Evaluation of the Removal of Indicator Bacteria from Domestic Sludge Processed by Autothermal Thermophilic Aerobic Digestion (ATAD)

Abstract
:1. Introduction
2. Experimental Section
2.1. ATAD Sludge Source and Sampling
2.2. Microbiological Analysis of ATAD Sludge Quality
2.2.1. Enumeration and Detection of the Indicator Organisms
2.2.2. Biochemical Identification of Microorganisms
2.2.3. Molecular Identification of ATAD Organisms via Intergenic Spacer Region Analysis
2.2.4. DNA Extraction via the CTAB Extraction Method
2.2.5. RISA-PCR
2.2.6. PCR Amplification of the V6-V8 Region of 16S rDNA Genes for Phylogenetic Identification
2.2.7. Cloning of the 16S rDNA Amplicons
2.3. Assessment of Sludge Bulk Water for Nuclease Activity with Potential to Degrade Cell-Free DNA
2.4. SXT/R391-like ICE Mobile Element Detection
3. Results
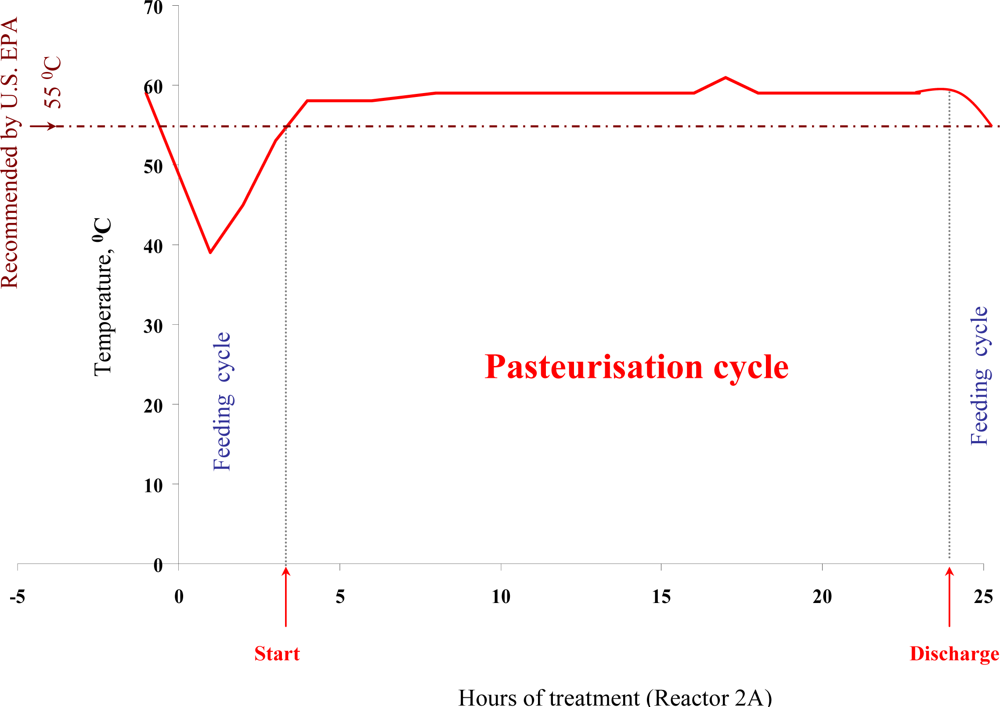
3.1. Alteration of Process Parameters during ATAD Treatment
3.2. Microbiological Analysis of the Indicator Bacteria and Fate during the Sludge Processing
3.2.1. Non-Presumptive Bacterial Growth on the Selective Plates
