
Plant Seed Mucilage—Great Potential for Sticky Matter


Abstract

1. Introduction
2. Methodology
3. Ecological Functions of Mucilage
4. Effects of Mucilage on Human and Animal Health
5. Potential Uses of Mucilage in Agriculture and Industry
6. Physical and Chemical Properties of Mucilage
7. Functional Properties of Plant Seed Mucilage
8. Gene Regulation of Seed Mucilage Synthesis
9. Summary
10. Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Galloway, A.F.; Knox, P.; Krause, K. Sticky mucilages and exudates of plants: Putative microenvironmental design elements with biotechnological value. New Phytol. 2020, 225, 1461–1469. [Google Scholar] [CrossRef]
- Huang, D.; Wang, C.; Yuan, J.; Cao, J.; Lan, H. Differentiation of the seed coat and composition of the mucilage of Lepidium perfoliatum L.: A desert annual with typical myxospermy. Acta Biochim. Biophys. Sin. 2015, 47, 775–787. [Google Scholar] [CrossRef]
- Western, T. The sticky tale of seed coat mucilages: Production, genetics, and role in seed germination and dispersal. Seed Sci. Res. 2012, 22, 1–25. [Google Scholar] [CrossRef]
- Kreitschitz, A.; Gorb, S. The micro- and nanoscale spatial architecture of the seed mucilage—Comparative study of selected plant species. PLoS ONE 2018, 13, 0200522. [Google Scholar] [CrossRef]
- Paynel, F.; Pavlov, A.; Ancelin, G.; Rihouey, C.; Picton, L.; Lebrun, L.; Morvan, C. Polysaccharide hydrolases are released with mucilages after water hydration of flax seeds. Plant Physiol. Biochem. 2012, 62C, 54–62. [Google Scholar] [CrossRef]
- Yang, X.; Baskin, J.; Baskin, C.; Huang, Z.Y. More than just a coating: Ecological importance, taxonomic occurrence and phylogenetic relationships of seed coat mucilage. Perspect. Plant Ecol. Evol. Syst. 2012, 14, 434–442. [Google Scholar] [CrossRef]
- Kreitschitz, A.; Tadele, Z.; Gola, E. Slime cells on the surface of Eragrostis seeds maintain a level of moisture around the grain to enhance germination. Seed Sci. Res. 2009, 19, 27–35. [Google Scholar] [CrossRef]
- Gorai, M.; El Aloui, W.; Yang, X.; Neffati, M. Toward understanding the ecological role of mucilage in seed germination of a desert shrub Henophyton deserti: Interactive effects of temperature, salinity and osmotic stress. Plant Soil 2014, 374, 727–738. [Google Scholar] [CrossRef]
- Zhao, C.; Jiang, L.; Shi, X.; Wang, L. Mucilage inhibits germination of desert ephemeral Nepeta micrantha under moderate osmotic stress and promotes recovery after release of this stress. Seed Sci. Technol. 2020, 48, 21–25. [Google Scholar] [CrossRef]
- Yang, X.; Baskin, C.C.; Baskin, J.M.; Liu, G.; Huang, Z. Seed Mucilage Improves Seedling Emergence of a Sand Desert Shrub. PLoS ONE 2012, 7, e34597. [Google Scholar] [CrossRef]
- Pan, V.S.; Girvin, C.; LoPresti, E.F. Anchorage by seed mucilage prevents seed dislodgement in high surface flow: A mechanistic investigation. Ann. Bot. 2022, 129, 30–37. [Google Scholar] [CrossRef] [PubMed]
- LoPresti, E.; Pan, V.; Goidell, J.; Weber, M.; Karban, R. Mucilage-Bound Sand Reduces Seed Predation by Ants but Not by Reducing Apparency: A Field Test of 53 Plant Species. Bull. Ecol. Soc. Am. 2019, 100, e02809. [Google Scholar] [CrossRef]
- Geneve, R.; Hildebrand, D.; Phillips, T.; AL-Amery, M.; Kester, S. Stress Influences Seed Germination in Mucilage-Producing Chia. Crop Sci. 2017, 57, 2160–2169. [Google Scholar] [CrossRef]
