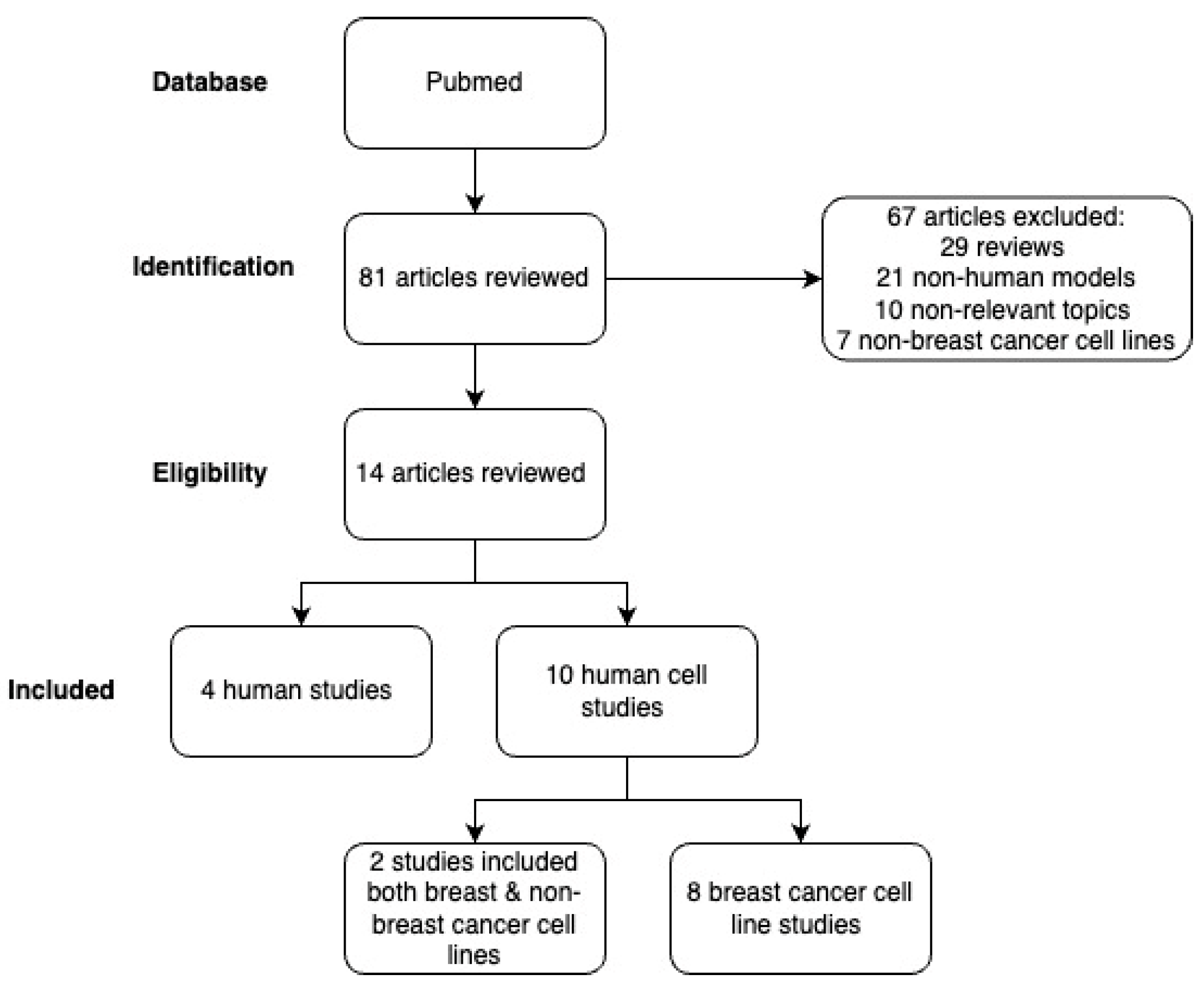
:1. Introduction
2. Materials and Methods
2.1. Inclusion/Exclusion Criteria
2.2. Search Strategy
2.3. Data Extraction
2.4. Outcome Measures
3. Results
3.1. Human Studies

{kind=link}
| First Author, Year Published | Study Design, Setting, Population, Period | Exposure Assessment | Glyphosate, AMPA, and/or GBH(s) | Method (LOD/LOQ), Corrections for Urine Concentration | Exposure Range/Median | Outcome Measures | Results | Strengths | Weaknesses |
|---|---|---|---|---|---|---|---|---|---|
| Eaton et al., 2022 [30] | Prospective cohort study; Northern Karst region of Puerto Rico; 205 pregnant women (18–40 years old) from the PROTECT birth cohort; 2012–2017 | Urinary Gly and AMPA levels at 16–20 (Visit 1) and 24–28 (Visit 3) weeks gestation | Gly and AMPA | GC-MS/MS (LOD: 0:20 µg/L), corrected for specific gravity | Median Gly level was 0.50 ng/mL (IQR: 0.29, 0.79); median AMPA level was 0.26 ng/mL (IQR: 0.17, 0.49). | Levels of oxidative stress biomarkers | An IQR increase in urinary AMPA was associated with higher 8-iso-PGF2α metabolite levels in mixed-effects model (6.71% [95% CI 1.51–12.17%]) and at visit 3 specifically (9.03% [95% CI 0.55–18.21%]). An IQR increase in Gly was associated with higher 8-iso-PGF2α levels only at visit 3 (7.23% [95% CI 0.11–14.86%]). | One of few studies to measure human exposure to both Gly and AMPA. First human study to look at association between AMPA and oxidative stress biomarkers. The use of two time points allowed for greater statistical power and determination of susceptibility periods. | Observational study so causality is indeterminable. The 8-iso-PGF2α/PGF2α ratio method used to show differences between oxidative stress and inflammation may not be an accurate representation. |
| Franke et al., 2021 [28] | Nested case–control pilot study; Hawaii; 250 women (45–75 years old) from the Multiethnic Cohort; 2001–2006 | Overnight urinary AMPA concentrations | AMPA | LC/HRAM-MS (LLOQ: 0.001 ng/mL), uncorrected and corrected for urinary creatinine | Urinary AMPA levels ranged from <LLOQ to 3698 ng/L. Adjusted geometric mean in cases was 0.087 ng/mg (95% CI 0.055–0.119); in controls was 0.063 ng/mg (95% CI 0.032–0.095). | Incident breast cancer | Urinary AMPA was associated with increased breast cancer risk. Highest vs. lowest quintile—OR 4.49; 95% CI 1.46–13.77 Second vs. lowest quintile—OR 3.03; 95% CI 1.02–9.03 | First prospective study to look at associations between urinary AMPA levels and breast cancer risk (measured by liquid chromatography mass spectrometry). | Observational study so causality is indeterminable. Used only one urine sample, may not be representative of usual AMPA exposure. |
| Lesseur et al., 2021 [9] | Pilot study nested within prospective cohort; UCSF, URMC, UMN, UW; 94 pregnant women and their term infants from The Infant Development and the Environment Study (TIDES) pregnancy cohort; August 2010–August 2012 | Urinary Gly and AMPA levels in 2nd trimester mothers | Gly and AMPA | UPLC-MS/MS (Gly LOD: 0.014 ng/mL; LOQ: 0.041 ng/mL), (AMPA LOD: 0.013 ng/mL; LOQ: 0.04 ng/mL), corrected for specific gravity | Urinary Gly levels ranged from 0.01 to 1.9 ng/mL (median: 0.22 ng/mL); AMPA levels ranged from 0.01 to 6 ng/mL (median: 0.14 ng/mL). | Anogenital distance (AGD) in newborns | Maternal urinary Gly was associated with longer (unadjusted) AGD-AC (p = 0.05). Maternal urinary AMPA was associated with longer AGD-AF (p = 0.01). | First study to look at association between maternal urinary Gly/AMPA levels and AGD in human newborns. Multicenter TIDES study included data from various geographic areas of the US. | Observational study so causality is indeterminable. Only one urine sample in 2nd trimester, which does not correspond with masculinization programming window. |
| Lucia et al., 2022 [11] | Cross-sectional study; Southern California; 392 postmenopausal women aged 45 to 66 years; 2017–2019 | First morning urinary Gly and AMPA levels on 2 days within a 10-day period | Gly and AMPA | LC-MS/MS (Gly—LOD: 0.014 ng/mL; LOQ: 0.041 ng/mL), (AMPA—LOD: 0.013 ng/mL; LOQ: 0.040 ng/mL), adjusted for urinary creatinine | Median Gly level was 0.12 ng/mL (IQR: 0.06, 0.22) and median AMPA level was 0.06 ng/mL (IQR: 0.02, 0.12). | DNA methylation level at >850,000 CpG sites | Urinary AMPA was associated with increased epigenetic age acceleration (p = 0.04). Urinary Gly was associated with DNA methylation of regions in the promoters of MSH4, KCNA6, ABAT, and NDUFAF2/ERCC8. Urinary AMPA was associated with DNA methylation in the ESR1 promoter. | Largest study on urinary Gly and AMPA levels in a nonagricultural setting. Most recent practices were used to analyze DNA methylation. Two urine samples were used per participant to provide a better representation of usual Gly/AMPA exposure. | Observational study so causality is indeterminable. Also indeterminable if differences in DNA methylation will affect gene expression due to lack of gene expression data. |
