
Effects of Xylitol on Tumor Progression in Syngeneic Mice Cancer Models

 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Mice and Study Design
Euthanization Protocol
2.2. Sample Collection
2.3. Metabolomic Analysis Procedure
3. Results
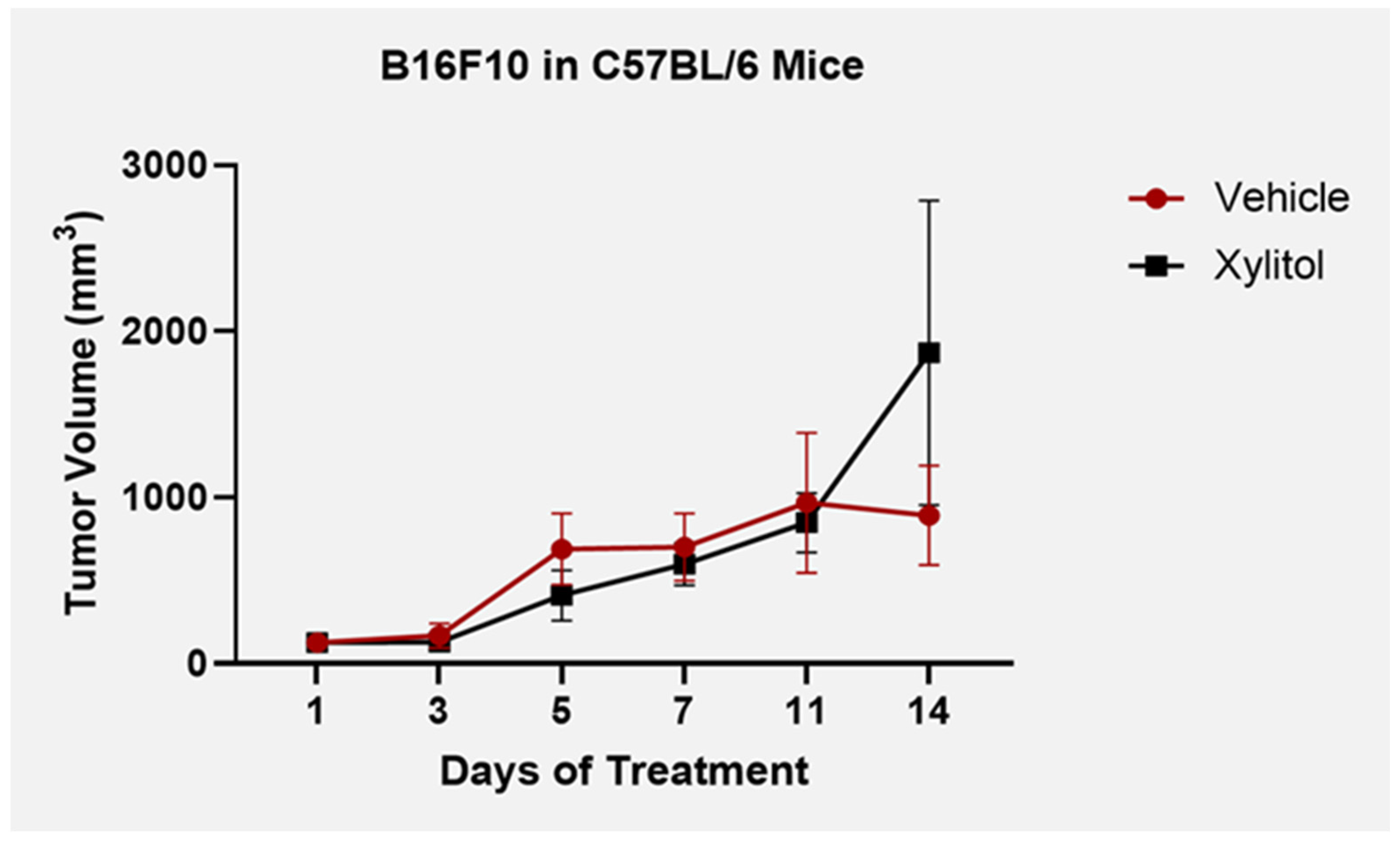
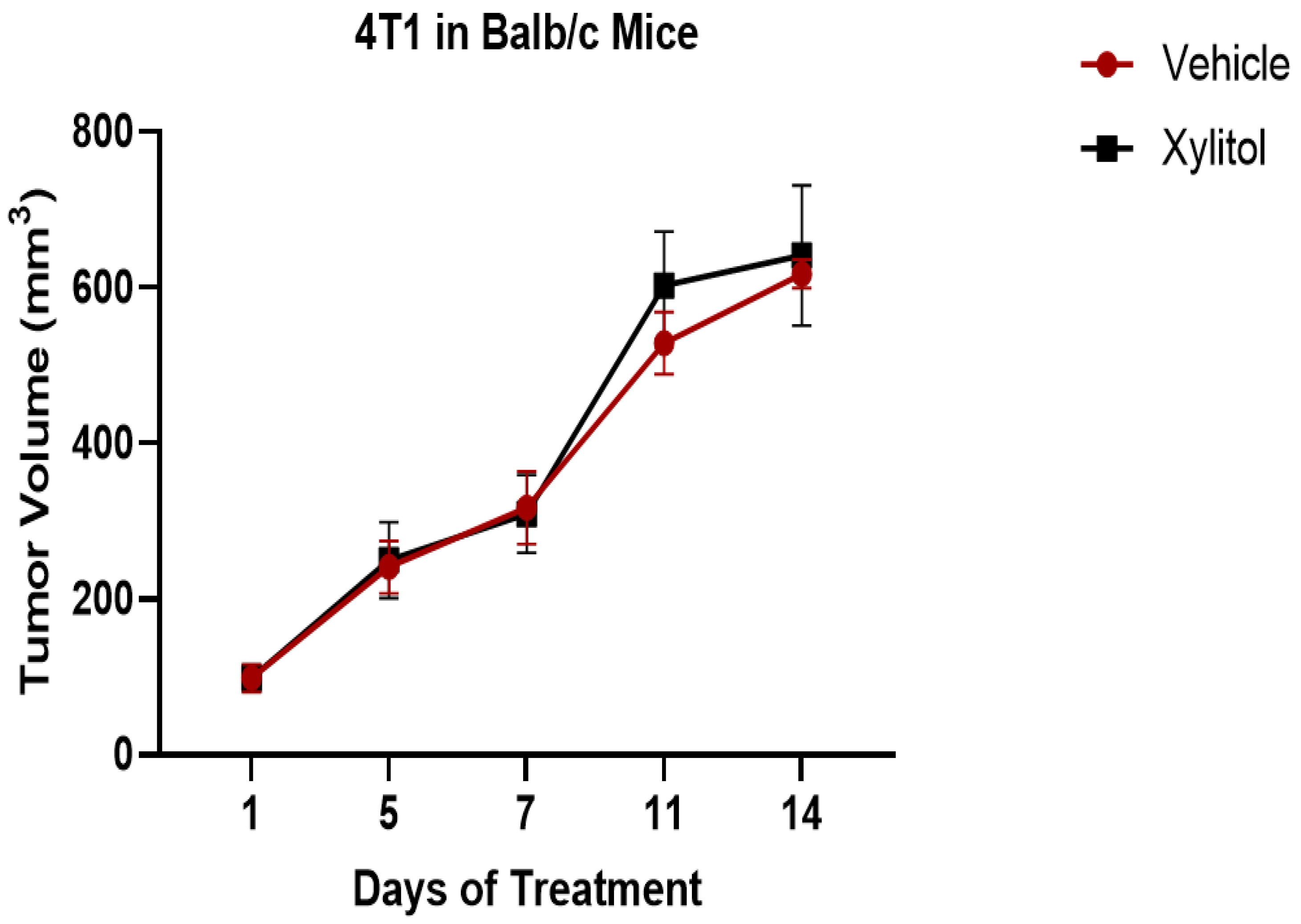
3.1. Tumor Volume
3.2. Metabolomic Analysis
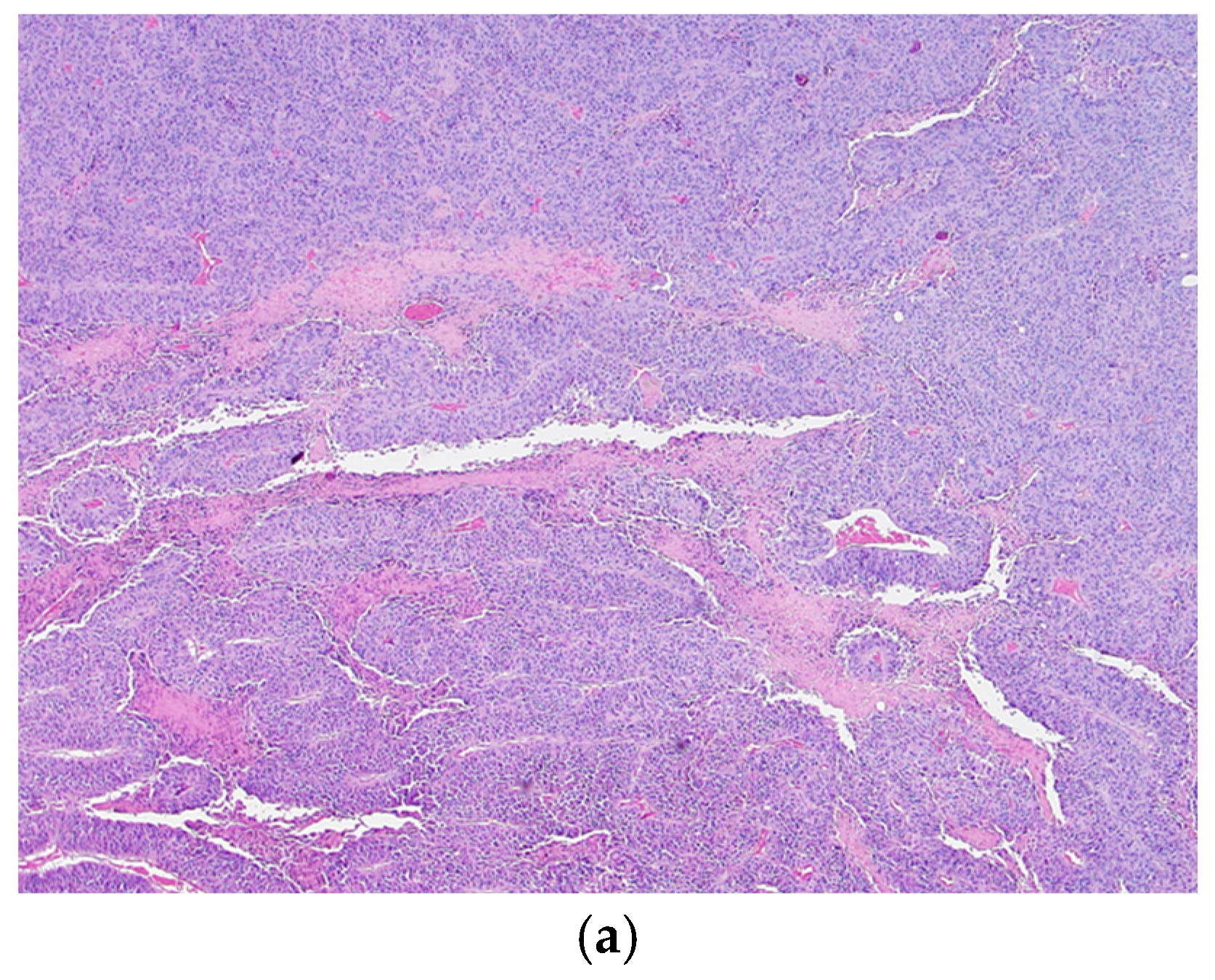
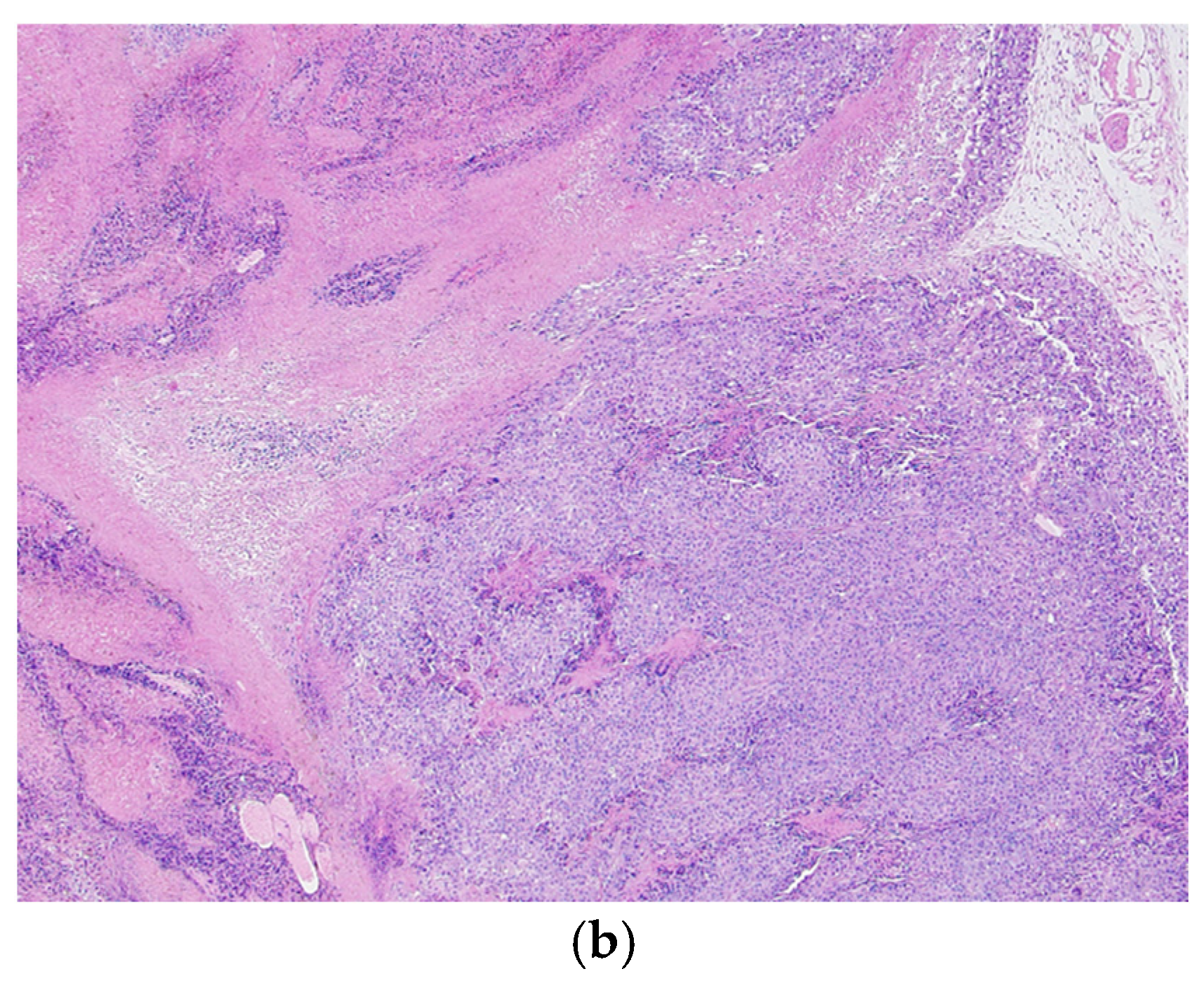
3.3. Histological Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bray, F.; Laversanne, M.; Sung, H.; Ferlay, J.; Siegel, R.L.; Soerjomataram, I.; Jemal, A. Global cancer statistics 2022: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2024, 74, 229–263. [Google Scholar] [CrossRef] [PubMed]
- Vallano, A.; Pontes, C. Escalating costs of innovative medicines: Perspective and proposals. Front. Public Health 2024, 12, 1449707. [Google Scholar] [CrossRef] [PubMed]
- Nayak, P.A.; Nayak, U.A.; Khandelwal, V. The effect of xylitol on dental caries and oral flora. Clin. Cosmet. Investig. Dent. 2014, 6, 89–94. [Google Scholar] [CrossRef] [PubMed]
- Park, E.; Park, M.H.; Na, H.S.; Chung, J. Xylitol induces cell death in lung cancer A549 cells by autophagy. Bio-Technol. Lett. 2015, 37, 983–990. [Google Scholar] [CrossRef] [PubMed]
- Tomonobu, N.; Komalasari, N.L.G.Y.; Sumardika, I.W.; Jiang, F.; Chen, Y.; Yamamoto, K.I.; Kinoshita, R.; Murata, H.; Inoue, Y.; Sakaguchi, M. Xylitol acts as an anticancer monosaccharide to induce selective cancer death via regulation of the glutathione level. Chem. Interact. 2020, 324, 109085. [Google Scholar] [CrossRef]