- Zhou, Z.; Xing, J.; Zhao, J.; Liu, L.; Gu, L.; Lan, H. The ecological roles of seed mucilage on germination of Lepidium perfoliatum, a desert herb with typical myxospermy in Xinjiang. Plant Growth Regul. 2022, 97, 185–201. [Google Scholar] [CrossRef]
- El-Newary, S.A. Mucilage of Cordia dichotoma seeds pulp: Isolation, purification and a new hypolipidemic agent in normal and hyperlipidemic rats. Planta Med. 2015, 81, 107. [Google Scholar] [CrossRef]
- Kumar, D.; Pandey, J.; Kumar, P.; Raj, V. Psyllium Mucilage and Its Use in Pharmaceutical Field: An Overview. Curr. Synth. Syst. Biotechnol. 2017, 5, 1000134. [Google Scholar] [CrossRef]
- Rubilar, M.; Gutiérrez, C.; Verdugo, M.; Shene, C.; Sineiro, J. Flaxseed as a source of functional ingredients. J. Soil Sci. Plant Nutr. 2010, 10, 373–377. [Google Scholar] [CrossRef]
- Uddin Zim, A.F.M.I.; Khatun, J.; Khan, M.; Hossain, M.D.; Hauqe, M. Evaluation of in vitro antioxidant activity of okra mucilage and its antidiabetic and antihyperlipidemic effect in alloxan-induced diabetic mice. Food Sci. Nutr. 2021, 9, 6854–6865. [Google Scholar] [CrossRef]
- Palla, A.H.; Gilani, A. Dual effectiveness of Flaxseed in constipation and diarrhea: Possible mechanism. J. Ethnopharmacol. 2015, 169, 60–68. [Google Scholar] [CrossRef]
- Sindhu, G.; Ratheesh, M.; Shyni, G.L.; Nambisan, B.; Helen, A. Anti-inflammatory and antioxidative effects of mucilage of Trigonella foenum graecum (Fenugreek) on adjuvant induced arthritic rats. Int. Immunopharmacol. 2012, 12, 205–211. [Google Scholar] [CrossRef]
- Mueller, M.; Čavarkapa, A.; Unger, F.M.; Viernstein, H.; Praznik, W. Prebiotic potential of neutral oligo- and polysaccharides from seed mucilage of Hyptis suaveolens. Food Chem. 2017, 221, 508–514. [Google Scholar] [CrossRef] [PubMed]
- Muñoz, L.; Tamargo García, A.; Cueva, C.; Laguna, L.; Moreno Arribas, M.V. Understanding the impact of chia seed mucilage on human gut microbiota by using the dynamic gastrointestinal model simgi®. J. Funct. Foods 2018, 50, 104–111. [Google Scholar]
- Mkedder, I.; Bouali, W.; Hassaine, H. Antibacterial Activity of Mucilage of Linum usitatissimum L. Seeds. South Asian J. Exp. Biol. 2021, 11, 305–310. [Google Scholar] [CrossRef]
- Khan, A.A.; Alam, T.; Singh, S.; Wali, M.; Maaz, M.; Jabin, A. Efficacy Evaluation of Linium usitatissimum (Linctus of Flax Mucilage) in Chronic Obstructive Pulmonary Disease Patients. Planta Med. 2016, 82, PB20. [Google Scholar] [CrossRef]
- Tamri, P.; Hemmati, A.A.; Ghafourian, M. Wound healing properties of quince seed mucilage: In vivo evaluation in rabbit full-thickness wound model. Int. J. Surg. 2014, 12, 843–847. [Google Scholar] [CrossRef]
- Hemmati, A.A.; Kalantari, H.; Rezai, S.; Zadeh, H. Healing effect of quince seed mucilage on T-2 toxin-induced dermal toxicity in rabbit. Exp. Toxicol. Pathol. 2012, 64, 181–186. [Google Scholar] [CrossRef]
- Tantiwatcharothai, S.; Prachayawarakorn, J. Characterization of an antibacterial wound dressing from basil seed (Ocimum basilicum L.) mucilage-ZnO nanocomposite. Int. J. Biol. Macromol. 2019, 135, 133–140. [Google Scholar] [CrossRef]
- Sayyad, F.; Sakhare, S. Isolation, Characterization and Evaluation of Ocimum basilicum Seed Mucilage for Tableting Performance. Indian J. Pharm. Sci. 2018, 80, 282–290. [Google Scholar] [CrossRef]
- Saeedi, M.; Morteza-Semnani, K.; Ansoroudi, F.; Fallah, S.; Amin, G. Evaluation of binding properties of Plantago psyllium seed mucilage. Acta Pharm. 2010, 60, 339–348. [Google Scholar] [CrossRef]