3.2. Human Breast Cell Studies
| First Author, Year Published | Chemical Used | Cell Line(s) | Treatment | Outcome Measures | Assays/Techniques Used | Main Findings | Proposed Mechanisms |
|---|---|---|---|---|---|---|---|
| Antoniou et al., 2019 [53] | Gly | MDA-MB-231 | 100 mg/L Gly for 6 days | Global proteome changes; presence of glyoxylate-modified cysteines or glycine to Gly substitutions | RPLC-MS/MS/MS | No global proteome changes; Gly did not substitute for glycine nor did glyoxylation occur in proteins. | Potential Gly toxicity is not through glycine substitution. |
| Coppola et al., 2022 [41] | Gly | MCF-7 and MCF-12A | For cell viability, apoptosis/necrosis, and ATP levels: 230 pM, 2.3 nM, 23 nM, 230 nM, or 2.3 µM of Gly For other outcomes: 2.3, 23, or 230 nM of Gly | Cell viability; cell proliferation; apoptosis and necrosis; ATP levels; intracellular ROS levels; estradiol (E2) secretion; gene expression of nuclear receptors (ERα, ERβ, AR, AhR and PgR) | MTS; CyQuant; RealTime-Glo Annexin V Apoptosis and Necrosis; Mitochondrial ToxGlo; ROS Detection; Estradiol ELISA; real-time PCR | MCF-7: Gly reduced cell viability at 2.3 nM, 230 nM, and 2.3 µM and decreased cell proliferation at 230 pM; 230 nM and 2.3 µM decreased intracellular ATP levels; 2.3 nM increased E2 secretion; 2.3 nM downregulated ERα and Erβ, whereas 23 nM upregulated ERα; 23 and 230 nM upregulated AR; 2.3 and 230 nM downregulated PgR; 23 nM upregulated AhR MCF-12A: Gly reduced cell proliferation at 23 nM, 230 nM, and 2.3 µM; 2.3 nM increased apoptosis at 7 and 8 h; all concentrations reduced ROS; 230 nM increased E2 secretion; all concentrations (2.3, 23, and 230 nM) upregulated ERα and ERβ and downregulated PgR; 23 and 230 nM downregulated AR; 2.3 and 23 nM downregulated AhR | Gly may increase mitochondrial membrane permeability, increasing intracellular calcium concentrations and thus reducing ATP synthesis. Gly might alter balance of ERα and ERβ receptors, affecting mammary gland development. Gly increased E2 secretion, which could activate vascular endothelial growth factor transcription, and may lead to mammary gland angiogenesis. |
| De Almeida et al., 2018 [45] | Gly, Roundup, and Wipeout | MCF7 and MDA-MB-231 | 0–500 µg/mL of Gly, Roundup (Ro), or Wipeout (Wo) for 24 h | Cell viability and genotoxicity | MTT; comet | Gly (500, 1000 µg/mL), Ro (500, 800 µg/mL), and Wo (500 µg/mL) induced DNA damage in MDA-MB-231. No cytotoxic effects were observed for MCF7 and MDA-MB-231. | Gly, Ro, and Wo might cause toxic effects through non-estrogenic mechanisms. |
| Gasnier et al., 2009 [47] | Gly and 4 Roundup (R) formulations | MDA-MB453-kb2 | 2% solutions (and consecutive dilutions up to 10−7) of 1 of 4 R formulations (7.2 g/L, 360 g/L, 400 g/L, 450 g/L of Gly) or Gly alone (360 g/L) for 24 h | Anti-androgenic effects (on androgen receptor) | Luciferase reporter gene | All R formulations and Gly lowered DHT-mediated transcriptional activity. | Gly might bind to a steroid receptor. Adjuvants might form vesicles that intensify Gly effects by enhancing stability, cell penetration, and bioavailability. |
| Hokanson et al., 2007 [48] | Gly | MCF-7 | Gly at 0.1, 0.01, 0.001 or 0.0001% dilutions of the 15% stock for 18 h with or without 3 × 10−10 M E2 | Gene expression | DNA microarray; quantitative real-time PCR | At 0.00023% Gly, HIF1 was upregulated (more than twofold) and CXCL12 and EGR1 downregulated (more than 50%). Gly plus estrogen had greater effects than estrogen alone. | Gly and estrogen might synergistically affect gene expression, potentially damaging adult and fetal cells. Altered levels of EGR1, HIF1, and CXCL12 may initiate apoptosis, increase tumor angiogenesis, inactivate tumor suppressor genes, and disrupt immune surveillance. |