3.2.2. Removal of Pathogens during ATAD Process
3.2.3. Seasonal Detection of Indicator Bacteria
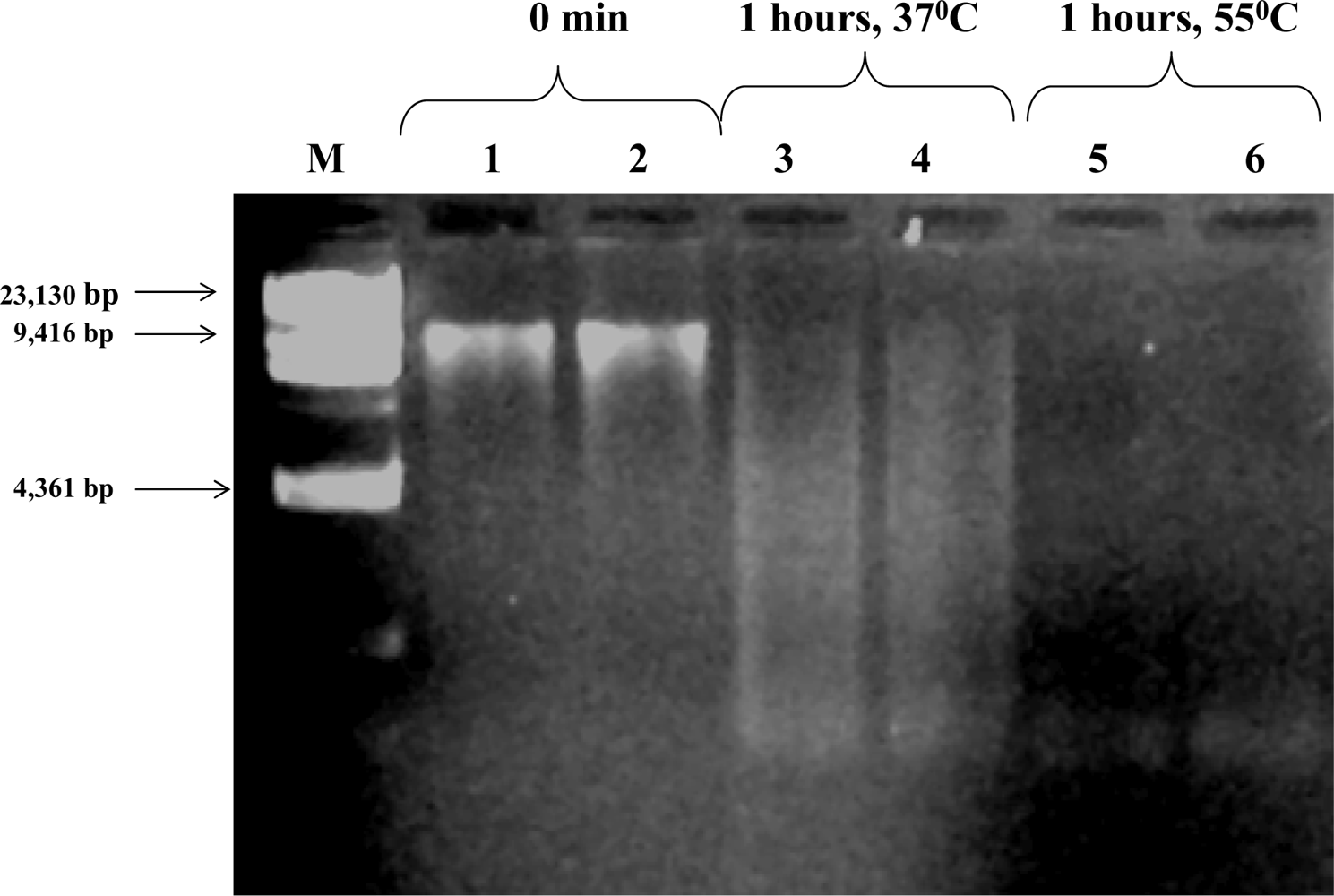
3.4. Exogenous DNA Degradation by the Sludge
3.5. Fate of the Mobile Elements during the ATAD Treatment
4. Discussion
5. Conclusions
Acknowledgments
References and Notes
- Feachem, RG; Bradley, DJ; Garelick, H; Mara, DD. Sanitation and Disease: Health Aspects of Excreta and Wastewater Management; John Wiley and Sons: Chichester, UK, 1983. [Google Scholar]
- Miguéns, JL; Mendes, JF. Travel and tourism: into a complex network. Physica A 2008, 387, 1. [Google Scholar]
- Tchobanogluous, G; Burton, FL. Wastewater Engineering: Treatment, Disposal, and Reuse; Irwin/McGraw-Hill: Boston, MA, USA, 1991. [Google Scholar]
- Kelly, HG; Melcer, H; Mavinic, DS. Autothermal thermophilic aerobic digestion of municipal sludges: A one-year, full-scale demonstration project. Water Environ. Res 1993, 657, 849–861. [Google Scholar]
- Autothermal Thermophilic Aerobic Digestion of Municipal Wastewater Sludge; Report EPA/625/10-90/007; US Environmental Protection Agency: Washington, DC, USA, 1990.