- Avlani, D.; Ash, D.; Majee, S.; Roy Biswas, G. Sweet Basil Seed Mucilage as a Gelling agent in Nasal Drug Delivery. Int. J. Pharmtech. Res. 2019, 12, 42–49. [Google Scholar] [CrossRef]
- Atabaki, R.; Hassanpour, M. Improvement of Lidocaine Local Anesthetic Action Using Lallemantia royleana Seed Mucilage as an Excipient. Iran. J. Pharm. Sci. 2014, 13, 1431–1436. [Google Scholar]
- Tosif, M.M.; Najda, A.; Bains, A.; Kaushik, R.; Dhull, S.B.; Chawla, P.; Walasek-Janusz, M. A Comprehensive Review on Plant-Derived Mucilage: Characterization, Functional Properties, Applications, and Its Utilization for Nanocarrier Fabrication Polymers. Polymers 2021, 13, 1066. [Google Scholar] [CrossRef] [PubMed]
- Ribes, S.; Gómez, N.; Fuentes, A.; Talens, P.; Barat, J. Chia (Salvia hispanica L.) seed mucilage as a fat replacer in yogurts: Effect on their nutritional, technological, and sensory properties. J. Dairy Sci. 2021, 104, 2822–2833. [Google Scholar] [CrossRef] [PubMed]
- Basiri, S.; Haidary, N.; Shekarforoush, S.S.; Niakousari, M. Flaxseed mucilage: A natural stabilizer in stirred yogurt. Carbohydr. Polym. 2018, 187, 59–65. [Google Scholar] [CrossRef]
- Fernandes, S.; Salas Mellado, M. Development of Mayonnaise with Substitution of Oil or Egg Yolk by the Addition of Chia (Salvia hispânica L.) Mucilage. J. Food Sci. 2017, 83, 74–83. [Google Scholar] [CrossRef]
- Fernandes, S.; Filipini, G.; Salas Mellado, M. Development of cake mix with reduced fat and high practicality by adding chia mucilage. Food Biosci. 2021, 42, 101148. [Google Scholar] [CrossRef]
- Campos, B.; Ruivo, T.; Scapim, M.; Madrona, G.; Bergamasco, R. Optimization of the Mucilage Extraction Process from Chia Seeds and Application in Ice Cream as a Stabilizer and Emulsifier. Food Sci. Technol. 2015, 65, 874–883. [Google Scholar] [CrossRef]
- Dokoohaki, Z.; Sekhavatizadeh, S.; Hosseinzadeh, S. Dairy dessert containing microencapsulated Lactobacillus rhamnosus (ATCC 53103) with quince seed mucilage as a coating material. LWT 2019, 115, 108429. [Google Scholar] [CrossRef]
- Bustamante, M.; Oomah, B.D.; Rubilar, M.; Shene, C. Effective Lactobacillus plantarum and Bifidobacterium infantis encapsulation with chia seed (Salvia hispanica L.) and flaxseed (Linum usitatissimum L.) mucilage and soluble protein by spray drying. Food Chem. 2016, 216, 97–105. [Google Scholar] [CrossRef]
- Kurd, F.; Fathi, M.; Shekarchizadeh, H. Nanoencapsulation of hesperetin using basil seed mucilage nanofibers: Characterization and release modeling. Food Biosci. 2019, 32, 100475. [Google Scholar] [CrossRef]
- Fahami, A.; Fathi, M. Development of cress seed mucilage/PVA nanofibers as a novel carrier for vitamin A delivery. Food Hydrocoll. 2018, 81, 31–38. [Google Scholar] [CrossRef]
- Hajivand, P.; Aryanejad, S.; Akbari, I.; Hemmati, A. Fabrication and characterization of a promising oregano-extract/psyllium-seed mucilage edible film for food packaging. J. Food Sci. 2020, 85, 2481–2490. [Google Scholar] [CrossRef]
- da Silveira Ramos, I.F.; Magalhães, L.M.; do OPessoa, C.; Ferreira, P.M.; dos Santos Rizzo, M.; Osajima, J.A.; Silva-Filho, E.C.; Nunes, C.; Raposo, F.; Coimbra, M.A.; et al. New properties of chia seed mucilage (Salvia hispanica L.) and potential application in cosmetic and pharmaceutical products. Ind. Crops Prod. 2021, 171, 113981. [Google Scholar] [CrossRef]