- Sahasakul, Y.; Angkhasirisap, W.; Lam-Ubol, A.; Aursalung, A.; Sano, D.; Takada, K.; Trachootham, D. Partial Substitution of Glucose with Xylitol Prolongs Survival and Suppresses Cell Proliferation and Glycolysis of Mice Bearing Orthotopic Xenograft of Oral Cancer. Nutrients 2022, 14, 2023. [Google Scholar] [CrossRef]
- Mehnert, H.; Förster, H.; Dehmel, K.H. The effect of intravenous administration of xylitol solutions in normal persons and in patients with liver diseases and diabetes mellitus. In Proceedings of the International Symposium on Metabolism, Physiology, and Clinical Use of Pentoses and Pentitols, Hakone, Japan, 27–29 August 1967; Springer: Berlin/Heidelberg, Germany, 1969; pp. 293–302. [Google Scholar]
- Sato, J.; Wang, Y.M.; van Eys, J. Metabolism of xylitol and glucose in rats bearing hepatocellular carcinomas. Cancer Res. 1981, 41, 3192–3199. [Google Scholar] [PubMed]
- Yi, E.Y.; Kim, Y.J. Xylitol inhibits in vitro and in vivo angiogenesis by suppressing the NF-κB and Akt signaling pathways. Int. J. Oncol. 2013, 43, 315–320. [Google Scholar] [CrossRef] [PubMed]
- Kharaeva, Z.F.; Mustafaev, M.S.; Khazhmetov, A.V.; Gazaev, I.H.; Blieva, L.Z.; Steiner, L.; Mayer, W.; Luca, C.; Korkina, L.G. Anti-Bacterial and Anti-Inflammatory Effects of Toothpaste with Swiss Medicinal Herbs towards Patients Suffering from Gingivitis and Initial Stage of Periodontitis: From Clinical Efficacy to Mechanisms. Dent. J. 2020, 8, 10. [Google Scholar] [CrossRef]
- Park, E.; Na, H.S.; Kim, S.M.; Wallet, S.; Cha, S.; Chung, J. Xylitol, an anticaries agent, exhibits potent inhibition of inflammatory responses in human THP-1-derived macrophages infected with Porphyromonas gingivalis. J. Periodontol. 2014, 85, e212–e223. [Google Scholar] [CrossRef]