- Kelly, HG. Emerging processes in biosolids treatment. J. Environ. Eng. Sci 2006, 5, 175–186. [Google Scholar]
- LaPara, TM; Alleman, JE. Thermophilic aerobic biological wastewater treatment. Water Res 1999, 33, 895–908. [Google Scholar]
- Piterina, AV; MacCusland, C; Bartlett, J; Pembroke, JT. Microbial ecology of auto-thermal aerobic digestion (ATAD): Diversity, dynamics and activity of bacterial communities involved in treatment of a municipal wastewater’. In Recent Advances in Applied Microbiology; Understanding and Exploiting Microbes and Their Interactions Biological, Physical, Chemical and Engineering Aspects; Formatex: Badajos, Spain, 2006; pp. 210–221. [Google Scholar]
- Layden, NM; Mavinic, DS; Kelly, HG; Moles, R; Bartlett, J. Autothermal thermophilic aerobic digestion (ATAD)—Part I: Review of origins, design, and process operation. J. Environ. Eng. Sci 2007, 6, 665–678. [Google Scholar]
- Layden, N; Kelly, H; Mavinic, D; Moles, R; Bartlett, J. Autothermal thermophilic aerobic digestion (ATAD)—Part II: Review of research and full-scale operating experiences. J. Environ. Eng. Sci 2007, 6, 679–690. [Google Scholar]
- LaPara, TM; Nakatsu, CH; Pantea, L; Alleman, JE. Phylogenetic analysis of bacterial communities in mesophilic and thermophilic bioreactors treating pharmaceutical wastewater. Appl. Environ. Microbiol 2000, 66, 3951–3959. [Google Scholar]
- EU Sludge Directive. Available online: http://ec.europa.eu/environment/waste/sludge/index.htm (accessed on 1 August 2010).
- Codes for Good Practise; Department of the Environment and Local Government & Department of Agriculture and Food and EPA: Dublin, Ireland, 1994. Available online: http://www.environ.ie/en/Publications/Environment/Water/FileDownLoad,17228,en.pdf (accessed on 1 August 2010).
- Ugwuanyi, JO; Harvey, LM; Mcneil, B. Effect of process temperature, pH and suspended solids content upon pasteurization of a model agricultural waste during thermophilic aerobic digestion. J. Appl. Microbiol 1999, 87, 387–395. [Google Scholar]
- Zabranska, J; Dohanyos, M; Jenicek, P; Ruzicikova, H; Vranova, A. Efficiency of autothermal thermophilic aerobic digestion and thermophilic anaerobic digestion of municipal wastewater sludge in removing Salmonella spp. and indicator bacteria. Water Sci. Technol 2003, 47, 151–156. [Google Scholar]
- Watanabe, H. Inactivation of pathogenic bacteria under mesophilic and thermophilic conditions. Water Sci. Tech 1997, 36, 25–32. [Google Scholar]
- Strauch, D. Pathogenic micro-organisms in sludge. Anaerobic digestion and disinfection methods to make sludge usable as a fertiliser. Eur. Water Manage 1998, 1, 12–26. [Google Scholar]
- Ayres, RM; Mara, DD. Analysis of Wastewater for Use in Agriculture: A Laboratory Manual of Parasitological and Bacteriological Techniques; WHO: Geneva, Switzerland, 1996. [Google Scholar]
- Standard Methods for the Examination of Water and Wastewater, 19th ed; American Public Health Association: Washington, DC, USA, 1995.