- Naveed, M.; Ahmed, M.A.; Benard, P.; Brown, L.K.; George, T.S.; Bengough, A.G.; Roose, T.; Koebernick, N.; Hallett, P.D. Surface tension, rheology and hydrophobicity of rhizodeposits and seed mucilage influence soil water retention and hysteresis. Plant Soil 2019, 437, 65–81. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.; Zheng, R.; Shi, X.; Wang, L. Soil microbes and seed mucilage promote growth of the desert ephemeral plant Nepeta micrantha under different water conditions. Flora Morphol. Distrib. Funct. Ecol. Plants 2021, 280, 151845. [Google Scholar] [CrossRef]
- Paynel, F.; Morvan, C.; Marais, S.; Lebrun, L. Improvement of the hydrolytic stability of new flax-based biocomposite materials. Polym. Degrad. Stab. 2013, 98, 190–197. [Google Scholar] [CrossRef]
- Ellerbrock, R.; Ahmed, M.; Gerke, H. Spectroscopic characterization of mucilage (Chia seed) and polygalacturonic acid. J. Soil Sci. Plant Nutr. 2019, 182, 888–895. [Google Scholar] [CrossRef]
- Kaur, M.; Kaur, R.; Punia, S. Characterization of mucilages extracted from different flaxseed (Linum usitatissiumum L.) cultivars: A heteropolysaccharide with desirable functional and rheological properties. Int. J. Biol. Macromol. 2018, 117, 917–927. [Google Scholar] [CrossRef]
- Nazari, M. Plant mucilage components and their functions in the rhizosphere. Rhizosphere 2021, 18, 100344. [Google Scholar] [CrossRef]
- Kreitschitz, A.; Gorb, S. How does the cell wall ‘stick’ in the mucilage? A detailed microstructural analysis of the seed coat mucilaginous cell wall. Flora 2017, 229, 9–22. [Google Scholar] [CrossRef]
- Oomah, B.D.; Kenaschuk, E.; Cui, S.; Mazza, G. Variation in the composition of water-soluble polysaccharides in flaxseed. J. Agric. Food Chem. 1995, 43, 1484–1488. [Google Scholar] [CrossRef]
- Porokhovinova, E.; Pavlov, A.V.; Brutch, N.; Morvan, C. Carbohydrate composition of flax mucilage and its relation to morphological characters. Agric. Biol. 2017, 52, 161–171. [Google Scholar] [CrossRef]
- Liu, Y.; Liu, Z.; Zhu, X.; Hu, X.; Zhang, H.; Guo, Q.; Yada, R.Y.; Cui, S.W. Seed coat mucilages: Structural, functional/bioactive properties, and genetic information. Compr. Rev. Food Sci. 2021, 20, 2534–2559. [Google Scholar] [CrossRef] [PubMed]
- Alizadeh Behbahani, B.; Imani Fooladi, A.A. Shirazi balangu (Lallemantia royleana) seed mucilage: Chemical composition, molecular weight, biological activity and its evaluation as edible coating on beefs. Int. J. Biol. Macromol. 2018, 114, 882–889. [Google Scholar] [CrossRef]
- Razavi, S.; Cui, S.; Ding, H. Structural and physicochemical characteristics of a novel water-soluble gum from Lallemantia royleana seed. Int. J. Biol. Macromol. 2016, 83, 142–151. [Google Scholar] [CrossRef]
- Naji-Tabasi, S.; Razavi, S.M.A.; Mohebbi, M.; Malaekeh-Nikouei, B. New studies on basil (Ocimum bacilicum L.) seed gum: Part I—Fractionation, physicochemical and surface activity characterization. Food Hydrocoll. 2016, 52, 350–358. [Google Scholar] [CrossRef]
- Timilsena, Y.; Adhikari, R.; Kasapis, S.; Adhikari, B. Molecular and functional characteristics of purified gum from Australian chia seeds. Carbohydr. Polym. 2016, 136, 128–136. [Google Scholar] [CrossRef]
- Lin, K.Y.; Daniel, J.; Whistler, R. Structure of chia seed polysaccharide exudate. Carbohyd. Polym. 1994, 23, 13–18. [Google Scholar] [CrossRef]
- Praznik, W.; Čavarkapa, A.; Unger, F.M.; Loeppert, R.; Holzer, W.; Viernstein, H.; Mueller, M. Molecular dimension and structural features of neutral polysaccharides from the seed mucilage of Hyptis suaveolens L. Food Chem. 2016, 221, 1997–2004. [Google Scholar] [CrossRef]