| Lin and Garry, 2000 [44] | Gly and Roundup | MCF-7 | Different concentrations that included up to 10 µg/mL for 72 h (cell viability) or 7 days (cell proliferation) | Cell viability; cell proliferation; apoptosis | FACS | Gly (0.228–2.28 µg/mL) and Roundup (1–10 µg/mL) induced cell proliferation in CD-treated (lacking estrogen) and non-CD-treated cultures. At cytotoxic concentrations, they induced necrosis (reduced cell viability and loss of cell membrane integrity). | Gly and Roundup might use a non-estrogenic mechanism to induce cell proliferation. Dependence on estrogenic mechanism is unknown for cytotoxic effects. |
| Mesnage et al., 2017 [42] | Gly, 4 GBH formulations, POEA (adjuvant) | MCF-7, T47D, T47D-KBluc, and MDA-MB-231 | 10−6 to 107 μg/L of Gly, GBH, or adjuvants for 6 days (ER-mediated cell proliferation), 24 h (ERE-mediated transcription), or 48 h (transcriptomics analysis) | ER-mediated cell proliferation (MCF-7, T47D, MDA-MB-231); transcriptomics analysis (MCF-7); ERE-mediated transcription (T47D-KBluc); molecular dynamic simulations | E-screen-MTT; ERE-luciferase reporter gene; microarray; RNA sequencing | Gly ≥ 10,000 μg/L promoted proliferation of MCF-7 cells and T47D less so; no effect in MDA-MB-231. Gly, but not Roundup nor POEA, increased ERE-mediated expression at ≥1000 μg/L; ER antagonist ICI 182,780 blocked this. Gly altered MCF-7 gene expression but not through ERα activation. Gly binding energy calculation predicts a weak and unstable interaction with ERα‘s active site. | Gly might activate ERα via a ligand-independent mechanism, possibly through the PKA pathway, but only at relatively high concentrations so humans exposed to Gly at typical levels would not be expected to exhibit ER activation. |
| Stur et al., 2019 [40] | AMPA and Roundup | MDA-MB-468 and MCF-7 | 0.01 to 10 mM of AMPA or 0.01% to 0.3% of Roundup for 3, 15, 24, and 48 h | Cell viability; gene expression | MTT; microarray | Roundup was more toxic than AMPA. After 48 h of 0.05% Roundup (1.1 mM Gly) exposure, 11 canonical pathways * were deregulated in both cell lines, including a more pronounced downregulation of cyclins and DNA damage repair pathways in MCF-7. 48 h of 10 mM AMPA exposure resulted in less differentially expressed genes, with most associated with metabolism in MDA-MB-468. | Roundup may deregulate ER-independent pathways related to cell cycle, DNA repair, and metabolism, which could change mitochondrial oxygen consumption, cause hypoxia, increase ROS, prevent DNA repair resulting in mutation buildup, and induce cell death. |
| Thongprakaisang et al., 2013 [43] | Gly | T47D, T47D-KBluc, and MDA-MB-231 | 10−12 to 10−6 M of Gly or estradiol (positive control) | Cell viability/number; estrogenicity and anti-estrogenicity of Gly (estrogen response element (ERE) transcription activity); ERα and ERβ expression | MTT; ERE-luciferase reporter gene; Western blot | Without E2 in the medium, Gly resulted in a 15–30% proliferation of T47D cells. Gly had no effect on the growth of hormone-independent MDA-MB-231 cells both in the absence/presence of E2. 1 nM of ER antagonist ICI 182780 weakened E2’s and Gly’s proliferative effects, while 10 nM completely inhibited the latter. In T47D-KBluc cells, Gly induced ERE activation 5–13-fold. When co-incubated, Gly suppressed E2-induced ERE activation. In T47D cells, Gly increased ERα and ERβ levels in a dose-dependent manner after 6 h exposure, but only ERα levels increased at the highest Gly concentration (10−7 M) after 24 h exposure. | Gly’s proliferative and stimulatory effects may occur via ER signaling since an ER antagonist inhibited this proliferation. At the ligand site of ERs, hydrophilic Gly may bind in a polar pocket. When the endogenous agonist E2 is present, Gly acts as an antagonist. Gly acts like a weak xenoestrogen that quickly activated ERβ while ERα activation was slower and longer. |
| Zhang et al., 2020 [46] | Gly | MELN | 500, 1000, and 1500 nM Gly | Aromatase activity; estrogenic activity; molecular dynamics | ELISA; luciferase reporter gene | Gly inhibited up to 30% of aromatase activity in a dose-dependent manner but did not interfere in the binding between E2 and ERα nor increase estrogen-responsive transcription. | Gly inhibits aromatase by potentially binding to an allosteric site. |
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| Gly | glyphosate |
| GBH | glyphosate-based herbicide |
| AMPA | aminomethylphosphonic acid |
References
- Benbrook, C.M. Trends in Glyphosate Herbicide Use in the United States and Globally. Environ. Sci. Eur. 2016, 28, 3. [Google Scholar] [CrossRef]
- Gaines, T.A. The Importance of Glyphosate in Non-GM Settings. Outlooks Pest Manag. 2018, 29, 255–257. [Google Scholar] [CrossRef]
