Epithelial Membrane Protein-3 and Chitinase-3-like Protein-1 as New Prognostic Predictors of Glioma, a Two-Gene Study

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. GEPIA Database
2.2. CGGA Database
2.3. cBioPortal
2.4. LinkedOmics
2.5. RNA Extraction and Real-Time Quantitative PCR (qRT-PCR)
2.6. Samples
2.7. Statistical Analysis
2.8. Immunohistochemistry (IHC)
3. Results
3.1. The Expression Levels of EMP3 and CHI3L1 in Glioma Were Higher Than in Normal Brain Tissues
3.2. Clinical Characteristics of Glioma Patients
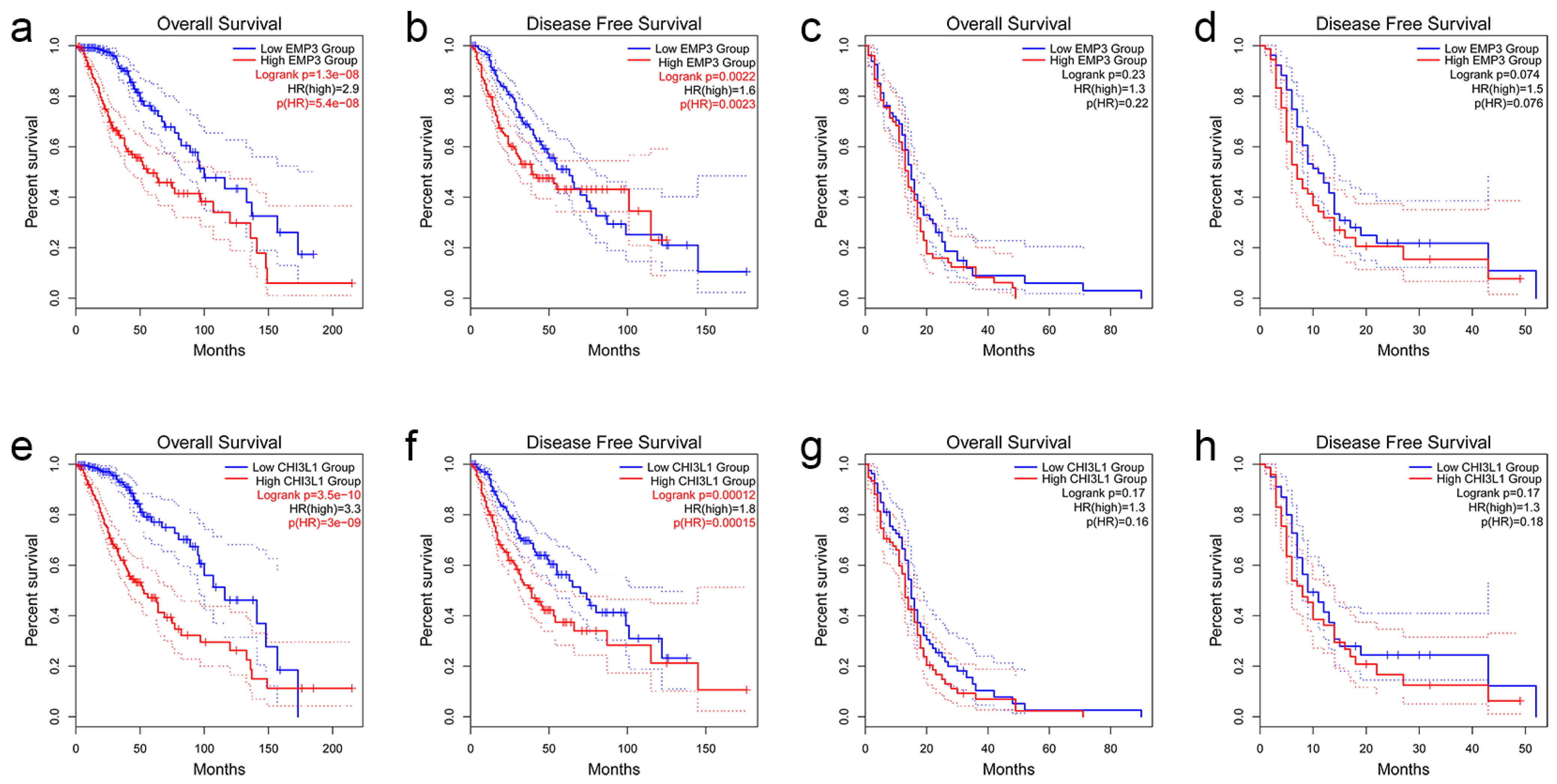
3.3. The Prognostic Significance of the Expression Levels of EMP3 and CHI3L1 in Glioma
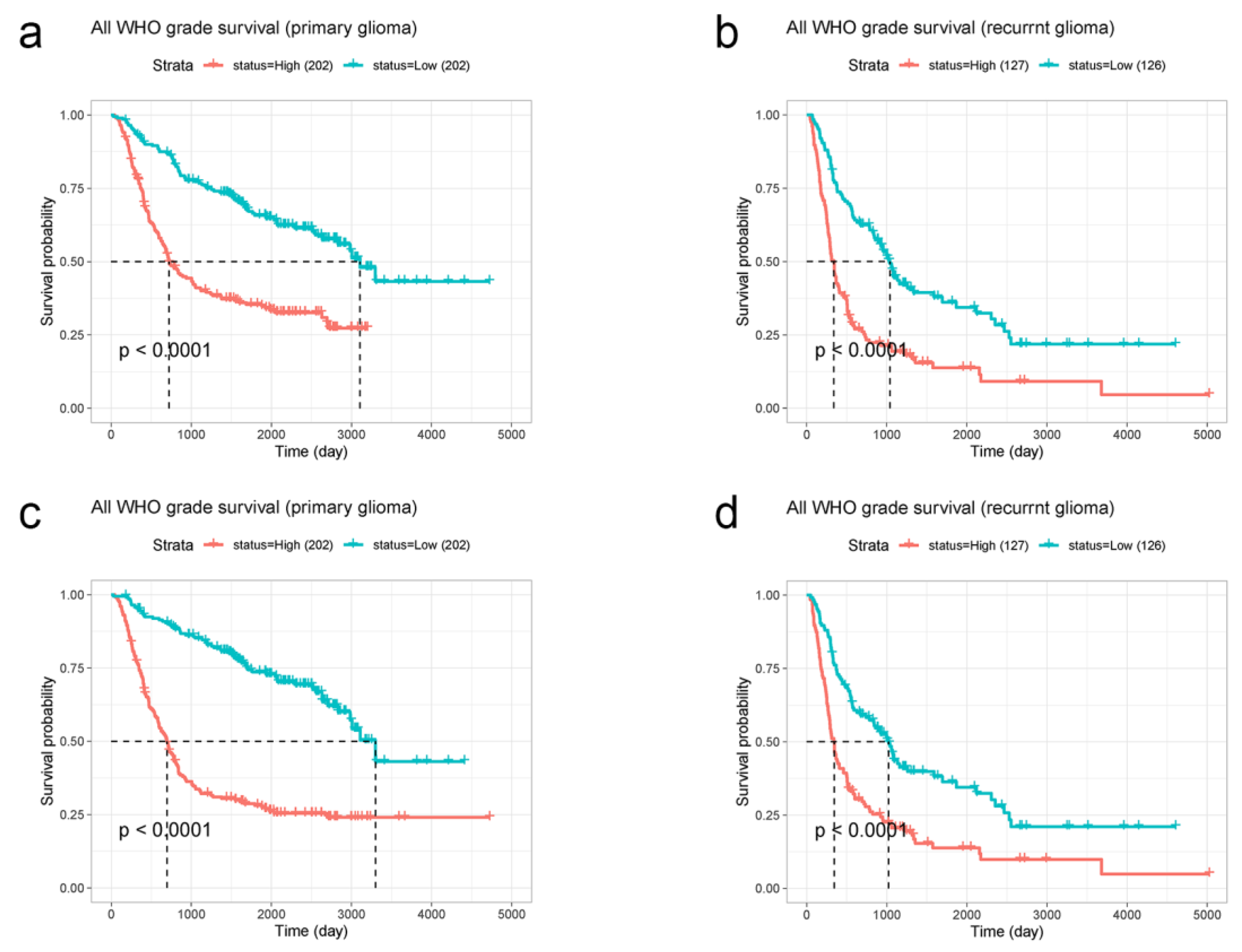
3.4. Prognostic Significance of EMP3 and CHI3L1 in Primary and Recurrent Gliomas of CGGA
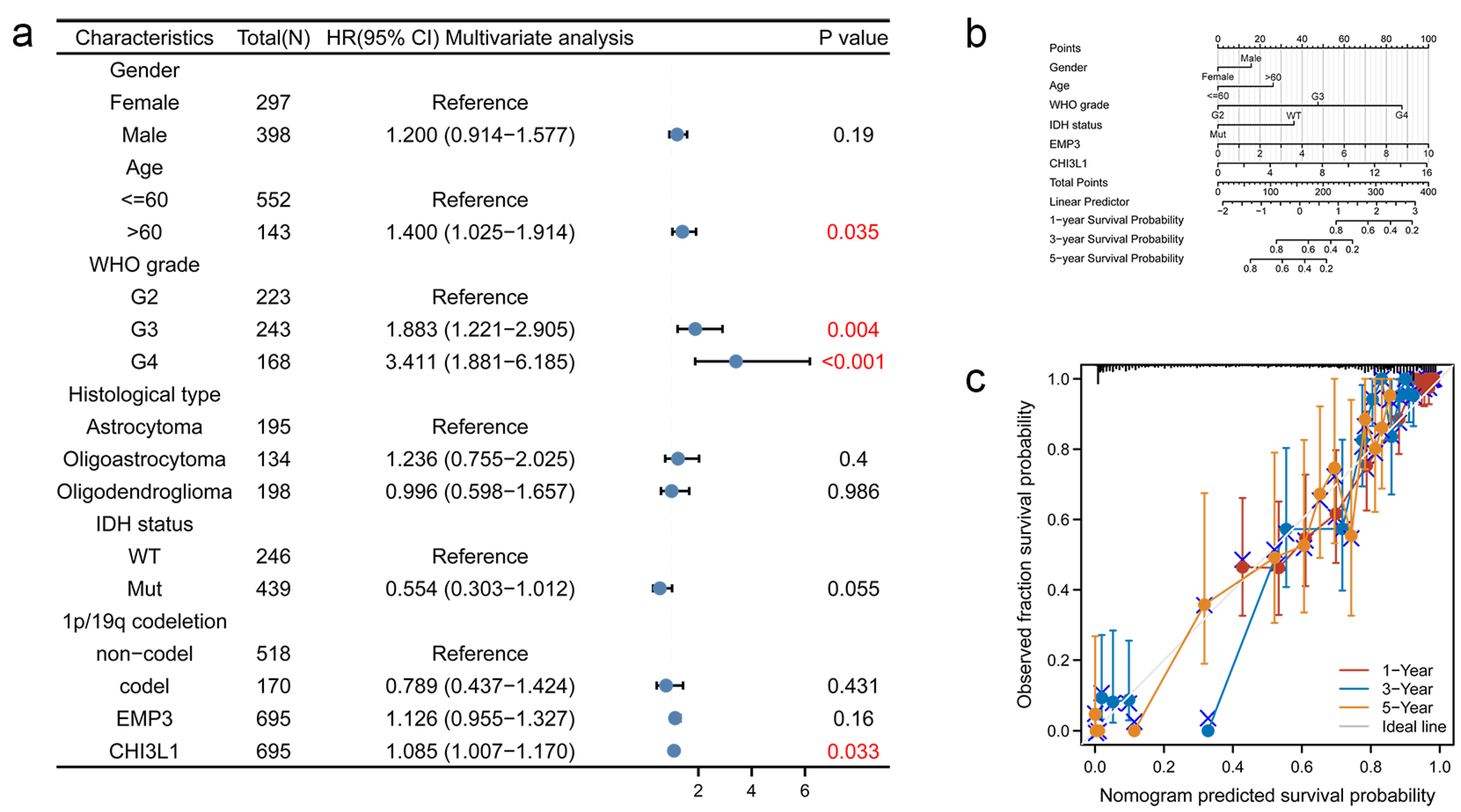
3.5. COX Regression Analysis of Overall Survival of Glioma Patients
3.6. Gene Changes and Prognosis of EMP3 and CHI3L1 in Glioma
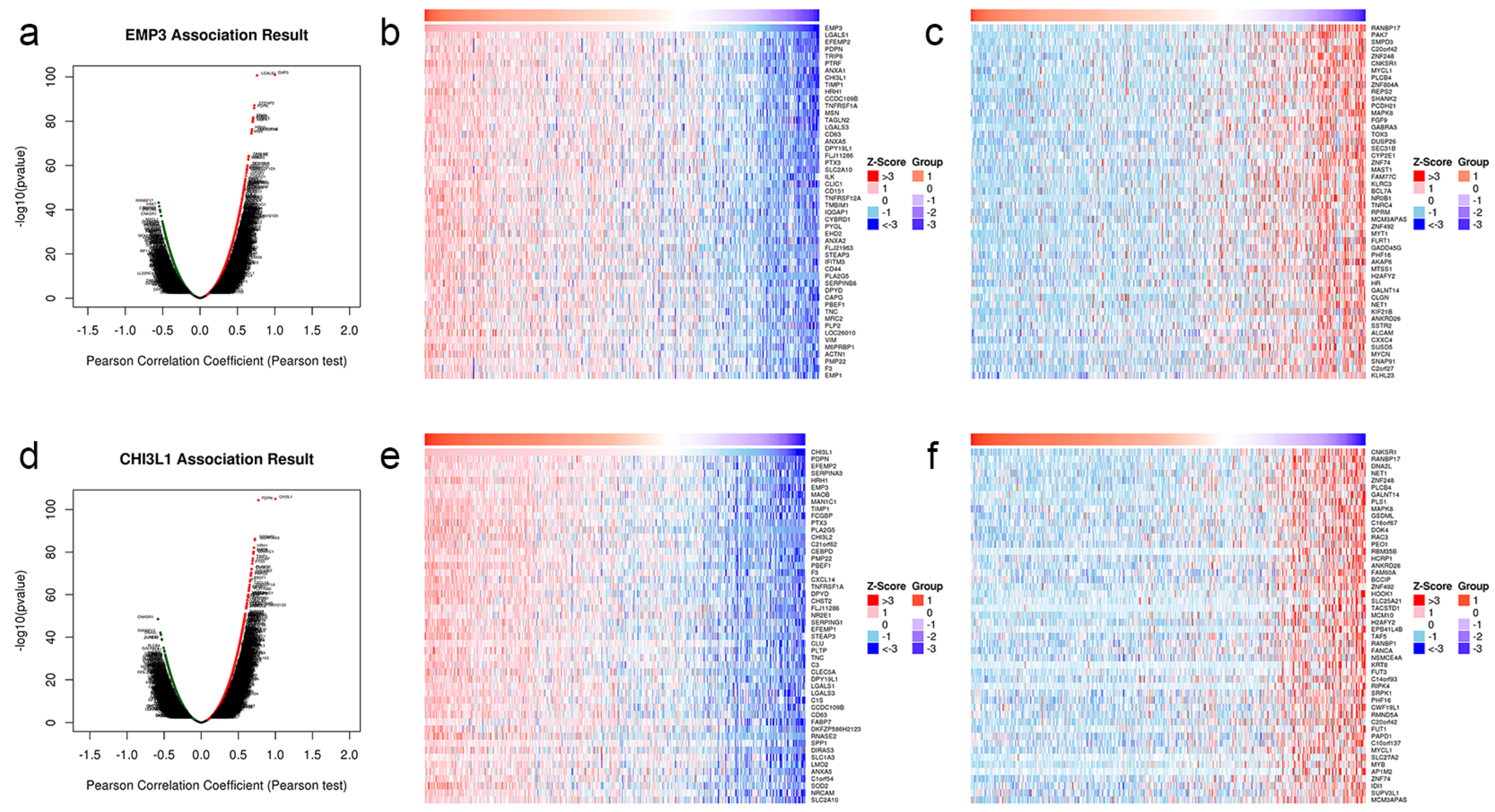
3.7. There Is Correlation between EMP3 and CHI3L1
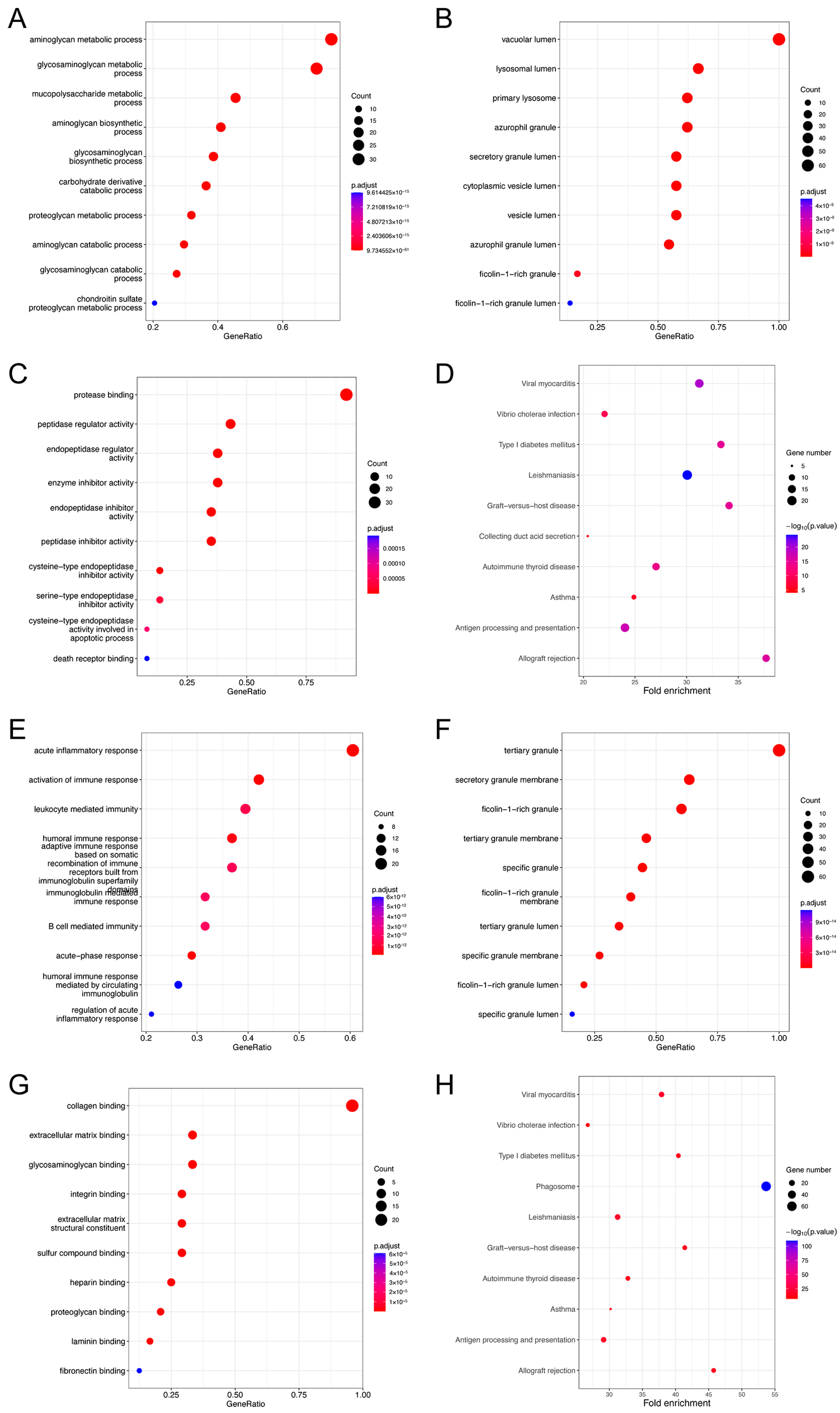
3.8. GO and KEGG Pathway Enrichment Analysis of EMP3 and CHI3L1 Co-Expression Genes in Glioma
3.9. The Expressions of CHI3L1 and EMP3 Were Low in Glioma Cells and Decreased with an Increase in Glioma Grade
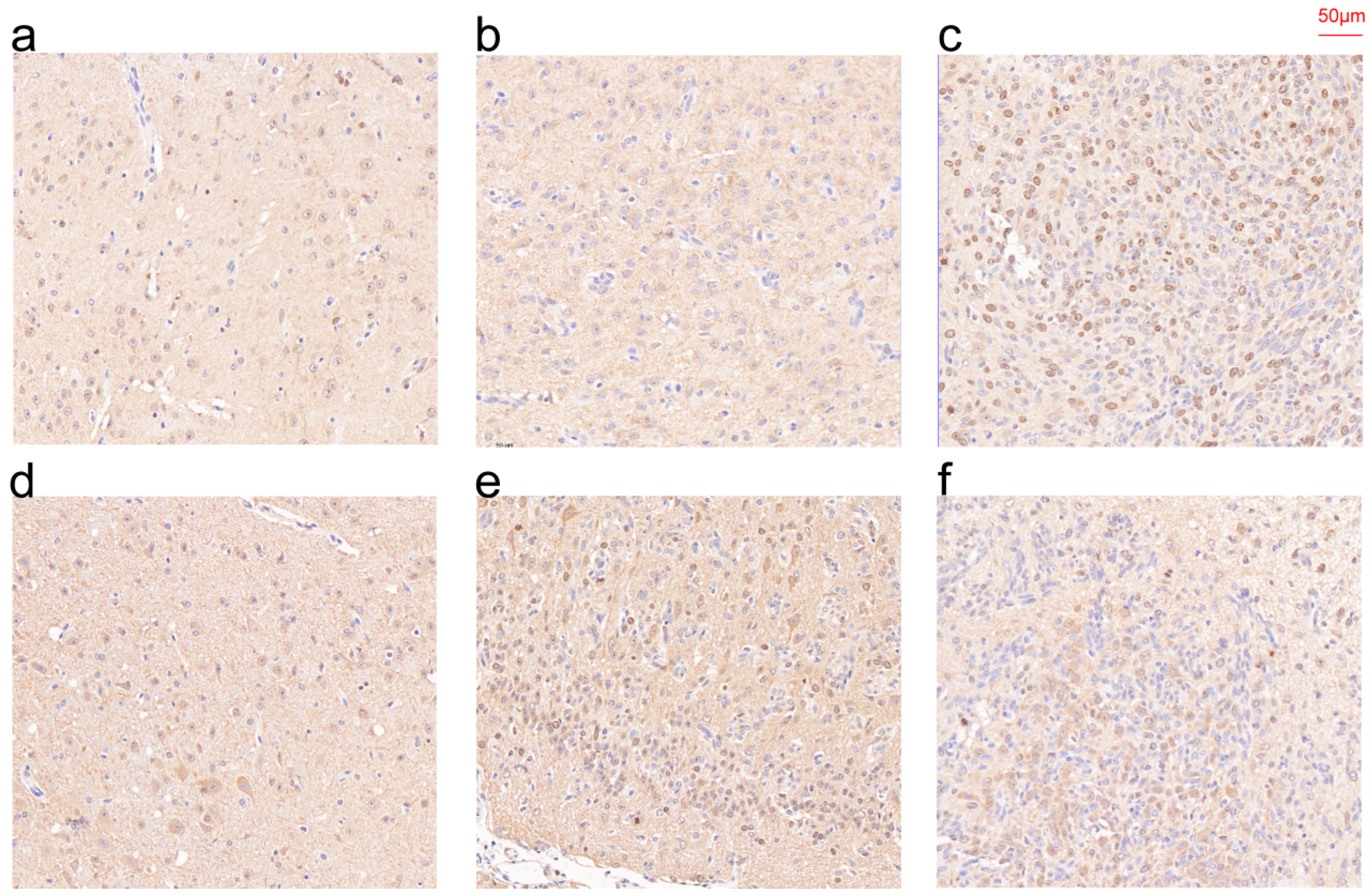
3.10. IHC Results of CHI3L1 and EMP3 in Patient Specimens
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Xu, D.; Liu, R.; Meng, L.; Zhang, Y.; Lu, G.; Ma, P. Long non-coding RNA ENST01108 promotes carcinogenesis of glioma by acting as a molecular sponge to modulate miR-489. Biomed. Pharmacother. 2018, 100, 20–28. [Google Scholar] [CrossRef]
- Ostrom, Q.T.; Gittleman, H.; Stetson, L.; Virk, S.M.; Barnholtz-Sloan, J.S. Epidemiology of gliomas. Cancer Treat. Res. 2015, 163, 1–14. Available online: https://pubmed.ncbi.nlm.nih.gov/25468222/ (accessed on 1 November 2022).