- Rinschen, M.M.; Ivanisevic, J.; Giera, M.; Siuzdak, G. Identification of bioactive metabolites using activity metabolomics. Nat. Rev. Mol. Cell Biol. 2019, 20, 353–367. [Google Scholar] [CrossRef] [PubMed]
- Vander Heiden, M.G.; DeBerardinis, R.J. Understanding the Intersections between Metabolism and Cancer Biology. Cell 2017, 168, 657–669. [Google Scholar] [CrossRef] [PubMed]
- Trachootham, D.; Chingsuwanrote, P.; Yoosadiang, P.; Mekkriangkrai, D.; Ratchawong, T.; Buraphacheep, N.; Kijanukul, S.; Saekhow, S.; Pongpitchayadej, O.; Vongvachvasin, K.; et al. Partial substitution of glucose with xylitol suppressed the glycolysis and selectively inhibited the proliferation of oral cancer cells. Nutr. Cancer 2017, 69, 862–872. [Google Scholar] [CrossRef] [PubMed]
- Qusa, M.H.; Siddique, A.B.; Nazzal, S.; El Sayed, K.A. Novel olive oil phenolic (−)-oleocanthal (+)-xylitol-based solid dispersion formulations with potent oral anti-breast cancer activities. Int. J. Pharm. 2019, 569, 118596. [Google Scholar] [CrossRef]
- Xiang, S.; Ye, K.; Li, M.; Ying, J.; Wang, H.; Han, J.; Shi, L.; Xiao, J.; Shen, Y.; Feng, X.; et al. Xylitol enhances synthesis of propionate in the colon via cross-feeding of gut microbiota. Microbiome 2021, 9, 62. [Google Scholar] [CrossRef] [PubMed]
- Meng, C.; Bai, C.; Brown, T.D.; Hood, L.E.; Tian, Q. Human Gut Microbiota and Gastrointestinal Cancer. Genom. Proteom. Bioinform. 2018, 16, 33–49. [Google Scholar] [CrossRef] [PubMed]
- Zhou, C.B.; Zhou, Y.L.; Fang, J.Y. Gut Microbiota in Cancer Immune Response and Immunotherapy. Trends Cancer 2021, 7, 647–660. [Google Scholar] [CrossRef] [PubMed]
- Tsai, Y.L.; Lin, T.L.; Chang, C.J.; Wu, T.R.; Lai, W.F.; Lu, C.C.; Lai, H.C. Probiotics, prebiotics and amelioration of diseases. J. Biomed. Sci. 2019, 26, 3. [Google Scholar] [CrossRef] [PubMed]
