Gal-3 Protein Expression and Localization in Prostate Tumours

 , , ,
, , ,  ,
,  , and
, and 
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Patients and Samples
2.2. Immunohistochemical Evaluation of Galectin-3 Expression and Localization in Normal and Tumour Prostate Tissue (In Vivo Analysis)
2.3. Exploration of Galectin-3 Expression in PCa and Its Impact on Patient Survival in Public Cancer Databases (In Silico Analysis)
2.4. Statistical Analysis
3. Results
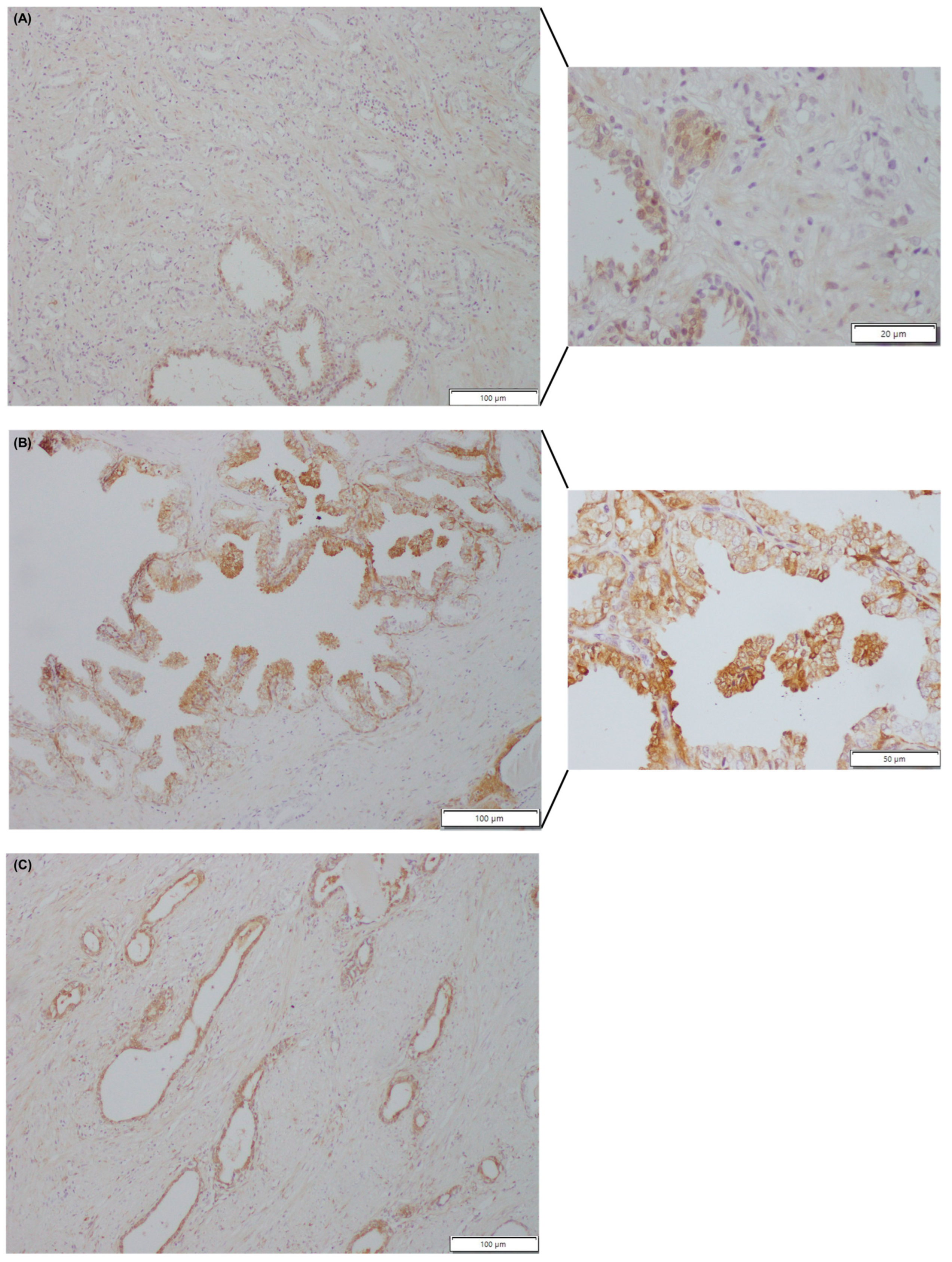