- Ostrom, Q.T.; Gittleman, H.; Truitt, G.; Boscia, A.; Kruchko, C.; Barnholtz-Sloan, J.S. CBTRUS Statistical Report: Primary Brain and Other Central Nervous System Tumors Diagnosed in the United States in 2011–2015. Neuro-Oncology 2018, 20, iv1–iv86. [Google Scholar] [CrossRef]
- Weller, M.; van den Bent, M.; Preusser, M.; Le Rhun, E.; Tonn, J.C.; Minniti, G.; Bendszus, M.; Balana, C.; Chinot, O.; Dirven, L.; et al. EANO guidelines on the diagnosis and treatment of diffuse gliomas of adulthood. Nat. Rev. Clin. Oncol. 2020, 18, 170–186. [Google Scholar] [CrossRef]
- Louis, D.N.; Perry, A.; Reifenberger, G.; Von Deimling, A.; Figarella-Branger, D.; Cavenee, W.K.; Ohgaki, H.; Wiestler, O.D.; Kleihues, P.; Ellison, D.W. The 2016 World Health Organization Classification of Tumors of the Central Nervous System: A summary. Acta Neuropathol. 2016, 131, 803–820. [Google Scholar] [CrossRef]
- Li, B.; Meng, C.; Zhang, X.; Cong, D.; Gao, X.; Gao, W.; Ju, D.; Hu, S. Effect of photodynamic therapy combined with torasemide on the expression of matrix metalloproteinase 2 and sodium-potassium-chloride cotransporter 1 in rat peritumoral edema and glioma. Oncol. Lett. 2016, 11, 2084–2090. [Google Scholar] [CrossRef]
- Gao, L. Glioma an overview of current classifications characteristics molecular biology and target therapies. Front. Biosci. 2015, 20, 1104–1115. [Google Scholar] [CrossRef]
- Zhang, A.; Xu, H.; Zhang, Z.; Liu, Y.; Han, X.; Yuan, L.; Ni, Y.; Gao, S.; Xu, Y.; Chen, S.; et al. Establishment of a nomogram with EMP3 for predicting clinical outcomes in patients with glioma: A bi-center study. CNS Neurosci. Ther. 2021, 27, 1238–1250. [Google Scholar] [CrossRef]
- Gerber, N.K.; Goenka, A.; Turcan, S.; Reyngold, M.; Makarov, V.; Kannan, K.; Beal, K.; Omuro, A.; Yamada, Y.; Gutin, P.; et al. Transcriptional diversity of long-term glioblastoma survivors. Neuro-Oncology 2014, 16, 1186–1195. [Google Scholar] [CrossRef]
- Jovčevska, I. Genetic secrets of long-term glioblastoma survivors. Bosn. J. Basic Med. Sci. 2018, 19, 116–124. [Google Scholar] [CrossRef]