- Herrmann, K.M.; Weaver, L.M. THE SHIKIMATE PATHWAY. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1999, 50, 473–503. [Google Scholar] [CrossRef] [PubMed]
- Medina-Pastor, P.; Triacchini, G. The 2018 European Union Report on Pesticide Residues in Food. EFSA J. 2020, 18, e06057. [Google Scholar] [CrossRef]
- Kolakowski, B.M.; Miller, L.; Murray, A.; Leclair, A.; Bietlot, H.; van de Riet, J.M. Analysis of Glyphosate Residues in Foods from the Canadian Retail Markets between 2015 and 2017. J. Agric. Food Chem. 2020, 68, 5201–5211. [Google Scholar] [CrossRef] [PubMed]
- Zoller, O.; Rhyn, P.; Rupp, H.; Zarn, J.A.; Geiser, C. Glyphosate Residues in Swiss Market Foods: Monitoring and Risk Evaluation. Food Addit. Contam. Part B 2018, 11, 83–91. [Google Scholar] [CrossRef]
- United States Food and Drug Administration. Pesticide Residue Monitoring Program Fiscal Year 2020 Pesticide Report; FDA: Washington, DC, USA, 2022. [Google Scholar]
- United States Department of Agriculture. Pesticide Data Program Annual Summary, Calendar Year 2021; USDA: Washington, DC, USA, 2022. [Google Scholar]
- Lesseur, C.; Pirrotte, P.; Pathak, K.V.; Manservisi, F.; Mandrioli, D.; Belpoggi, F.; Panzacchi, S.; Li, Q.; Barrett, E.S.; Nguyen, R.H.N.; et al. Maternal Urinary Levels of Glyphosate during Pregnancy and Anogenital Distance in Newborns in a US Multicenter Pregnancy Cohort. Environ. Pollut. Barking Essex 1987 2021, 280, 117002. [Google Scholar] [CrossRef]
- Lucia, R.M.; Liao, X.; Huang, W.-L.; Forman, D.; Kim, A.; Ziogas, A.; Norden-Krichmar, T.M.; Goodman, D.; Alvarez, A.; Masunaka, I.; et al. Urinary Glyphosate and AMPA Levels in a Cross-Sectional Study of Postmenopausal Women: Associations with Organic Eating Behavior and Dietary Intake. Int. J. Hyg. Environ. Health 2023, 252, 114211. [Google Scholar] [CrossRef]
- Lucia, R.M.; Huang, W.-L.; Pathak, K.V.; McGilvrey, M.; David-Dirgo, V.; Alvarez, A.; Goodman, D.; Masunaka, I.; Odegaard, A.O.; Ziogas, A.; et al. Association of Glyphosate Exposure with Blood DNA Methylation in a Cross-Sectional Study of Postmenopausal Women. Environ. Health Perspect. 2022, 130, 47001. [Google Scholar] [CrossRef]
- Mills, P.J.; Kania-Korwel, I.; Fagan, J.; McEvoy, L.K.; Laughlin, G.A.; Barrett-Connor, E. Excretion of the Herbicide Glyphosate in Older Adults between 1993 and 2016. JAMA 2017, 318, 1610–1611. [Google Scholar] [CrossRef]
- Parvez, S.; Gerona, R.R.; Proctor, C.; Friesen, M.; Ashby, J.L.; Reiter, J.L.; Lui, Z.; Winchester, P.D. Glyphosate Exposure in Pregnancy and Shortened Gestational Length: A Prospective Indiana Birth Cohort Study. Environ. Health Glob. Access Sci. Source 2018, 17, 23. [Google Scholar] [CrossRef]
- Schütze, A.; Morales-Agudelo, P.; Vidal, M.; Calafat, A.M.; Ospina, M. Quantification of Glyphosate and Other Organophosphorus Compounds in Human Urine via Ion Chromatography Isotope Dilution Tandem Mass Spectrometry. Chemosphere 2021, 274, 129427. [Google Scholar] [CrossRef]
- Zhang, L.; Rana, I.; Shaffer, R.M.; Taioli, E.; Sheppard, L. Exposure to Glyphosate-Based Herbicides and Risk for Non-Hodgkin Lymphoma: A Meta-Analysis and Supporting Evidence. Mutat. Res. 2019, 781, 186–206. [Google Scholar] [CrossRef]
- Boffetta, P.; Ciocan, C.; Zunarelli, C.; Pira, E. Exposure to Glyphosate and Risk of Non-Hodgkin Lymphoma: An Updated Meta-Analysis. Med. Lav. 2021, 112, 194–199. [Google Scholar] [CrossRef] [PubMed]
- Mink, P.J.; Mandel, J.S.; Sceurman, B.K.; Lundin, J.I. Epidemiologic Studies of Glyphosate and Cancer: A Review. Regul. Toxicol. Pharmacol. 2012, 63, 440–452. [Google Scholar] [CrossRef]
- Davoren, M.J.; Schiestl, R.H. Glyphosate-Based Herbicides and Cancer Risk: A Post-IARC Decision Review of Potential Mechanisms, Policy and Avenues of Research. Carcinogenesis 2018, 39, 1207–1215. [Google Scholar] [CrossRef]
- Marino, M.; Mele, E.; Viggiano, A.; Nori, S.L.; Meccariello, R.; Santoro, A. Pleiotropic Outcomes of Glyphosate Exposure: From Organ Damage to Effects on Inflammation, Cancer, Reproduction and Development. Int. J. Mol. Sci. 2021, 22, 12606. [Google Scholar] [CrossRef]
- International Agency for Research on Cancer. IARC Monographs on the Evaluation of Carcinogenic Risks to Humans; IARC: Lyon, France, 2017; pp. 321–399. [Google Scholar]
- United States Environmental Protection Agency. Revised Glyphosate Issue Paper: Evaluation of Carcinogenic Potential; EPA: Washington, DC, USA, 2017. [Google Scholar]
- United States Environmental Protection Agency Glyphosate. Available online: https://www.epa.gov/ingredients-used-pesticide-products/glyphosate (accessed on 23 April 2023).