- Karazhiyan, H.; Razavi, S.; Phillips, G.; Fang, Y.; Al-Assaf, S.; Nishinari, K. Rheological properties of Lepidium sativum seed extract as a function of concentration, temperature and time. Food Hydrocoll. 2009, 23, 2062–2068. [Google Scholar] [CrossRef]
- Alpizar Reyes, E.; Carrillo Navas, H.; Gallardo Rivera, R.; Varela Guerrero, V.; Alvarez Ramirez, J.; Pérez Alonso, C. Functional properties and physicochemical characteristics of tamarind (Tamarindus indica L.) seed mucilage powder as a novel hydrocolloid. J. Food Eng. 2017, 209, 68–75. [Google Scholar] [CrossRef]
- Koocheki, A.; Razavi, S.M.A.; Hesarinejad, M.A. Effect of Extraction Procedures on Functional Properties of Eruca sativa Seed Mucilage. Food Biophys. 2012, 7, 84–92. [Google Scholar] [CrossRef]
- Yaseen, E.I.; Herald, T.J.; Aramouni, F.M.; Alavi, S. Rheological properties of selected gum solutions. Food Res. Int. 2005, 38, 111–119. [Google Scholar] [CrossRef]
- Naji-Tabasi, S.; Razavi, S.M.A. New studies on basil (Ocimum bacilicum L.) seed gum: Part II—Emulsifying and foaming characterization. Carbohydr. Polym. 2016, 149, 140–150. [Google Scholar] [CrossRef] [PubMed]
- Qian, K.; Cui, S.; Wu, Y.; Goff, H.D. Flaxseed gum from flaxseed hulls: Extraction, fractionation, and characterization. Food Hydrocoll. 2012, 28, 275–283. [Google Scholar] [CrossRef]
- Naji-Tabasi, S.; Razavi, S. Functional properties and applications of basil seed gum: An overview. Food Hydrocoll. 2017, 73, 313–325. [Google Scholar] [CrossRef]
- Muñoz, L.; Natalia, C.; Zúñiga-López, M.; Moncada-Basualto, M.; Haros, C.M. Physicochemical and functional properties of soluble fiber extracted from two phenotypes of chia (Salvia hispanica L.) seeds. J. Food Compos. Anal. 2021, 104, 104138. [Google Scholar] [CrossRef]
- Koocheki, A.; Taherian, A.; Bostan, A. Studies on the steady shear flow behavior and functional properties of Lepidium perfoliatum seed gum. Food Res. Int. 2013, 50, 446–456. [Google Scholar] [CrossRef]
- Rashid, F.; Ahmed, Z.; Hussain, S.; Huang, J.Y.; Ahmad, A. Linum usitatissimum L. seeds: Flax gum extraction, physicochemical and functional characterization. Carbohydr. Polym. 2019, 215, 29–38. [Google Scholar] [CrossRef]
- Nazir, S.; Wani, I.A. Functional characterization of basil (Ocimum basilicum L.) seed mucilage. Bioact. Carbohydr. Diet. Fibre. 2021, 25, 100261. [Google Scholar] [CrossRef]
- Jouki, M.; Mortazavi, S.; Tabatabaee, F.; Koocheki, A. Optimization of extraction, antioxidant activity and functional properties of quince seed mucilage by RSM. Int. J. Biol. Macromol. 2014, 66, 113–124. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.H.; Xu, S.Y.; Wang, Z. Gelation properties of flaxseed gum. Int. J. Food Eng. 2006, 77, 295–303. [Google Scholar] [CrossRef]
- Pérez-Orozco, J.; Sanchez-Herrera, L.; Ortiz Basurto, R. Effect of concentration, temperature, pH, co-solutes on the rheological properties of Hyptis suaveolens L. mucilage dispersions. Food Hydrocoll. 2018, 87, 297–306. [Google Scholar] [CrossRef]
- Wannerberger, K.; Nylander, T.; Nyman, M. Rheological and Chemical Properties of Mucilage in Different Varieties from Linseed (Linum usitatissimum). Acta Agric. Scand. 1991, 41, 311–319. [Google Scholar] [CrossRef]
- Ben-Tov, D.; Idan-Molakandov, A.; Hugger, A.; Ben-Shlush, I.; Günl, M.; Yang, B.; Usadel, B.; Harpaz-Saad, S. The role of COBRA-LIKE 2 function, as part of the complex network of interacting pathways regulating Arabidopsis seed mucilage polysaccharide matrix organization. Plant J. 2018, 94, 497–512. [Google Scholar] [CrossRef]