- Carrington, EG; Davis, RD; Hall, JE; Pike, EB; Smith, SR; Unwin, RJ. Review of the scientific evidence relating to the controls on agricultural use of sewage sludge. In Report DETR4415/3 [part1] and Report DETR 4454/4; WRC Publications: Medmenham, UK, 1998. [Google Scholar]
- Gerba, CP; Pepper, IL; Whitehead, LF. A risk assessment of emerging pathogens of concern in the land application of biosolids. Water Sci. Technol 2002, 46, 225–230. [Google Scholar]
- Sidhu, J; Gibbs, RA; Ho, GE; Unkovich, I. Selection of Salmonella typhimurium as an indicator for pathogen regrowth potential in composted biosolids. Lett. Appl. Microbiol 1999, 29, 303–307. [Google Scholar]
- Zaleski, KJ; Josephson, KL; Gerba, CP; Pepper, IL. Potential regrowth and recolonization of salmonellae and indicators in biosolids and biosolid-amended soil. Appl. Environ. Microbiol 2005, 71, 3701–8708. [Google Scholar]
- Russ, CF; Yanko, WA. Factors affecting salmonella’s repopulation in composted sludges. Appl. Environ. Microbiol 1981, 41, 597–602. [Google Scholar]
- Pflug, IJ; Holcomb, RG; Gomez, MM. Principles of the thermal destruction of microorganisms. In Disinfection, Sterilization and Preservation, 5th ed; Block, SS, Ed.; Lippincott, Williams and Wilkins: Philadelphia, PA, USA, 2001; pp. 79–129. [Google Scholar]
- Moats, WA; Dabbah, R; Edwards, VM. Interpretation of nonlogarithmic survivor curves of heated bacteria. J. Food Sci 1971, 36, 523–526. [Google Scholar]
- Doyle, ME; Mazzotta, AS. Review of studies on the thermal resistance of Salmonellae. J. Food Prot 2000, 63, 779–795. [Google Scholar]
- Chiruta, J; Davey, KR; Thomas, CJ. Thermal inactivation kinetics of three vegetative bacteria as influenced by combined temperature and pH in a liquid medium. Food Bioprod. Process 1997, 75, 174–180. [Google Scholar]
- Smith, MG. Survival of E. coli and Salmonella after chilling and freezing in liquid media. J. Food Sci 1995, 60, 509–512. [Google Scholar]
- Spinks, AT; Dunstan, RH; Harrison, T; Coombes, P; Kuczera, G. Thermal inactivation of water-borne pathogenic and indicator bacteria at sub-boiling temperatures. Water Res 2006, 40, 1326–1332. [Google Scholar]
- Sharma, S; Sachdeva, P; Virdi, JS. Emerging water-borne pathogens. Appl. Microbiol. Biotechnol 2003, 61, 424–428. [Google Scholar]
- Godfree, A; Farrell, J. Processes for managing pathogens. J. Environ. Qual 2005, 34, 105–113. [Google Scholar]
- Hay, JC. Pathogen destruction and biosolids compost. Biocycle 1996, 37, 67–77. [Google Scholar]
- United States Environmental Protection Agency; 40 CFR Part 503. Standards for the use or disposal of sewage sludge. Fed Regist 1994, 58, 9248–9415.