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef]
- Lichtenstein, P.; Holm, N.V.; Verkasalo, P.K.; Iliadou, A.; Kaprio, J.; Koskenvuo, M.; Pukkala, E.; Skytthe, A.; Hemminki, K. Environmental and Heritable Factors in the Causation of Cancer: Analyses of Cohorts of Twins from Sweden, Denmark, and Finland. N. Engl. J. Med. 2000, 343, 78–85. [Google Scholar] [CrossRef]
- Foulkes, W.D. Inherited Susceptibility to Common Cancers. N. Engl. J. Med. 2008, 359, 2143. [Google Scholar] [CrossRef]
- Nelson, H.D.; Zakher, B.; Cantor, A.; Fu, R.; Griffin, J.; O’Meara, E.S.; Buist, D.S.M.; Kerlikowske, K.; van Ravesteyn, N.T.; Trentham-Dietz, A.; et al. Risk Factors for Breast Cancer for Women Age 40 to 49: A Systematic Review and Meta-Analysis. Ann. Intern. Med. 2012, 156, 635. [Google Scholar] [CrossRef]
- Singletary, S.E. Rating the Risk Factors for Breast Cancer. Ann. Surg. 2003, 237, 474–482. [Google Scholar] [CrossRef]
- Franke, A.A.; Li, X.; Shvetsov, Y.B.; Lai, J.F. Pilot Study on the Urinary Excretion of the Glyphosate Metabolite Aminomethylphosphonic Acid and Breast Cancer Risk: The Multiethnic Cohort Study. Environ. Pollut. Barking Essex 1987 2021, 277, 116848. [Google Scholar] [CrossRef]
- Varshavsky, J.; Smith, A.; Wang, A.; Hom, E.; Izano, M.; Huang, H.; Padula, A.; Woodruff, T.J. Heightened Susceptibility: A Review of How Pregnancy and Chemical Exposures Influence Maternal Health. Reprod. Toxicol. Elmsford N 2020, 92, 14–56. [Google Scholar] [CrossRef]
- Eaton, J.L.; Cathey, A.L.; Fernandez, J.A.; Watkins, D.J.; Silver, M.K.; Milne, G.L.; Velez-Vega, C.; Rosario, Z.; Cordero, J.; Alshawabkeh, A.; et al. The Association between Urinary Glyphosate and Aminomethyl Phosphonic Acid with Biomarkers of Oxidative Stress among Pregnant Women in the PROTECT Birth Cohort Study. Ecotoxicol. Environ. Saf. 2022, 233, 113300. [Google Scholar] [CrossRef]
- Valko, M.; Rhodes, C.J.; Moncol, J.; Izakovic, M.; Mazur, M. Free Radicals, Metals and Antioxidants in Oxidative Stress-Induced Cancer. Chem. Biol. Interact. 2006, 160, 1–40. [Google Scholar] [CrossRef]
- Reuter, S.; Gupta, S.C.; Chaturvedi, M.M.; Aggarwal, B.B. Oxidative Stress, Inflammation, and Cancer: How Are They Linked? Free Radic. Biol. Med. 2010, 49, 1603–1616. [Google Scholar] [CrossRef]
- Cairns, R.A.; Harris, I.S.; Mak, T.W. Regulation of Cancer Cell Metabolism. Nat. Rev. Cancer 2011, 11, 85–95. [Google Scholar] [CrossRef]
- Page, M.J.; McKenzie, J.E.; Bossuyt, P.M.; Boutron, I.; Hoffmann, T.C.; Mulrow, C.D.; Shamseer, L.; Tetzlaff, J.M.; Akl, E.A.; Brennan, S.E.; et al. The PRISMA 2020 Statement: An Updated Guideline for Reporting Systematic Reviews. BMJ 2021, 372, n71. [Google Scholar] [CrossRef]
- Salazar-Martinez, E.; Romano-Riquer, P.; Yanez-Marquez, E.; Longnecker, M.P.; Hernandez-Avila, M. Anogenital Distance in Human Male and Female Newborns: A Descriptive, Cross-Sectional Study. Environ. Health 2004, 3, 8. [Google Scholar] [CrossRef]
- Swan, S.H.; Kristensen, D.M. Anogenital Distance: A Marker of Steroidal Endocrine Disruption. In Encyclopedia of Reproduction, 2nd ed.; Skinner, M.K., Ed.; Academic Press: Oxford, UK, 2018; pp. 588–593. ISBN 978-0-12-815145-7. [Google Scholar]
- Kresovich, J.K.; Xu, Z.; O’Brien, K.M.; Weinberg, C.R.; Sandler, D.P.; Taylor, J.A. Methylation-Based Biological Age and Breast Cancer Risk. J. Natl. Cancer Inst. 2019, 111, 1051–1058. [Google Scholar] [CrossRef]
- Ambatipudi, S.; Horvath, S.; Perrier, F.; Cuenin, C.; Hernandez-Vargas, H.; Le Calvez-Kelm, F.; Durand, G.; Byrnes, G.; Ferrari, P.; Bouaoun, L.; et al. DNA Methylome Analysis Identifies Accelerated Epigenetic Ageing Associated with Postmenopausal Breast Cancer Susceptibility. Eur. J. Cancer 2017, 75, 299–307. [Google Scholar] [CrossRef]
- Garcia-Martinez, L.; Zhang, Y.; Nakata, Y.; Chan, H.L.; Morey, L. Epigenetic Mechanisms in Breast Cancer Therapy and Resistance. Nat. Commun. 2021, 12, 1786. [Google Scholar] [CrossRef] [PubMed]
- Stur, E.; Aristizabal-Pachon, A.F.; Peronni, K.C.; Agostini, L.P.; Waigel, S.; Chariker, J.; Miller, D.M.; Thomas, S.D.; Rezzoug, F.; Detogni, R.S.; et al. Glyphosate-Based Herbicides at Low Doses Affect Canonical Pathways in Estrogen Positive and Negative Breast Cancer Cell Lines. PLoS ONE 2019, 14, e0219610. [Google Scholar] [CrossRef]
- Coppola, L.; Tait, S.; Fabbrizi, E.; Perugini, M.; La Rocca, C. Comparison of the Toxicological Effects of Pesticides in Non-Tumorigenic MCF-12A and Tumorigenic MCF-7 Human Breast Cells. Int. J. Environ. Res. Public Health 2022, 19, 4453. [Google Scholar] [CrossRef]
- Mesnage, R.; Phedonos, A.; Biserni, M.; Arno, M.; Balu, S.; Corton, J.C.; Ugarte, R.; Antoniou, M.N. Evaluation of Estrogen Receptor Alpha Activation by Glyphosate-Based Herbicide Constituents. Food Chem. Toxicol. 2017, 108, 30–42. [Google Scholar] [CrossRef]