- Wang, Y.-W.; Cheng, H.-L.; Ding, Y.-R.; Chou, L.-H.; Chow, N.-H. EMP1, EMP 2, and EMP3 as novel therapeutic targets in human cancer. Biochim. Et Biophys. Acta Rev. Cancer 2017, 1868, 199–211. Available online: https://pubmed.ncbi.nlm.nih.gov/28408326/ (accessed on 17 August 2022). [CrossRef]
- Ben-Porath, I.; Kozak, C.A.; Benvenisty, N. Chromosomal Mapping ofTmp(Emp1),Xmp(Emp2), andYmp(Emp3), Genes Encoding Membrane Proteins Related toPmp22. Genomics 1998, 49, 443–447. [Google Scholar] [CrossRef]
- Christians, A.; Poisel, E.; Hartmann, C.; Deimling, A.; Pusch, S. Characterization of the epithelial membrane protein 3 interaction network reveals a potential functional link to mitogenic signal transduction regulation. Int. J. Cancer 2019, 145, 461–473. [Google Scholar] [CrossRef]
- Taylor, V.; Suter, U. Epithelial membrane protein-2 and epithelial membrane protein-3: Two novel members of the peripheral myelin protein 22 gene family. Gene 1996, 175, 115–120. [Google Scholar] [CrossRef]
- Thul, P.J.; Åkesson, L.; Wiking, M.; Mahdessian, D.; Geladaki, A.; Ait Blal, H.; Alm, T.; Asplund, A.; Björk, L.; Breckels, L.M.; et al. A subcellular map of the human proteome. Science 2017, 356, eaal3321. [Google Scholar] [CrossRef]
- Hsieh, Y.-H.; Hsieh, S.-C.; Lee, C.-H.; Yang, S.-F.; Cheng, C.-W.; Tang, M.-J.; Lin, C.-L.; Lin, C.-L.; Chou, R.-H. Targeting EMP3 suppresses proliferation and invasion of hepatocellular carcinoma cells through inactivation of PI3K/Akt pathway. Oncotarget 2015, 6, 34859–34874. [Google Scholar] [CrossRef]
- Wang, Y.-W.; Li, W.-M.; Wu, W.-J.; Chai, C.-Y.; Liu, H.-S.; Lai, M.-D.; Chow, N.-H. Potential Significance of EMP3 in Patients with Upper Urinary Tract Urothelial Carcinoma: Crosstalk with ErbB2-PI3K-Akt Pathway. J. Urol. 2014, 192, 242–251. [Google Scholar] [CrossRef]
- Uhlén, M.; Fagerberg, L.; Hallström, B.M.; Lindskog, C.; Oksvold, P.; Mardinoglu, A.; Sivertsson, Å.; Kampf, C.; Sjöstedt, E.; Asplund, A.; et al. Tissue-based map of the human proteome. Science 2015, 347, 1260419. [Google Scholar] [CrossRef]