- Finkel, SE; Kolter, R. DNA as a nutrient: novel role for bacterial competence gene homologs. J. Bacteriol 2001, 183, 6288–6293. [Google Scholar]
- Nielsen, KM; Johnsen, PJ; Bensasson, D; Daffonchio, D. Release and persistence of extracellular DNA in the environment. Environ .Safety Res 2007, 61, 37–53. [Google Scholar]
- Chen, I; Dubnau, D. DNA uptake during bacterial transformation. Nat. Rev. Microbiol 2004, 3, 241–249. [Google Scholar]
- Dubnau, D. DNA uptake in bacteria. Annu. Rev. Microbiol 1999, 53, 217–244. [Google Scholar]
- Weiss, MS; Abele, U; Weckesser, J; Welte, W; Schiltz, E; Schulz, GE. Molecular architecture and electrostatic properties of a bacterial porin. Science 1991, 254, 1627–1630. [Google Scholar]
- Paul, JH; Jeffrey, WH; Deflaun, MF. Dynamics of extracellular DNA in the marine environment. Appl. Environ. Microbiol 1987, 53, 170–179. [Google Scholar]
- Burns, R. Enzymes in the Environment: Activity, Ecology and Applications; CRS Press, Taylor and Francis Group: New York, NY, USA, 2002. [Google Scholar]
- Aardema, BW; Lorenz, MG; Krumbein, WE. Protection of sediment-adsorbed transforming DNA against enzymatic inactivation. Appl. Environ. Microbiol 1983, 46, 417–420. [Google Scholar]
- Buchanan, JT; Simpson, AJ; Aziz, RK; Liu, GY; Kristian, SA; Kotb, M; Feramisco, J; Nizet, V. DNase expression allows the pathogen group a Streptococcus to escape killing in neutrophil extracellular traps. Curr. Biol 2006, 16, 396–400. [Google Scholar]
- Citak, S; Varlik, O; Gundogan, N. Slime production and DNase activity of Spaphylococci isolated from raw milk. J. Food Saf 2007, 23, 281–288. [Google Scholar]
- Kneitel, JM; Chase, JM. Trade-offs in community ecology: linking spatial scales and species coexistence. Ecol. Lett 2004, 7, 69–80. [Google Scholar]
- Ruiz, TR; Andrews, TR; Smith, GB. Identification and characterization of nuclease activities in anaerobic environmental samples. Can. J. Microbiol 2000, 46, 736–740. [Google Scholar]
- Ni, H. Bacteria learn antibiotic resistance in the sludge. Drug discov.Today 2003, 8, 10–11. [Google Scholar]
- Coughter, JP; Stewart, GJ. Genetic exchange in the environment. Antonie Leeuwenhoek 1989, 55, 15–22. [Google Scholar]
- Redfield, RJ. Genes for breakfast: The have-your-cake-and-eat-it-too of bacterial transformation. J. Hered 1993, 84, 400–404. [Google Scholar]
- White, PA; McIver, CJ; Rawlinson, WD. Integrons and gene cassettes in the Enterobacteriaceae. Antimicrob. Agents Chemother 2001, 45, 2658–2661. [Google Scholar]
- Moura, A; Henriques, I; Ribeiro, R; Correia, A. Prevalence and characterization of integrons from bacteria isolated from a slaughterhouse wastewater treatment plant. J. Antimicrob. Chemother 2007, 60, 1243–1250. [Google Scholar]
- da Costa, PM; Vaz-Pires, P; Bernardo, F. Antimicrobial resistance in Enterococcus spp. isolates in inflow, effluent and sludge from municipal sewage waste treatment plants. Water Res 2006, 40, 1735–1740. [Google Scholar]
- Schwartz, T; Kohnen, W; Janses, B. Detection of antibiotic resistant bacteria and their resistance genes in wastewater, surface water, and drinking water biofilms. FEMS Microbiol. Ecol 2003, 43, 325–335. [Google Scholar]
- Tennstedt, T; Szczepanowski, R; Braun, S. Occurrence of integron-associated resistance gene cassettes located on antibiotic resistance plasmids isolated from a wastewater treatment plant. FEMS Microbiol. Ecol 2003, 45, 239–252. [Google Scholar]