- Thongprakaisang, S.; Thiantanawat, A.; Rangkadilok, N.; Suriyo, T.; Satayavivad, J. Glyphosate Induces Human Breast Cancer Cells Growth via Estrogen Receptors. Food Chem. Toxicol. 2013, 59, 129–136. [Google Scholar] [CrossRef] [PubMed]
- Lin, N.; Garry, V.F. In Vitro Studies of Cellular and Molecular Developmental Toxicity of Adjuvants, Herbicides, and Fungicides Commonly Used in Red River Valley, Minnesota. J. Toxicol. Environ. Health A 2000, 60, 423–439. [Google Scholar] [CrossRef] [PubMed]
- De Almeida, L.K.S.; Pletschke, B.I.; Frost, C.L. Moderate Levels of Glyphosate and Its Formulations Vary in Their Cytotoxicity and Genotoxicity in a Whole Blood Model and in Human Cell Lines with Different Estrogen Receptor Status. 3 Biotech 2018, 8, 438. [Google Scholar] [CrossRef]
- Zhang, C.; Schilirò, T.; Gea, M.; Bianchi, S.; Spinello, A.; Magistrato, A.; Gilardi, G.; Di Nardo, G. Molecular Basis for Endocrine Disruption by Pesticides Targeting Aromatase and Estrogen Receptor. Int. J. Environ. Res. Public Health 2020, 17, 5664. [Google Scholar] [CrossRef]
- Gasnier, C.; Dumont, C.; Benachour, N.; Clair, E.; Chagnon, M.-C.; Séralini, G.-E. Glyphosate-Based Herbicides Are Toxic and Endocrine Disruptors in Human Cell Lines. Toxicology 2009, 262, 184–191. [Google Scholar] [CrossRef]
- Hokanson, R.; Fudge, R.; Chowdhary, R.; Busbee, D. Alteration of Estrogen-Regulated Gene Expression in Human Cells Induced by the Agricultural and Horticultural Herbicide Glyphosate. Hum. Exp. Toxicol. 2007, 26, 747–752. [Google Scholar] [CrossRef]
- McKeown, S.R. Defining Normoxia, Physoxia and Hypoxia in Tumours—Implications for Treatment Response. Br. J. Radiol. 2014, 87, 20130676. [Google Scholar] [CrossRef]
- Infantino, V.; Santarsiero, A.; Convertini, P.; Todisco, S.; Iacobazzi, V. Cancer Cell Metabolism in Hypoxia: Role of HIF-1 as Key Regulator and Therapeutic Target. Int. J. Mol. Sci. 2021, 22, 5703. [Google Scholar] [CrossRef]
- Yang, Y.; Li, J.; Lei, W.; Wang, H.; Ni, Y.; Liu, Y.; Yan, H.; Tian, Y.; Wang, Z.; Yang, Z.; et al. CXCL12-CXCR4/CXCR7 Axis in Cancer: From Mechanisms to Clinical Applications. Int. J. Biol. Sci. 2023, 19, 3341–3359. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Guo, H.; Yu, H.; Chen, Y.; Xu, H.; Zhao, G. The Role of the Transcription Factor EGR1 in Cancer. Front. Oncol. 2021, 11, 642547. [Google Scholar] [CrossRef]
- Antoniou, M.N.; Nicolas, A.; Mesnage, R.; Biserni, M.; Rao, F.V.; Martin, C.V. Glyphosate Does Not Substitute for Glycine in Proteins of Actively Dividing Mammalian Cells. BMC Res. Notes 2019, 12, 494. [Google Scholar] [CrossRef] [PubMed]
- Mueller, S.H.; Lai, A.G.; Valkovskaya, M.; Michailidou, K.; Bolla, M.K.; Wang, Q.; Dennis, J.; Lush, M.; Abu-Ful, Z.; Ahearn, T.U.; et al. Aggregation Tests Identify New Gene Associations with Breast Cancer in Populations with Diverse Ancestry. Genome Med. 2023, 15, 7. [Google Scholar] [CrossRef] [PubMed]
- Miedl, H.; Oswald, D.; Haslinger, I.; Gstoettner, M.; Wenzl, R.; Proestling, K.; Schneeberger, C.; Yotova, I.; Schreiber, M. Association of the Estrogen Receptor 1 Polymorphisms Rs2046210 and Rs9383590 with the Risk, Age at Onset and Prognosis of Breast Cancer. Cells 2023, 12, 515. [Google Scholar] [CrossRef]
- Baker, M.E.; Lathe, R. The Promiscuous Estrogen Receptor: Evolution of Physiological Estrogens and Response to Phytochemicals and Endocrine Disruptors. J. Steroid Biochem. Mol. Biol. 2018, 184, 29–37. [Google Scholar] [CrossRef]
- Benachour, N.; Sipahutar, H.; Moslemi, S.; Gasnier, C.; Travert, C.; Séralini, G.E. Time- and Dose-Dependent Effects of Roundup on Human Embryonic and Placental Cells. Arch. Environ. Contam. Toxicol. 2007, 53, 126–133. [Google Scholar] [CrossRef] [PubMed]
- Mesnage, R.; Bernay, B.; Séralini, G.-E. Ethoxylated Adjuvants of Glyphosate-Based Herbicides Are Active Principles of Human Cell Toxicity. Toxicology 2013, 313, 122–128. [Google Scholar] [CrossRef] [PubMed]
- Richard, S.; Moslemi, S.; Sipahutar, H.; Benachour, N.; Seralini, G.-E. Differential Effects of Glyphosate and Roundup on Human Placental Cells and Aromatase. Environ. Health Perspect. 2005, 113, 716–720. [Google Scholar] [CrossRef] [PubMed]
- Defarge, N.; Takács, E.; Lozano, V.L.; Mesnage, R.; Spiroux de Vendômois, J.; Séralini, G.-E.; Székács, A. Co-Formulants in Glyphosate-Based Herbicides Disrupt Aromatase Activity in Human Cells below Toxic Levels. Int. J. Environ. Res. Public Health 2016, 13, 264. [Google Scholar] [CrossRef] [PubMed]
- Truzzi, F.; Mandrioli, D.; Gnudi, F.; Scheepers, P.T.J.; Silbergeld, E.K.; Belpoggi, F.; Dinelli, G. Comparative Evaluation of the Cytotoxicity of Glyphosate-Based Herbicides and Glycine in L929 and Caco2 Cells. Front. Public Health 2021, 9, 643898. [Google Scholar] [CrossRef] [PubMed]