- Yadav, M.K.; Kumari, I.; Singh, B.; Sharma, K.K.; Tiwari, S.K. Probiotics, prebiotics and synbiotics: Safe options for next-generation therapeutics. Appl. Microbiol. Biotechnol. 2022, 106, 505–521. [Google Scholar] [CrossRef]
- Ireson, C.R.; Alavijeh, M.S.; Palmer, A.M.; Fowler, E.R.; Jones, H.J. The role of mouse tumour models in the discovery and development of anticancer drugs. Br. J. Cancer 2019, 121, 101–108. [Google Scholar] [CrossRef] [PubMed]
- Lofgren, J.; Miller, A.L.; Lee, C.C.S.; Bradshaw, C.; Flecknell, P.; Roughan, J. Analgesics promote welfare and sustain tumor growth in orthotopic 4T1 and B16 mouse cancer models. Lab. Anim. 2018, 52, 351–364. [Google Scholar] [CrossRef]
- El-Ashmawy, N.E.; Khedr, E.G.; Ebeid, E.M.; Salem, M.L.; Zidan, A.A.; Mosalam, E.M. Enhanced anticancer effect and reduced toxicity of doxorubicin in combination with thymoquinone released from poly-N-acetyl glucosamine nanomatrix in mice bearing solid Ehrlish carcinoma. Eur. J. Pharm. Sci. Off. J. Eur. Fed. Pharm. Sci. 2017, 109, 525–532. [Google Scholar] [CrossRef] [PubMed]
- Ylikahri, R. Metabolic and nutritional aspects of xylitol. Adv. Food Res. 1979, 25, 159–180. [Google Scholar] [CrossRef] [PubMed]
- DeBerardinis, R.J.; Chandel, N.S. We need to talk about the Warburg effect. Nat Metab. 2020, 2, 127–129. [Google Scholar] [CrossRef] [PubMed]
- Cortinovis, C.; Caloni, F. Household Food Items Toxic to Dogs and Cats. Front. Vet. Sci. 2016, 3, 26. [Google Scholar] [CrossRef]
- Marlowe, F.W.; Berbesque, J.C. Tubers as fallback foods and their impact on Hadza hunter-gatherers. Am. J. Phys. Anthropol. 2009, 140, 751–758. [Google Scholar] [CrossRef]