- Kanton, S.; Boyle, M.J.; He, Z.; Santel, M.; Weigert, A.; Sanchís-Calleja, F.; Guijarro, P.; Sidow, L.; Fleck, J.S.; Han, D.; et al. Organoid single-cell genomic atlas uncovers human-specific features of brain development. Nature 2019, 574, 418–422. [Google Scholar] [CrossRef]
- Scrideli, C.A.; Carlotti, C.G.; Okamoto, O.K.; Andrade, V.S.; Cortez, M.A.A.; Motta, F.J.N.; Lucio-Eterovic, A.K.; Neder, L.; Rosemberg, S.; Oba-Shinjo, S.M.; et al. Gene expression profile analysis of primary glioblastomas and non-neoplastic brain tissue: Identification of potential target genes by oligonucleotide microarray and real-time quantitative PCR. J. Neuro-Oncol. 2008, 88, 281–291. [Google Scholar] [CrossRef]
- Bolin, L.M.; McNeil, T.; Lucian, L.A.; DeVaux, B.; Franz-Bacon, K.; Gorman, D.M.; Zurawski, S.; Murray, R.; McClanahan, T.K. HNMP-1: A Novel Hematopoietic and Neural Membrane Protein Differentially Regulated in Neural Development and Injury. J. Neurosci. 1997, 17, 5493–5502. [Google Scholar] [CrossRef]
- Jun, F.; Hong, J.; Liu, Q.; Guo, Y.; Liao, Y.; Huang, J.; Wen, S.; Shen, L. Epithelial membrane protein 3 regulates TGF-β signaling activation in CD44-high glioblastoma. Oncotarget 2016, 8, 14343–14358. [Google Scholar] [CrossRef]
- Yue, H.; Xu, Q.; Xie, S. High EMP3 expression might independently predict poor overall survival in glioblastoma and its expression is related to DNA methylation. Medicine 2018, 97, e9538. [Google Scholar] [CrossRef]
- Zhao, T.; Su, Z.; Li, Y.; Zhang, X.; You, Q. Chitinase-3 like-protein-1 function and its role in diseases. Signal Transduct. Target. Ther. 2020, 5, 201. Available online: https://www.nature.com/articles/s41392-020-00303-7 (accessed on 1 November 2022). [CrossRef]
- Pelloski, C.E. YKL-40 Expression is Associated with Poorer Response to Radiation and Shorter Overall Survival in Glioblastoma. Clin. Cancer Res. 2005, 11, 3326–3334. [Google Scholar] [CrossRef]