- Defarge, N.; Spiroux de Vendômois, J.; Séralini, G.E. Toxicity of Formulants and Heavy Metals in Glyphosate-Based Herbicides and Other Pesticides. Toxicol. Rep. 2018, 5, 156–163. [Google Scholar] [CrossRef] [PubMed]
- Sritana, N.; Suriyo, T.; Kanitwithayanun, J.; Songvasin, B.H.; Thiantanawat, A.; Satayavivad, J. Glyphosate Induces Growth of Estrogen Receptor Alpha Positive Cholangiocarcinoma Cells via Non-Genomic Estrogen Receptor/ERK1/2 Signaling Pathway. Food Chem. Toxicol. 2018, 118, 595–607. [Google Scholar] [CrossRef] [PubMed]
- Clair, É.; Mesnage, R.; Travert, C.; Séralini, G.-É. A Glyphosate-Based Herbicide Induces Necrosis and Apoptosis in Mature Rat Testicular Cells in Vitro, and Testosterone Decrease at Lower Levels. Toxicol. In Vitro 2012, 26, 269–279. [Google Scholar] [CrossRef] [PubMed]
- Coperchini, F.; Greco, A.; Croce, L.; Denegri, M.; Magri, F.; Rotondi, M.; Chiovato, L. In Vitro Study of Glyphosate Effects on Thyroid Cells. Environ. Pollut. 2023, 317, 120801. [Google Scholar] [CrossRef]
- Mesnage, R.; Ibragim, M.; Mandrioli, D.; Falcioni, L.; Tibaldi, E.; Belpoggi, F.; Brandsma, I.; Bourne, E.; Savage, E.; Mein, C.A.; et al. Comparative Toxicogenomics of Glyphosate and Roundup Herbicides by Mammalian Stem Cell-Based Genotoxicity Assays and Molecular Profiling in Sprague-Dawley Rats. Toxicol. Sci. 2021, 186, 83–101. [Google Scholar] [CrossRef]
- Vanlaeys, A.; Dubuisson, F.; Seralini, G.-E.; Travert, C. Formulants of Glyphosate-Based Herbicides Have More Deleterious Impact than Glyphosate on TM4 Sertoli Cells. Toxicol. Vitro Int. J. Publ. Assoc. BIBRA 2018, 52, 14–22. [Google Scholar] [CrossRef] [PubMed]
- Gillezeau, C.; van Gerwen, M.; Shaffer, R.M.; Rana, I.; Zhang, L.; Sheppard, L.; Taioli, E. The Evidence of Human Exposure to Glyphosate: A Review. Environ. Health 2019, 18, 2. [Google Scholar] [CrossRef]
- Faniband, M.H.; Norén, E.; Littorin, M.; Lindh, C.H. Human Experimental Exposure to Glyphosate and Biomonitoring of Young Swedish Adults. Int. J. Hyg. Environ. Health 2021, 231, 113657. [Google Scholar] [CrossRef]
- Connolly, A.; Jones, K.; Basinas, I.; Galea, K.S.; Kenny, L.; McGowan, P.; Coggins, M.A. Exploring the Half-Life of Glyphosate in Human Urine Samples. Int. J. Hyg. Environ. Health 2019, 222, 205–210. [Google Scholar] [CrossRef]
- Nakamura, J.; Lu, Q.; Aberdeen, G.; Albrecht, E.; Brodie, A. The Effect of Estrogen on Aromatase and Vascular Endothelial Growth Factor Messenger Ribonucleic Acid in the Normal Nonhuman Primate Mammary Gland. J. Clin. Endocrinol. Metab. 1999, 84, 1432–1437. [Google Scholar] [CrossRef]
- Romaniuk, A.; Lyndin, M.; Sikora, V.; Lyndina, Y.; Romaniuk, S.; Sikora, K. Heavy Metals Effect on Breast Cancer Progression. J. Occup. Med. Toxicol. Lond. Engl. 2017, 12, 32. [Google Scholar] [CrossRef] [PubMed]
- Cadet, J.; Douki, T.; Gasparutto, D.; Ravanat, J.-L. Oxidative Damage to DNA: Formation, Measurement and Biochemical Features. Mutat. Res. Mol. Mech. Mutagen. 2003, 531, 5–23. [Google Scholar] [CrossRef]
- Kwiatkowska, M.; Reszka, E.; Woźniak, K.; Jabłońska, E.; Michałowicz, J.; Bukowska, B. DNA Damage and Methylation Induced by Glyphosate in Human Peripheral Blood Mononuclear Cells (in Vitro Study). Food Chem. Toxicol. 2017, 105, 93–98. [Google Scholar] [CrossRef] [PubMed]
- Suárez-Larios, K.; Salazar-Martínez, A.-M.; Montero-Montoya, R. Screening of Pesticides with the Potential of Inducing DSB and Successive Recombinational Repair. J. Toxicol. 2017, 2017, 3574840. [Google Scholar] [CrossRef]
- Woźniak, E.; Sicińska, P.; Michałowicz, J.; Woźniak, K.; Reszka, E.; Huras, B.; Zakrzewski, J.; Bukowska, B. The Mechanism of DNA Damage Induced by Roundup 360 PLUS, Glyphosate and AMPA in Human Peripheral Blood Mononuclear Cells—Genotoxic Risk Assessement. Food Chem. Toxicol. 2018, 120, 510–522. [Google Scholar] [CrossRef]
- Peillex, C.; Pelletier, M. The Impact and Toxicity of Glyphosate and Glyphosate-Based Herbicides on Health and Immunity. J. Immunotoxicol. 2020, 17, 163–174. [Google Scholar] [CrossRef]
- Sender, R.; Fuchs, S.; Milo, R. Revised Estimates for the Number of Human and Bacteria Cells in the Body. PLoS Biol. 2016, 14, e1002533. [Google Scholar] [CrossRef]
- Lynch, S.V.; Pedersen, O. The Human Intestinal Microbiome in Health and Disease. N. Engl. J. Med. 2016, 375, 2369–2379. [Google Scholar] [CrossRef] [PubMed]
- Samsel, A.; Seneff, S. Glyphosate, Pathways to Modern Diseases III: Manganese, Neurological Diseases, and Associated Pathologies. Surg. Neurol. Int. 2015, 6, 45. [Google Scholar] [CrossRef]
- Puigbò, P.; Leino, L.I.; Rainio, M.J.; Saikkonen, K.; Saloniemi, I.; Helander, M. Does Glyphosate Affect the Human Microbiota? Life 2022, 12, 707. [Google Scholar] [CrossRef]