- Marean, C.W. When the sea saved humanity. Sci. Am. 2010, 303, 54–61. [Google Scholar] [CrossRef]
- Iannotti, L.L.; Gyimah, E.A.; Reid, M.; Chapnick, M.; Cartmill, M.K.; Lutter, C.K.; Hilton, C.; Gildner, T.E.; Quinn, E.A. Child dietary patterns in Homo sapiens evolution: A systematic review. Evol. Med. Public Health 2022, 10, 371–390. [Google Scholar] [CrossRef] [PubMed]
- Olm, M.R.; Dahan, D.; Carter, M.M.; Merrill, B.D.; Yu, F.B.; Jain, S.; Meng, X.; Tripathi, S.; Wastyk, H.; Neff, N.; et al. Robust variation in infant gut microbiome assembly across a spectrum of lifestyles. Science 2022, 376, 1220–1223. [Google Scholar] [CrossRef] [PubMed]
- De Filippo, C.; Cavalieri, D.; Di Paola, M.; Ramazzotti, M.; Poullet, J.B.; Massart, S.; Collini, S.; Pieraccini, G.; Lionetti, P. Impact of diet in shaping gut microbiota revealed by a comparative study in children from Europe and rural Africa. Proc. Natl. Acad. Sci. USA 2010, 107, 14691–14696. [Google Scholar] [CrossRef]
- Fan, X.; Liu, H.; Liu, M.; Wang, Y.; Qiu, L.; Cui, Y. Increased utilization of fructose has a positive effect on the development of breast cancer. PeerJ 2017, 5, e3804. [Google Scholar] [CrossRef] [PubMed]
- Ting, K.K.Y. Fructose-induced metabolic reprogramming of cancer cells. Front. Immunol. 2024, 15, 1375461. [Google Scholar] [CrossRef] [PubMed]
- Nakagawa, T.; Lanaspa, M.A.; Millan, I.S.; Fini, M.; Rivard, C.J.; Sanchez-Lozada, L.G.; Andres-Hernando, A.; Tolan, D.R.; Johnson, R.J. Fructose contributes to the Warburg effect for cancer growth. Cancer Metab. 2020, 8, 16. [Google Scholar] [CrossRef]