- Dean, G.; Zheng, H.; Tewari, J.; Huang, J.; Young, D.; Hwang, Y.; Western, T.; Carpita, N.; McCann, M.; Mansfield, S.; et al. The Arabidopsis MUM2 Gene Encodes a β-Galactosidase Required for the Production of Seed Coat Mucilage with Correct Hydration Properties. Plant Cell 2018, 19, 4007–4021. [Google Scholar] [CrossRef]
- McGee, R.; Dean, G.H.; Mansfield, S.D.; Haughn, G.W. Assessing the utility of seed coat-specific promoters to engineer cell wall polysaccharide composition of mucilage. Plant Mol. Biol. 2019, 101, 373–387. [Google Scholar] [CrossRef]
- Dean, G.H.; Jin, Z.; Shi, L.; Esfandiari, E.; McGee, R.; Nabata, K.; Lee, T.; Kunst, L.; Western, T.L.; Haughn, G.W. Identification of a seed coat-specific promoter fragment from the Arabidopsis MUCILAGE-MODIFIED4 gene. Plant Mol. Biol. 2017, 95, 33–50. [Google Scholar] [CrossRef]
- Zhang, Y.; Yin, Q.; Qin, W.; Gao, H.; Du, J.; Chen, J.; Li, H.; Zhou, G.; Wu, H.; Wu, A. The Class II KNOX family members KNAT3 and KNAT7 redundantly participate in Arabidopsis seed coat mucilage biosynthesis. J. Exp. Bot. 2022, 73, 3477–3495. [Google Scholar] [CrossRef]
- Fabrissin, I.; Cueff, G.; Berger, A.; Granier, F.; Sallé, C.; Poulain, D.; Ralet, M.C.; North, H. Natural Variation Reveals a Key Role for Rhamnogalacturonan I in Seed Outer Mucilage and Underlying Genes. Plant Physiol. 2019, 181, 1498–1518. [Google Scholar] [CrossRef]
- Huang, J.; DeBowles, D.; Esfandiari, E.; Dean, G.; Carpita, N.; Haughn, G. The Arabidopsis Transcription Factor LUH/MUM1 Is Required for Extrusion of Seed Coat Mucilage. Plant Physiol. 2011, 156, 491–502. [Google Scholar] [CrossRef] [PubMed]
- Kong, Y.; Zhou, G.; Abdeen, A.; Schafhauser, J.; Richardson, B.; Atmodjo, M.; Jung, J.; Wicker, L.; Mohnen, D.; Western, T.L.; et al. AtGATL5 is Involved in the Production of Arabidopsis Seed Coat Mucilage. Plant Physiol. 2013, 163, 1203–1217. [Google Scholar] [CrossRef] [PubMed]
- Saez-Aguayo, S.; Rautengarten, C.; Temple, H.; Sanhueza, D.; Ejsmentewicz, T.; Sandoval-Ibañez, O.; Doñas, D.; Parra-Rojas, J.P.; Ebert, B.; Lehner, A.; et al. UUAT1 Is a Golgi-Localized UDP-Uronic Acid Transporter That Modulates the Polysaccharide Composition of Arabidopsis Seed Mucilage. Plant Cell 2017, 29, 129–143. [Google Scholar] [CrossRef] [PubMed]
- Griffiths, J.; Crepeau, M.-J.; Ralet, M.-C.; Seifert, G.; North, H. Dissecting seed mucilage adherence mediated by FEI2 and SOS5. Front. Plant Sci. 2016, 7, e0145092. [Google Scholar] [CrossRef] [PubMed]
- Saez-Aguayo, S.; Ralet, M.C.; Berger, A.; Botran, L.; Ropartz, D.; Marion-Poll, A.; North, H. PECTIN METHYLESTERASE INHIBITOR6 promotes Arabidopsis mucilage release by limiting methylesterification of homogalacturonan in seed coat epidermal cells. Plant Cell 2013, 25, 308–323. [Google Scholar] [CrossRef]
- Soto-Cerda, B.; Cloutier, S.; Quian Ulloa, R.; Gajardo Balboa, H.; Olivos, M.; You, F. Genome-Wide Association Analysis of Mucilage and Hull Content in Flax (Linum usitatissimum L.) Seeds. Int. J. Mol. Sci. 2018, 19, 2870. [Google Scholar] [CrossRef]
- Turbant, A.; Fournet, F.; Lequart-Pillon, M.; Zabijak, L.; Pageau, K.; Bouton, S.; Van Wuytswinkel, O. PME58 plays a role in pectin distribution during seed coat mucilage extrusion through homogalacturonan modification. J. Exp. Bot. 2016, 67, 2177–2190. [Google Scholar] [CrossRef]
- Xu, Y.; Wang, Y.; Wang, X.; Pei, S.; Kong, Y.; Hu, R.; Zhou, G. Transcription Factors BLH2 and BLH4 Regulate Demethylesterification of Homogalacturonan in Seed Mucilage. Plant Physiol. 2020, 183, 96–111. [Google Scholar] [CrossRef]