- McPherson, P; Gealt, MA. Isolation of indigenous wastewater bacterial strains capable of mobilizing plasmid pBR325. Appl. Environ. Microbiol 1986, 51, 904–909. [Google Scholar]
- Lorenz, MG; Wackernagel, W. Bacterial gene transfer by natural genetic transformation in the environment. Microbiol. Rev 1994, 58, 563–602. [Google Scholar]
- Levy, SB. The challenge of antibiotic resistance. Sci. Am 1998, 278, 32–39. [Google Scholar]
- Lindberg, RH; Björklund, K; Rendahl, P. Environmental risk assessment of antibiotics in the Swedish environment with emphasis on sewage treatment plants. Water Res 2007, 41, 613–619. [Google Scholar]
- Schwarzenlander, C; Averhoff, B. Characterization of DNA transport in the thermophilic. bacterium Thermus thermophilus HB27. FEBS 2006, 273, 4210–4218. [Google Scholar]
- Layden, N. An evaluation of autothermal thermophilic aerobic digestion (ATAD) of municipal slugde in Ireland. J. Environ. Eng. Sci 2007, 6, 19–29. [Google Scholar]
- Frydrych, I; Dziworska, G; Bilska, J. Comparative analysis of the thermal insulation properties of fabrics made of natural and man-made cellulose fibres. Fibres Text East Eur 2002, 10/12, 40–44. [Google Scholar]
- Pereira-Neto, JT; Stentiford, EI; Smith, DV. Survival of faecal indicator micro-organisms in refuse/sludge composting using the aerated static pile system. Waste Manage. Res 1986, 4, 397–406. [Google Scholar]
- Plym-Forshell, L. Survival of salmonellas and Ascaris suum eggs in a thermophilic biogas plant. Acta veter. Scand 1995, 36, 79–85. [Google Scholar]
- Shuval, H; Jodice, R; Consiglio, M; Spaggiarri, G; Spigoni, C. Control of enteric micro-organisms by aerobic-thermophilic co-composting of wastewater sludge and agro-industry wastes. Water Sci Technol 1991, 24, 401–405. [Google Scholar]
- Soares, HM; Cardenas, B; Weir, D; Switzenbaum, MS. Evaluating pathogen regrowth in biosolids compost. BioCycle 1995, 36, 70–74. [Google Scholar]
- Holmes, B; Willcox, WR; Lapage, SP. Identification of Enterobacteriaceae by the API 20E system. J. Clin. Pathol 1978, 31, 22–30. [Google Scholar]
- Nubel, U; Engelen, B; Felske, A; Snaidr, J; Wieshuber, A; Amann, RI; Ludwig, W; Backhaus, H. Sequence heterogeneities of genes encoding 16S rRNAs in Paenibacillus polymyxa detected by temperature gradient gel electrophoresis. J. Bacteriol 1996, 178, 5636–5643. [Google Scholar]
- Fisher, MM; Triplett, EW. Automated approaches for ribosomal intergenic spacer analysis of microbial diversity and its application to freshwater bacterial communities. Appl. Environ. Microbiol 1999, 65, 4630–4636. [Google Scholar]
- Nagpal, ML; Fox, KF; Fox, A. Utility of 16S-23S rRNA spacer region methodology: how similar are interspace regions within a genome and between strains for closely related organisms? J. Microbiol. Meth 1998, 33, 211–219. [Google Scholar]
- Yu, Z; Mohn, W. Bacterial Diversity and Community Structure in an Aerated Lagoon Revealed by Ribosomal Intergenic Spacer Analyses and 16S Ribosomal DNA Sequencing. Appl. Environ. Microbiol 2001, 67, 1565–1574. [Google Scholar]
- Scheinert, P; Krausse, R; Ullman, U; Soller, R; Krupp, G. Molecular differentiation of bacteria by PCR amplification of the 16S-23S rRNA spacer. J. Microbiol. Methods 1996, 26, 103–117. [Google Scholar]