- Junker, N.; Johansen, J.S.; Hansen, L.T.; Lund, E.L.; Kristjansen, P.E.G. Regulation of YKL-40 expression during genotoxic or microenvironmental stress in human glioblastoma cells. Cancer Sci. 2005, 96, 183–190. [Google Scholar] [CrossRef]
- Kavsan, V.M.; Baklaushev, V.P.; Balynska, O.V.; Iershov, A.V. Gene Encoding Chitinase 3-Like 1 Protein (CHI3L1) is a Putative Oncogene. Int. J. Biol. Sci. 2011, 7, 230–237. [Google Scholar] [CrossRef]
- Tang, Z.; Li, C.; Kang, B.; Gao, G.; Li, C.; Zhang, Z. GEPIA: A web server for cancer and normal gene expression profiling and interactive analyses. Nucleic Acids Res. 2017, 45, W98–W102. [Google Scholar] [CrossRef]
- Zhao, Z.; Zhang, K.-N.; Wang, Q.; Li, G.; Zeng, F.; Zhang, Y.; Wu, F.; Chai, R.; Wang, Z.; Zhang, C.; et al. Chinese Glioma Genome Atlas (CGGA): A Comprehensive Resource with Functional Genomic Data from Chinese Glioma Patients. Genom. Proteom. Bioinform. 2021, 19, 1–12. [Google Scholar] [CrossRef]
- Gao, J.; Aksoy, B.A.; Dogrusoz, U.; Dresdner, G.; Gross, B.E.; Sumer, S.O.; Sun, Y.; Jacobsen, A.; Sinha, R.; Larsson, E.; et al. Integrative Analysis of Complex Cancer Genomics and Clinical Profiles Using the cBioPortal. Sci. Signal. 2013, 6, pl1. [Google Scholar] [CrossRef]
- Cerami, E.; Gao, J.; Dogrusoz, U.; Gross, B.E.; Sumer, S.O.; Aksoy, B.A.; Jacobsen, A.; Byrne, C.J.; Heuer, M.L.; Larsson, E.; et al. The cBio Cancer Genomics Portal: An Open Platform for Exploring Multidimensional Cancer Genomics Data: Figure 1. Cancer Discov. 2012, 2, 401–404. [Google Scholar] [CrossRef]
- Vasaikar, S.V.; Straub, P.; Wang, J.; Zhang, B. LinkedOmics: Analyzing multi-omics data within and across 32 cancer types. Nucleic Acids Res. 2017, 46, D956–D963. [Google Scholar] [CrossRef]
- Lange, A.M.; Lo, H.-W. Inhibiting TRK Proteins in Clinical Cancer Therapy. Cancers 2018, 10, 105. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC5923360/ (accessed on 1 November 2022). [CrossRef]
- Gao, Y.-F.; Zhu, T.; Mao, C.-X.; Liu, Z.-X.; Wang, Z.-B.; Mao, X.-Y.; Li, L.; Yin, J.-Y.; Zhou, H.-H.; Liu, Z.-Q. PPIC, EMP3 and CHI3L1 Are Novel Prognostic Markers for High Grade Glioma. Int. J. Mol. Sci. 2016, 17, 1808. [Google Scholar] [CrossRef]