- Trachootham, D.; Alexandre, J.; Huang, P. Targeting cancer cells by ROS-mediated mechanisms: A radical therapeutic approach? Nature reviews. Drug Discov. 2009, 8, 579–591. [Google Scholar] [CrossRef] [PubMed]
- Niu, B.; Liao, K.; Zhou, Y.; Wen, T.; Quan, G.; Pan, X.; Wu, C. Application of glutathione depletion in cancer therapy: Enhanced ROS-based therapy, ferroptosis, and chemotherapy. Biomaterials 2021, 277, 121110. [Google Scholar] [CrossRef] [PubMed]
- Wen, Y.; Chen, H.; Zhang, L.; Wu, M.; Zhang, F.; Yang, D.; Shen, J.; Chen, J. Glycyrrhetinic acid induces oxidative/nitrative stress and drives ferroptosis through activating NADPH oxidases and iNOS, and depriving glutathione in tri-ple-negative breast cancer cells. Free Radic. Biol. Med. 2021, 173, 41–51. [Google Scholar] [CrossRef]
- Hellstrand, K.; Brune, M.; Naredi, P.; Mellqvist, U.H.; Hansson, M.; Gehlsen, K.R.; Hermodsson, S. Histamine: A novel approach to cancer immunotherapy. Cancer Investig. 2000, 18, 347–355. [Google Scholar] [CrossRef] [PubMed]
- Stoyanov, E.; Uddin, M.; Mankuta, D.; Dubinett, S.M.; Levi-Schaffer, F. Mast cells and histamine enhance the proliferation of non-small cell lung cancer cells. Lung Cancer 2012, 75, 38–44. [Google Scholar] [CrossRef]
- Chakrabarty, R.P.; Chandel, N.S. Mitochondria as Signaling Organelles Control Mammalian Stem Cell Fate. Cell Stem Cell 2021, 28, 394–408. [Google Scholar] [CrossRef] [PubMed]
- Simonsen, T.G.; Gaustad, J.V.; Leinaas, M.N.; Rofstad, E.K. High interstitial fluid pressure is associated with tumor-line specific vascular abnormalities in human melanoma xenografts. PLoS ONE 2012, 7, e40006. [Google Scholar] [CrossRef] [PubMed]
- Shin, G.R.; Kim, H.E.; Kim, J.H.; Choi, S.; Kim, M.S. Advances in Injectable In Situ-Forming Hydrogels for Intra-tumoral Treatment. Pharmaceutics 2021, 13, 1953. [Google Scholar] [CrossRef] [PubMed]