- Sun, J.; Yuan, C.; Wang, M.; Ding, A.; Chai, G.; Sun, Y.; Zhou, G.; Yang, D.H.; Kong, Y. MUD1, a RING-v E3 ubiquitin ligase, has an important role in the regulation of pectin methylesterification in Arabidopsis seed coat mucilage. Plant Physiol. Biochem. 2021, 168, 230–238. [Google Scholar] [CrossRef]
- Han, X.; Zhang, L.; Niu, D.; Shuzhen, N.; Miao, X.; Hu, X.; Li, C.; Fu, H. Transcriptome and co-expression network analysis reveal molecular mechanisms of mucilage formation during seed development in Artemisia sphaerocephala. Carbohydr. Polym. 2021, 251, 117044. [Google Scholar] [CrossRef]
- Han, X.; Zhang, L.; Miao, X.; Hu, X.; Shuzhen, N.; Fu, H. Transcriptome analysis reveals the molecular mechanisms of mucilage biosynthesis during Artemisia sphaerocephala seed development. Ind. Crops Prod. 2020, 145, 111991. [Google Scholar] [CrossRef]
- Cao, J.; Xu, D.; Huang, D.; Yuan, J.; Zhao, J.; Wang, W.; Lan, H. Cloning, characterization, and functional analysis of seed coat mucilage-related gene TTG1 from Lepidium perfoliatum. Plant Sci. J. 2014, 32, 371–382. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
| Application Area | Plant Source | Applied Form | Achieved Properties | Reference |
|---|---|---|---|---|
| Food industry | Salvia hispanica L., Linum usitatissimum L. | Additive in yogurts | Improved nutritional properties, syneresis and viscosity | Refs. [33,34] |
| Salvia hispanica L. | Additive in mayonnaise | Increased stability, reducing fat | Ref. [35] | |
| Salvia hispanica L. | Additive in cakes | Improved nutritional qualities | Ref. [36] | |
| Salvia hispanica L. | Additive in ice cream | Replacement for stabilizers and emulsifiers | Ref. [37] | |
| Salvia hispanica L. Linum usitatissimum L. Cydonia oblonga Miller | Encapsulation of probiotics | Better resistance in the digestive tract | Refs. [38,39] | |
| Ocimum basilicum L. Lepidium sativum L. | Encapsulation of vitamins and flavonoids | Better resistance in the digestive tract | Refs. [40,41] | |
| Plantago psyllium L. | Production of edible films | Increased food shelf life | Ref. [42] | |
| Pharmaceutical industry | Lallemantia royleana (Benth.) | Formation of gels | Healing effects against dermal toxicity and burns | Ref. [31] |
| Ocimum basilicum L. | Wound dressing formation | Antimicrobial effects | Ref. [27] | |
| Ocimum basilicum L. Plantago psyllium L. | Formation of medicinal tablets | Slower release, replacement of chemical preparations | Refs. [28,29] | |
| Ocimum basilicum L. | Formation of nasal gel | Analgesic effects | Ref. [30] | |
| Cosmetics | Salvia hispanica L. | Gel formation | UV-protective effects | Ref. [43] |
| Agriculture | Salvia hispanica L. | Hydrogels in arid areas | Retention of water | Refs. [44,45] |
| Engineering industry | Linum usitatissimum L. | Biocomposite binder | Inexpensive and biocompound | Ref. [46] |
| Plant Source of Seed Mucilage | Carbohydrates | Reference |
|---|---|---|
| Linum usitatissimum L. | Rhamnogalacturonan and arabinoxylan | Ref. [5] |
| Linum usitatissimum L. | D-xylose, L-arabinose, D-galactose, L-ramnose, L-fucose, L-galactose, D-galacturonic acid | Ref. [51] |
| Lepidium perfoliatum L. | Methylesterified homogalacturonan, callose, hemicellulose | Ref. [2] |
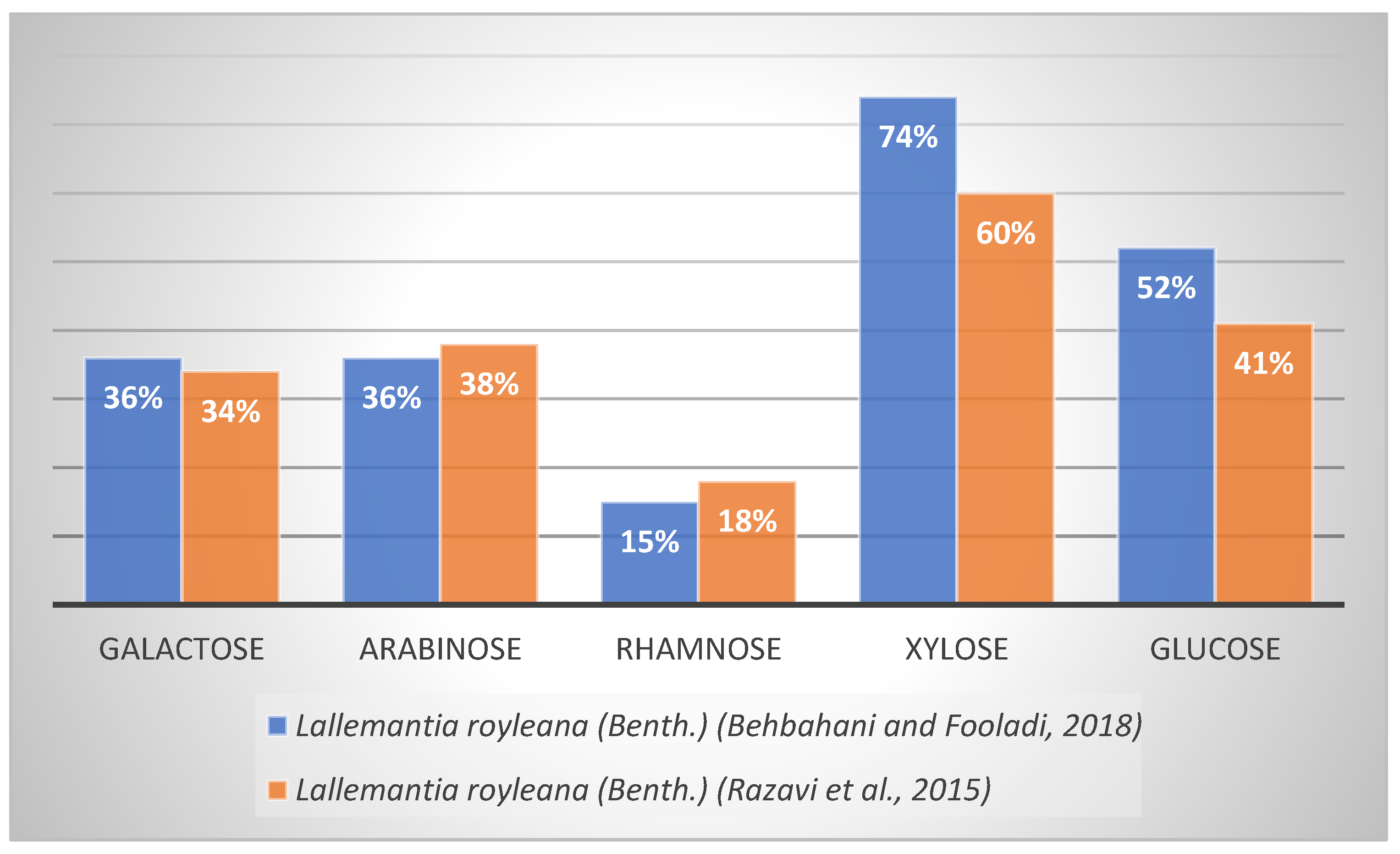
| Lallemantia royleanaBenth. | Galactose, arabinose, rhamnose, xylose, glucose | Refs. [54,55] |