- Ku, B.M.; Lee, Y.K.; Ryu, J.; Jeong, J.Y.; Choi, J.; Eun, K.M.; Shin, H.Y.; Kim, D.G.; Hwang, E.M.; Yoo, J.C.; et al. CHI3L1 (YKL-40) is expressed in human gliomas and regulates the invasion, growth and survival of glioma cells. Int. J. Cancer 2011, 128, 1316–1326. [Google Scholar] [CrossRef]
- Ernst, A.; Hofmann, S.; Ahmadi, R.; Becker, N.; Korshunov, A.; Engel, F.; Hartmann, C.; Felsberg, J.; Sabel, M.; Peterziel, H.; et al. Genomic and Expression Profiling of Glioblastoma Stem Cell-like Spheroid Cultures Identifies Novel Tumor-Relevant Genes Associated with Survival. Clin. Cancer Res. 2009, 15, 6541–6550. [Google Scholar] [CrossRef]
- Deluche, E.; Bessette, B.; Durand, S.; Caire, F.; Rigau, V.; Robert, S.; Chaunavel, A.; Forestier, L.; Labrousse, F.; Jauberteau, M.-O.; et al. CHI3L1, NTRK2, 1p/19q and IDH Status Predicts Prognosis in Glioma. Cancers 2019, 11, 544. [Google Scholar] [CrossRef]
- Qin, G.; Li, X.; Chen, Z.; Liao, G.; Su, Y.; Chen, Y.; Zhang, W. Prognostic Value of YKL-40 in Patients with Glioblastoma: A Systematic Review and Meta-analysis. Mol. Neurobiol. 2016, 54, 3264–3270. [Google Scholar] [CrossRef]
- Chen, Q.; Jin, J.; Huang, X.; Wu, F.; Huang, H.; Zhan, R. EMP3 mediates glioblastoma-associated macrophage infiltration to drive T cell exclusion. J. Exp. Clin. Cancer Res. 2021, 40, 160. [Google Scholar] [CrossRef]
- Heiland, D.H.; Ferrarese, R.; Claus, R.; Dai, F.; Masilamani, A.P.; Kling, E.; Weyerbrock, A.; Kling, T.; Nelander, S.; Carro, M.S. c-Jun-N-terminal phosphorylation regulates DNMT1 expression and genome wide methylation in gliomas. Oncotarget 2016, 8, 6940–6954. [Google Scholar] [CrossRef]
- Turcan, S.; Rohle, D.; Goenka, A.; Walsh, L.A.; Fang, F.; Yilmaz, E.; Campos, C.; Fabius, A.W.M.; Lu, C.; Ward, P.S.; et al. IDH1 mutation is sufficient to establish the glioma hypermethylator phenotype. Nature 2012, 483, 479–483. Available online: https://www.ncbi.nlm.nih.gov/pubmed/22343889/ (accessed on 1 November 2022). [CrossRef]