- Gould, H.J., 3rd; Paul, D. Targeted Osmotic Lysis: A Novel Approach to Targeted Cancer Therapies. Biomedicines 2022, 10, 838. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.X.; Liu, W.J.; Zhang, H.R.; Zhang, Z.W. Delivery of bevacizumab by intracranial injection: Assessment in glioma model. OncoTargets Ther. 2018, 11, 2673–2683. [Google Scholar] [CrossRef]
- Algrafi, A.S.; Jamal, A.A.; Ismaeel, D.M. Microbiota as a New Target in Cancer Pathogenesis and Treatment. Cureus 2023, 15, e47072. [Google Scholar] [CrossRef]
- Wu, H.J.; Wu, E. The role of gut microbiota in immune homeostasis and autoimmunity. Gut Microbes 2012, 3, 4–14. [Google Scholar] [CrossRef]
- Halley, A.; Leonetti, A.; Gregori, A.; Tiseo, M.; Deng, D.M.; Giovannetti, E.; Peters, G.J. The Role of the Microbiome in Cancer and Therapy Efficacy: Focus on Lung Cancer. Anticancer Res. 2020, 40, 4807–4818. [Google Scholar] [CrossRef] [PubMed]
- Zuo, Q.L.; Cai, X.; Zheng, X.Y.; Chen, D.S.; Li, M.; Liu, Z.Q.; Chen, K.Q.; Han, F.F.; Zhu, X. Influences of Xylitol Consumption at Different Dosages on Intestinal Tissues and Gut Microbiota in Rats. J. Agric. Food Chem. 2021, 69, 12002–12011. [Google Scholar] [CrossRef]
- Koppel, N.; Maini Rekdal, V.; Balskus, E.P. Chemical transformation of xenobiotics by the human gut microbiota. Science 2017, 356, eaag2770. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mouse Strain | Cancer Cell Line | Immune System | Mice |
|---|---|---|---|
| BALB/C | 4T1 mammary carcinoma | Adaptive immunity | 20–10 controls and 10 experimental |
| C57BL/6 | B16F10 melanoma | Innate immunity | 20–10 controls and 10 experimental |