- Sambrook, J; Fritschi, EF; Maniatis, T. Molecular Cloning: A Laboratory Manual; Cold Spring Harbor Laboratory Press: New York, NY, USA, 1989. [Google Scholar]
- McGrath, BM; O’Halloran, JA; Piterina, AV; Pembroke, JT. Molecular tools to detect the IncJ elements: A family of integrating, antibiotic resistant mobile genetic elements. J. Microbiol. Methods 2006, 66, 32–42. [Google Scholar]
- Piterina, AV; Bartlett, J; Pembroke, JT. Molecular Analysis of Bacterial Community DNA in Sludge Undergoing Autothermal Thermophilic Aerobic Digestion (ATAD): Pitfalls and Improved Methodology to Enhance Diversity Recovery. Diversity 2010, 2, 505–526. [Google Scholar]
- Sanger, F; Nicken, S; Couslon, AR. DNA sequencing with chain-terminating inhibitors. Biotechnol 1992, 24, 104–108. [Google Scholar]
- Altschul, SF; Madden, TL; Schaffer, AA; Zhang, J; Zhang, Z; Miller, W; Lipman, DJ. Gapped BLAST and PSI-BLAST: a new generation of protein database search programs. Nucleic Acids Res 1997, 25, 3389–3402. [Google Scholar]
- Stackebrandt, E; Goebel, BM. Taxonomic note: a place for DNA-DNA reassociation and 16S rRNA sequence analysis in the present species in bacteriology. Int. J. Syst. Bacteriol 1994, 44, 846–849. [Google Scholar]
- Drancourt, M; Bollet, C; Carlioz, A; Martelin, R; Gayral, JP; Raoult, D. 16S ribosomal DNA sequence analysis of a large collection of environmental and clinical unidentifiable bacterial isolates. J. Clin. Microbiol 2000, 38, 3623–3630. [Google Scholar]
- Randolph, MK; William, J. Fate of pathogens in thermophilic aerobic sludge digestion. Wat. Res 1982, 16, 1051–1060. [Google Scholar]
- Golueke, CG. When is compost “safe”? In The Art and Science of Composting; JG Press, Inc: Emmaus, PL, USA, 1991; pp. 220–229. [Google Scholar]
- Dumontet, S; Dinel, H; Baloda, SB. Pathogen reduction in sewage sludge by composting and other biological treatments: A review. Biol. Agr. Hortic 1999, 16, 409–430. [Google Scholar]
- Emerson, KR; Lund, RE; Thurston, RV. Aqueous ammonia equilibrium calculations: Effects of pH and temperature. J. Fish. Res. Board Can 1975, 32, 2379–2383. [Google Scholar]
- Park, GW; Diez-Gonzalez, F. Utilization of carbonate and ammonia based treatments to eliminate Escherichia coli O157:H7 and Salmonella 42 Typhimurium DT104 from cattle manure. J. Appl. Microbiol 2003, 94, 675–685. [Google Scholar]
- Mendez, JM; Jimenez, BE; Barrios, JA. Improved alkaline stabilization of municipal wastewater sludge. Water Sci. Technol 2002, 46, 139–146. [Google Scholar]
- Mendez, JM; Jimenez, B; Maya, C. Disinfection kinetics of pathogens in physicochemical sludge treated with ammonia. Water Sci. Technol 2004, 50, 67–74. [Google Scholar]
- Ottoson, J; Nordin, A; von Rosen, D; Vinnerås, B. Salmonella reduction in manure by the addition of urea and ammonia. Bioresource. Technol 2007, 99, 1610–1615. [Google Scholar]



{kind=link}
{kind=link}
| Primer | Specificity | Sequence (5’-3’) | Position | Th b, (°C) | Refs. |
|---|---|---|---|---|---|
| U968 a | Universal bacterial | AACGCGAAGAACCTTAC | 968–984 nt, 16S rDNA | 56 | [67] |
| L1401 | CGGTGTGTACAAGACCC | 1,385–1,401nt,16S rDNA | 56 | [67] | |
| S926f | CTYAAAKGAATTGACGG | 910 to 926 nt, 16S rDNA | 53 | [70] | |
| L189r | TACTGAGATGYTTMARTTC | 189–207 nt, 23S rDNA | 53 | [70] | |
| T7 | pGEM-TA vector | TAATACGACTCACTATAGGG | T7 promoter | 53 | Promega,UK |