- Ponta, H.; Sherman, L.; Herrlich, P.A. CD44: From adhesion molecules to signalling regulators. Nat. Rev. Mol. Cell Biol. 2003, 4, 33–45. [Google Scholar] [CrossRef]
- Martija, A.A.; Pusch, S. The Multifunctional Role of EMP3 in the Regulation of Membrane Receptors Associated with IDH-Wild-Type Glioblastoma. Int. J. Mol. Sci. 2021, 22, 5261. [Google Scholar] [CrossRef]
- Thornton, N.; Crew, V.K.; Tilley, L.; Green, C.A.; Tay, C.L.; Griffiths, R.E.; Singleton, B.K.; Spring, F.; Walser, P.; Alattar, A.G.; et al. Disruption of the tumour-associated EMP3 enhances erythroid proliferation and causes the MAM-negative phenotype. Nat. Commun. 2020, 11, 3569. [Google Scholar] [CrossRef]
- He, C.H.; Lee, C.G.; Cruz, C.S.D.; Lee, C.M.; Zhou, Y.; Ahangari, F.; Ma, B.; Herzog, E.L.; Rosenberg, S.A.; Li, Y.; et al. Chitinase 3-like 1 Regulates Cellular and Tissue Responses via IL-13 Receptor α2. Cell Rep. 2013, 4, 830–841. [Google Scholar] [CrossRef]
- Ichimura, K.; Pearson, D.M.; Kocialkowski, S.; Bäcklund, L.M.; Chan, R.; Jones, D.T.; Collins, V.P. IDH1 mutations are present in the majority of common adult gliomas but rare in primary glioblastomas. Neuro-Oncology 2009, 11, 341–347. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | Primers (5′-3′) |
|---|---|
| EMP3-forward | AAGATCAGTACCTCTCAGATGG |
| EMP3-reverse | GCAGCACAAGAGACGTATCATA |
| CHI3L1-forward | CTGTGGGGATAGTGAGGCAT |
| CHI3L1-reverse | CTTGCCAAAATGGTGTCCTT |
| Human-GAPDH-forward | GGAAGCTTGTCATCAATGGAAATC |
| Human-GAPDH-reverse | TGATGACCCTTTTGGCTCCC |
| Case (N) | Expression of CHI3L1 | p Value | EMP3 Expression | p Value | ||
|---|---|---|---|---|---|---|
| Age | <45 | 383 | 351.93 ± 56.01 | <0.001 | 53.98 ± 4.48 | <0.001 |
| >=45 | 310 | 1242.16 ± 167.98 | 122.92 ± 10.46 | |||
| Gender | Male | 398 | 768.95 ± 114.63 | <0.001 | 92.49 ± 7.97 | <0.001 |
| Female | 295 | 724.80 ± 118.66 | 74.47 ± 6.89 | |||
| Histology | LGG | 444 | 247.71 ± 56.08 | <0.001 | 39.12 ± 3.41 | <0.001 |