- Rossetti, M.F.; Canesini, G.; Lorenz, V.; Milesi, M.M.; Varayoud, J.; Ramos, J.G. Epigenetic Changes Associated with Exposure to Glyphosate-Based Herbicides in Mammals. Front. Endocrinol. 2021, 12, 671991. [Google Scholar] [CrossRef] [PubMed]
- Bukowska, B.; Woźniak, E.; Sicińska, P.; Mokra, K.; Michałowicz, J. Glyphosate Disturbs Various Epigenetic Processes in Vitro and in Vivo—A Mini Review. Sci. Total Environ. 2022, 851, 158259. [Google Scholar] [CrossRef]
- Muñoz, J.P.; Bleak, T.C.; Calaf, G.M. Glyphosate and the Key Characteristics of an Endocrine Disruptor: A Review. Chemosphere 2021, 270, 128619. [Google Scholar] [CrossRef] [PubMed]
- Ingaramo, P.; Alarcón, R.; Muñoz-de-Toro, M.; Luque, E.H. Are Glyphosate and Glyphosate-Based Herbicides Endocrine Disruptors That Alter Female Fertility? Mol. Cell. Endocrinol. 2020, 518, 110934. [Google Scholar] [CrossRef]
- de Araújo-Ramos, A.T.; Passoni, M.T.; Romano, M.A.; Romano, R.M.; Martino-Andrade, A.J. Controversies on Endocrine and Reproductive Effects of Glyphosate and Glyphosate-Based Herbicides: A Mini-Review. Front. Endocrinol. 2021, 12, 627210. [Google Scholar] [CrossRef]
- Kaboli Kafshgiri, S.; Farkhondeh, T.; Miri-Moghaddam, E. Glyphosate Effects on the Female Reproductive Systems: A Systematic Review. Rev. Environ. Health 2022, 37, 487–500. [Google Scholar] [CrossRef] [PubMed]
- Serra, L.; Estienne, A.; Vasseur, C.; Froment, P.; Dupont, J. Review: Mechanisms of Glyphosate and Glyphosate-Based Herbicides Action in Female and Male Fertility in Humans and Animal Models. Cells 2021, 10, 3079. [Google Scholar] [CrossRef]
- Milesi, M.M.; Lorenz, V.; Durando, M.; Rossetti, M.F.; Varayoud, J. Glyphosate Herbicide: Reproductive Outcomes and Multigenerational Effects. Front. Endocrinol. 2021, 12, 672532. [Google Scholar] [CrossRef]
- Guyton, K.Z.; Loomis, D.; Grosse, Y.; El Ghissassi, F.; Benbrahim-Tallaa, L.; Guha, N.; Scoccianti, C.; Mattock, H.; Straif, K.; International Agency for Research on Cancer Monograph Working Group. Carcinogenicity of Tetrachlorvinphos, Parathion, Malathion, Diazinon, and Glyphosate. Lancet Oncol. 2015, 16, 490–491. [Google Scholar] [CrossRef]
- Eriksson, M.; Hardell, L.; Carlberg, M.; Akerman, M. Pesticide Exposure as Risk Factor for Non-Hodgkin Lymphoma Including Histopathological Subgroup Analysis. Int. J. Cancer 2008, 123, 1657–1663. [Google Scholar] [CrossRef]
- Karunanayake, C.P.; Spinelli, J.J.; McLaughlin, J.R.; Dosman, J.A.; Pahwa, P.; McDuffie, H.H. Hodgkin Lymphoma and Pesticides Exposure in Men: A Canadian Case-Control Study. J. Agromedicine 2012, 17, 30–39. [Google Scholar] [CrossRef] [PubMed]
- De Roos, A.J.; Blair, A.; Rusiecki, J.A.; Hoppin, J.A.; Svec, M.; Dosemeci, M.; Sandler, D.P.; Alavanja, M.C. Cancer Incidence among Glyphosate-Exposed Pesticide Applicators in the Agricultural Health Study. Environ. Health Perspect. 2005, 113, 49–54. [Google Scholar] [CrossRef]
- Andreotti, G.; Koutros, S.; Hofmann, J.N.; Sandler, D.P.; Lubin, J.H.; Lynch, C.F.; Lerro, C.C.; De Roos, A.J.; Parks, C.G.; Alavanja, M.C.; et al. Glyphosate Use and Cancer Incidence in the Agricultural Health Study. JNCI J. Natl. Cancer Inst. 2017, 110, 509–516. [Google Scholar] [CrossRef]
- Engel, L.S.; Werder, E.; Satagopan, J.; Blair, A.; Hoppin, J.A.; Koutros, S.; Lerro, C.C.; Sandler, D.P.; Alavanja, M.C.; Beane Freeman, L.E. Insecticide Use and Breast Cancer Risk among Farmers’ Wives in the Agricultural Health Study. Environ. Health Perspect. 2017, 125, 097002. [Google Scholar] [CrossRef] [PubMed]
- Hill, K.G.; Woodward, D.; Woelfel, T.; Hawkins, J.D.; Green, S. Planning for Long-Term Follow-Up: Strategies Learned from Longitudinal Studies. Prev. Sci. Off. J. Soc. Prev. Res. 2016, 17, 806–818. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schluter, H.M.; Bariami, H.; Park, H.L. Potential Role of Glyphosate, Glyphosate-Based Herbicides, and AMPA in Breast Cancer Development: A Review of Human and Human Cell-Based Studies. Int. J. Environ. Res. Public Health 2024, 21, 1087. https://doi.org/10.3390/ijerph21081087
Schluter HM, Bariami H, Park HL. Potential Role of Glyphosate, Glyphosate-Based Herbicides, and AMPA in Breast Cancer Development: A Review of Human and Human Cell-Based Studies. International Journal of Environmental Research and Public Health. 2024; 21(8):1087. https://doi.org/10.3390/ijerph21081087
Chicago/Turabian StyleSchluter, Hannah M., Hajar Bariami, and Hannah Lui Park. 2024. "Potential Role of Glyphosate, Glyphosate-Based Herbicides, and AMPA in Breast Cancer Development: A Review of Human and Human Cell-Based Studies" International Journal of Environmental Research and Public Health 21, no. 8: 1087. https://doi.org/10.3390/ijerph21081087