| SP6 | ATTTAGGTGACACTATAG | SP6 promoter | 53 | Promega,UK | |
| Int Forw | Integrase gene, R391 ICE element | AACTAGGGCTGGGCTTATAA CATGGCC | ------- | 56 | [73] |
| Int Rev | AAAGATGGCAGCTTGCCGCA A CCTC | ------- | 56 | [73] |
| Samples utilized in this study | Temp (°C) | pH | Total solid content (%) |
|---|---|---|---|
| Feed (Pre thickened) | 11 | 6.3 | 6.3 |
| Reactor 1A (Stage 1) | 43 | 7.0 | 5.8 |
| Reactor 2A (2 h) (Stage 2 early) | 53 | 8.1 | 5.1 |
| Reactor2A (24 h)(Late Stage 2 prior to discharge) | 63 | 9.1 | 4.2 |
| Stabilised Product | 14 | 7.8 | 4.6 |
| Sample point | Fecal coliforms 1 (McConkey agar) | Total Coliforms (McConkey agar) | Fecal Coliforms (EMBA agar) | Total Coliforms (EMBA agar) |
|---|---|---|---|---|
| Primary Sludge | 3.6 × 105 | 6.1 × 107 | 2.1 × 104 | 8 × 106 |
| Secondary Sludge | 5.1 × 103 | 7.1 × 10 5 | 2 × 103 | 3.5 × 104 |
| ATAD Product | <1 | <1 | <1 | <1 |
| Sampling point | Fecal Coliforms | Total Coliforms | Salmonella spp. | Total Enterococci |
|---|---|---|---|---|
| Primary Sludge | 3.6 × 105 | 6.1 × 107 | 1.2 × 104 | 7.3 × 105 |
| Secondary Sludge | 5.1 × 103 | 7.1 × 10 5 | 8 × 102 | 9 × 103 |
| ATAD Product | Non detectable < 1 | Non detectable < 1 | Non detectable < 1 | Non detectable < 1 |
| Sample point | Fecal Coliforms (MPN/100 mL) | Total enteroccoci (MPN/100 mL) | Salmonella sp. |
|---|---|---|---|
| Product (March) | below detectable limit | below detectable limit | below detectable limit |
| Product (July) | below detectable limit | below detectable limit | below detectable limit |
| Product (October) | below detectable limit | below detectable limit | below detectable limit |
| Product (January) | below detectable limit | below detectable limit | below detectable limit |
| Detection limit | 20 1 | 20 | 10 |
| Sampling Point | Season | ||||
|---|---|---|---|---|---|
| Winter | Spring | Summer | Autumn | ||
| January | March | July | October | ||
| Inlet | ND1 | D | D | D | |
| Biosolids | Bulk water | ND | ND | ND | ND |
| Particulate matter | ND | ND | ND | ND | |
© 2010 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Piterina, A.V.; Bartlett, J.; Pembroke, T.J. Evaluation of the Removal of Indicator Bacteria from Domestic Sludge Processed by Autothermal Thermophilic Aerobic Digestion (ATAD). Int. J. Environ. Res. Public Health 2010, 7, 3422-3441. https://doi.org/10.3390/ijerph7093422
Piterina AV, Bartlett J, Pembroke TJ. Evaluation of the Removal of Indicator Bacteria from Domestic Sludge Processed by Autothermal Thermophilic Aerobic Digestion (ATAD). International Journal of Environmental Research and Public Health. 2010; 7(9):3422-3441. https://doi.org/10.3390/ijerph7093422
Chicago/Turabian StylePiterina, Anna V., John Bartlett, and Tony J. Pembroke. 2010. "Evaluation of the Removal of Indicator Bacteria from Domestic Sludge Processed by Autothermal Thermophilic Aerobic Digestion (ATAD)" International Journal of Environmental Research and Public Health 7, no. 9: 3422-3441. https://doi.org/10.3390/ijerph7093422
APA StylePiterina, A. V., Bartlett, J., & Pembroke, T. J. (2010). Evaluation of the Removal of Indicator Bacteria from Domestic Sludge Processed by Autothermal Thermophilic Aerobic Digestion (ATAD). International Journal of Environmental Research and Public Health, 7(9), 3422-3441. https://doi.org/10.3390/ijerph7093422