| GBM | 249 | 1646.08 ± 195.80 | 166.32 ± 12.31 | |||
| WHO | II | 188 | 170.43 ± 43.25 | <0.001 | 36.31 ± 5.75 | <0.001 |
| III | 256 | 304.46 ± 91.85 | 41.18 ± 4.14 | |||
| IV | 249 | 1646.08 ± 195.80 | 166.32 ± 12.31 | |||
| Tumor type | Primary | 422 | 596.04 ± 90.95 | <0.001 | 80.28 ± 7.73 | <0.001 |
| Recurrence | 271 | 990.14 ± 156.96 | 91.89 ± 6.99 | |||
| IDH-1 status | Mutant | 356 | 157.87 ± 53.79 | <0.001 | 25.24 ± 2.56 | <0.001 |
| Wild-type | 286 | 1588.96 ± 177.28 | 154.13 ± 9.85 | |||
| 1p19q status | Codeletion | 145 | 66.72 ± 29.58 | <0.05 | 23.93 ± 5.34 | <0.001 |
| Non-codeletion | 478 | 1049.18 ± 117.30 | 114.40 ± 7.33 | |||
| MGMT status | Methylation | 315 | 608.28 ± 93.16 | <0.001 | 84.80 ± 8.98 | <0.001 |
| Non-methylation | 227 | 894.40 ± 154.03 | 102.02 ± 9.66 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shen, K.; Zhu, J.; Zhou, S.; Jin, X.; Zhai, W.; Sun, L.; Wu, J.; Yu, Z. Epithelial Membrane Protein-3 and Chitinase-3-like Protein-1 as New Prognostic Predictors of Glioma, a Two-Gene Study. Curr. Oncol. 2023, 30, 8686-8702. https://doi.org/10.3390/curroncol30100629
Shen K, Zhu J, Zhou S, Jin X, Zhai W, Sun L, Wu J, Yu Z. Epithelial Membrane Protein-3 and Chitinase-3-like Protein-1 as New Prognostic Predictors of Glioma, a Two-Gene Study. Current Oncology. 2023; 30(10):8686-8702. https://doi.org/10.3390/curroncol30100629
Chicago/Turabian StyleShen, Kecheng, Jiandong Zhu, Shijie Zhou, Xin Jin, Weiwei Zhai, Liang Sun, Jiang Wu, and Zhengquan Yu. 2023. "Epithelial Membrane Protein-3 and Chitinase-3-like Protein-1 as New Prognostic Predictors of Glioma, a Two-Gene Study" Current Oncology 30, no. 10: 8686-8702. https://doi.org/10.3390/curroncol30100629
APA StyleShen, K., Zhu, J., Zhou, S., Jin, X., Zhai, W., Sun, L., Wu, J., & Yu, Z. (2023). Epithelial Membrane Protein-3 and Chitinase-3-like Protein-1 as New Prognostic Predictors of Glioma, a Two-Gene Study. Current Oncology, 30(10), 8686-8702. https://doi.org/10.3390/curroncol30100629