| Ocimum basilicum L. | Glucose, galactose, mannose, arabinose, xylose, rhamnose | Ref. [56] |
| Salvia hispanica L. | Xylose, glucose, arabinose, galactose, glucuronic acid, galacturonic acid | Ref. [57] |
| Salvia hispanica L. | Residues of D-xylosyl, D-glucosyl, 4-0-methyl-D-glucuronopyranosyl | Ref. [58] |
| Hyptis suaveolens L. | Galactose, glucose, mannose, galactoglucan, galactoglucomannan, fucose, xylose, 4-O-methylglucuronic acid | Refs. [21,59] |
| Lepidium sativum L. | Mannose, arabinose, galacturonic acid, fructose, glucuronic acid, galactose, rhamnose, glucose | Ref. [60] |
| Function in the Process | Genes/Transcription Factors | Reference |
|---|---|---|
| Mucilage synthesis and release | Transparent testa 8; subtilisin-like serine protease; galacturosyl transferase-like 5; mucilage-modified 4; agamous-like MADS-box protein AGL62; glycosyl hydrolase family 17; pectin methylesterase inhibitor 6 | Refs. [85,86] |
| Mucilage amount | Mucilage-modified 2 (MUM2) | Ref. [76] |
| Mucilage proper structure | Mucilage-modified 2 (MUM2) | Ref. [76] |
| Mucilage polysaccharide composition | Mucilage-modified 2 (MUM2) + testa-abundant 2 (TBA2); peroxidase 36 (PER36); mucilage-modified 4 (MUM4) | Ref. [77] |
| Mucilage production | Knotted arabidopsis thaliana 3 (KNAT3) and knotted arabidopsis thaliana 7 (KNAT7) | Ref. [79] |
| Mucilage cellulose deposition | Cobra-like 2 (COBL2) | Ref. [75] |
| Mucilage composition | UDP-uronic acid transporter1 (UUAT 1) | Ref. [83] |
| Mucilage extrusion | Leunig homolog (LUH)/mucilage-modified 1 (MUM 1); enzymes MUM 2; subsilin protease 1.7; beta-xylosidase 1 | Ref. [81] |
| Mucilage adherence | Cellulose synthase 5 (CESA5)/mucilage-modified 3 (MUM3) | Ref. [84] |
| Mucilage structure and organization | Pectin methylesterase 8 (PME 8) + BLH 2 and BLH 4 | Refs. [87,88] |
| Mucilage rhamnogalacturonan I size | Mucilage-related 70 (MUCI 70); galacturonosyltransferase-like 5 (GATL 5) | Ref. [80] |
| Mucilage rhamnogalacturonan I amount | Copper amine oxidase 1 (CuAOX 1) | Ref. [80] |
| Spatial Localization | Genes/Transcription Factors | Reference |
|---|---|---|
| Epidermal cells | Cobra-like 2 (COBL2) | Ref. [75] |
| Cell wall | Mucilage-modified 2 (MUM2) | Ref. [76] |
| Seed coat cells | Mucilage-modified 4 (MUM4) | Ref. [77] |
| Mucilage-secreting cells | BLH 2 and BLH 4 | Ref. [88] |
| Nucleus | Leunig homolog LUH | Ref. [81] |
| Endoplasmic reticulum; Golgi apparatus | Galacturonosyltransferase-like 5 (GATL5) | Ref. [82] |
| Golgi apparatus | UDP-uronic acid transporter1 (UUAT 1) | Ref. [83] |
| Developing seed coat | Mucilage defect 1 (MUD1) | Ref. [89] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kučka, M.; Ražná, K.; Harenčár, Ľ.; Kolarovičová, T. Plant Seed Mucilage—Great Potential for Sticky Matter. Nutraceuticals 2022, 2, 253-269. https://doi.org/10.3390/nutraceuticals2040019
Kučka M, Ražná K, Harenčár Ľ, Kolarovičová T. Plant Seed Mucilage—Great Potential for Sticky Matter. Nutraceuticals. 2022; 2(4):253-269. https://doi.org/10.3390/nutraceuticals2040019
Chicago/Turabian StyleKučka, Matúš, Katarína Ražná, Ľubomír Harenčár, and Terézia Kolarovičová. 2022. "Plant Seed Mucilage—Great Potential for Sticky Matter" Nutraceuticals 2, no. 4: 253-269. https://doi.org/10.3390/nutraceuticals2040019
APA StyleKučka, M., Ražná, K., Harenčár, Ľ., & Kolarovičová, T. (2022). Plant Seed Mucilage—Great Potential for Sticky Matter. Nutraceuticals, 2(4), 253-269. https://doi.org/10.3390/nutraceuticals2040019