| Metabolite | Fold Change | log2 (FC) | p Value |
|---|---|---|---|
| L-tyrosine methyl ester | 0.13098 | −2.9326 | 0.00055 |
| tryptamine | 0.43146 | −1.2127 | 0.006415 |
| 2,3-bisphospho-D-glycerate | 0.34972 | −1.5157 | 0.017592 |
| ATP/dGTP | 0.29392 | −1.7665 | 0.019468 |
| cysteine | 0.54949 | −0.86384 | 0.02808 |
| choline+ | 0.59869 | −0.74012 | 0.032148 |
| GDP | 0.58635 | −0.77016 | 0.03544 |
| 2-/3-phosphoglycerate | 0.30686 | −1.7044 | 0.037869 |
| glutathione | 0.42802 | −1.2242 | 0.041884 |
| GAP/DHAP | 0.29889 | −1.7423 | 0.041934 |
| D-erythrose-4-phosphate | 0.25721 | −1.959 | 0.043739 |
| CDP | 0.34568 | −1.5325 | 0.047149 |
| FAD | 0.3264 | −1.6153 | 0.048366 |
| putrescine | 0.28675 | −1.8022 | 0.048865 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cannon, M.; Cosantino, A.; Tran, L.; Chandel, N.S.; Ghoreishi, N. Effects of Xylitol on Tumor Progression in Syngeneic Mice Cancer Models. Nutraceuticals 2025, 5, 4. https://doi.org/10.3390/nutraceuticals5010004
Cannon M, Cosantino A, Tran L, Chandel NS, Ghoreishi N. Effects of Xylitol on Tumor Progression in Syngeneic Mice Cancer Models. Nutraceuticals. 2025; 5(1):4. https://doi.org/10.3390/nutraceuticals5010004
Chicago/Turabian StyleCannon, Mark, Ashlee Cosantino, Lori Tran, Navdeep S. Chandel, and Nayereh Ghoreishi. 2025. "Effects of Xylitol on Tumor Progression in Syngeneic Mice Cancer Models" Nutraceuticals 5, no. 1: 4. https://doi.org/10.3390/nutraceuticals5010004
APA StyleCannon, M., Cosantino, A., Tran, L., Chandel, N. S., & Ghoreishi, N. (2025). Effects of Xylitol on Tumor Progression in Syngeneic Mice Cancer Models. Nutraceuticals, 5(1), 4. https://doi.org/10.3390/nutraceuticals5010004