3.1. Immunohistochemical Evaluation of Galectin-3 Expression and Localization in Normal and Tumour Prostate Tissue (In Vivo Analysis)
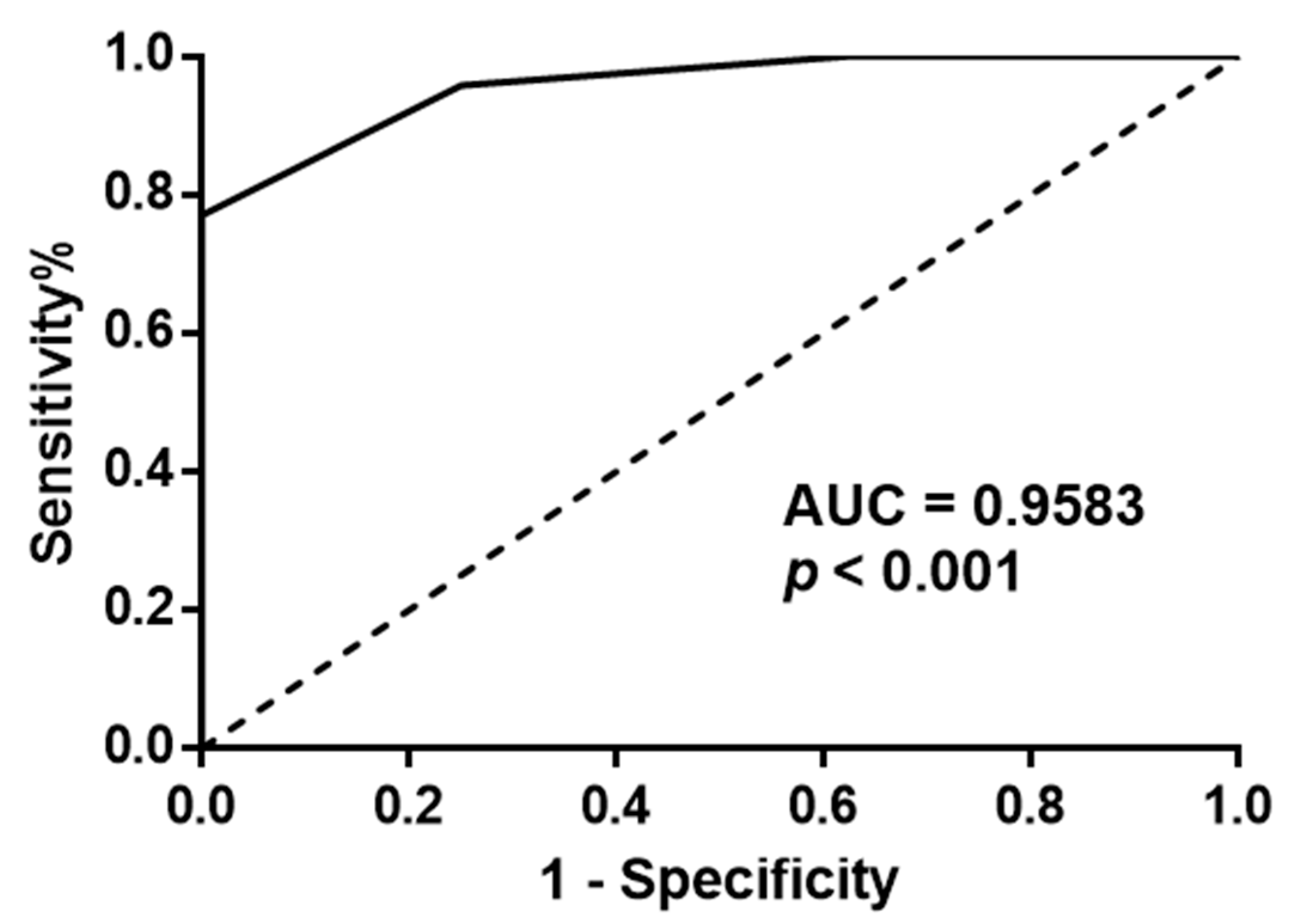
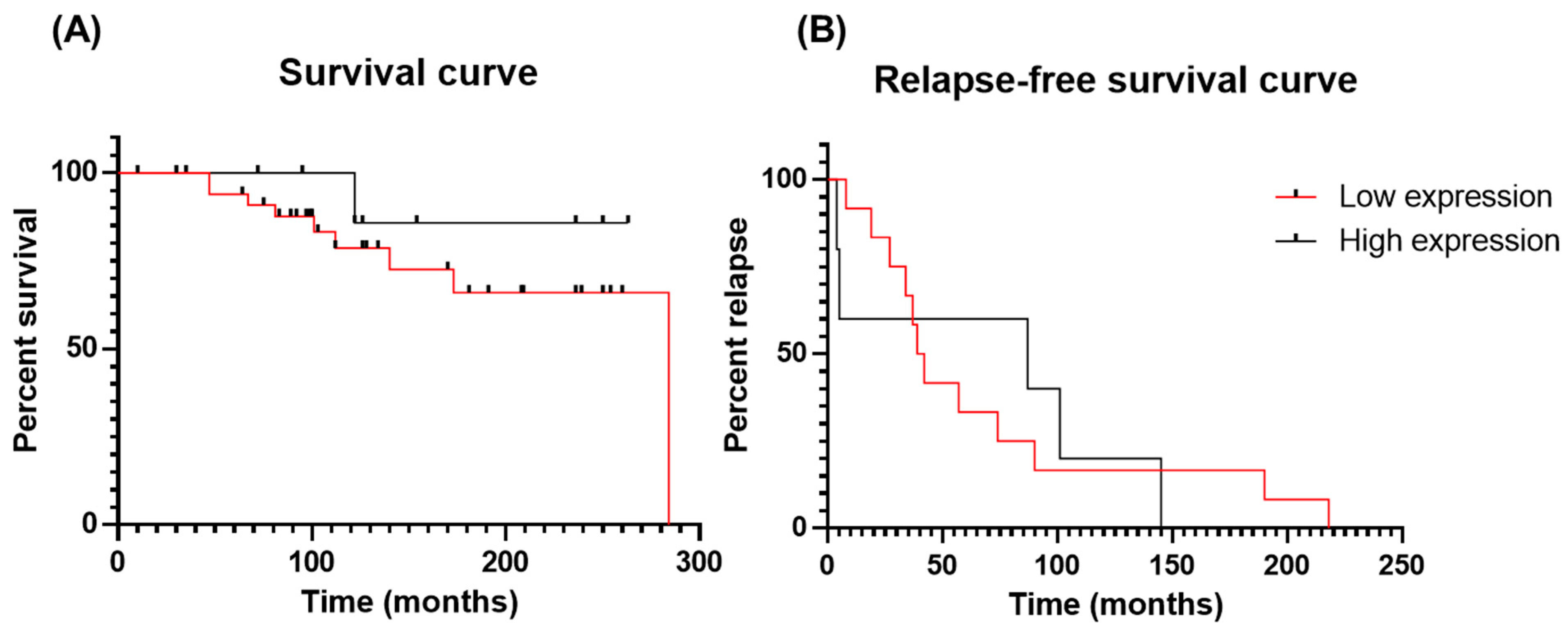
3.2. Impact of Galectin-3 Expression on PCa Patient Survival and Relapse (In Vivo Analysis)
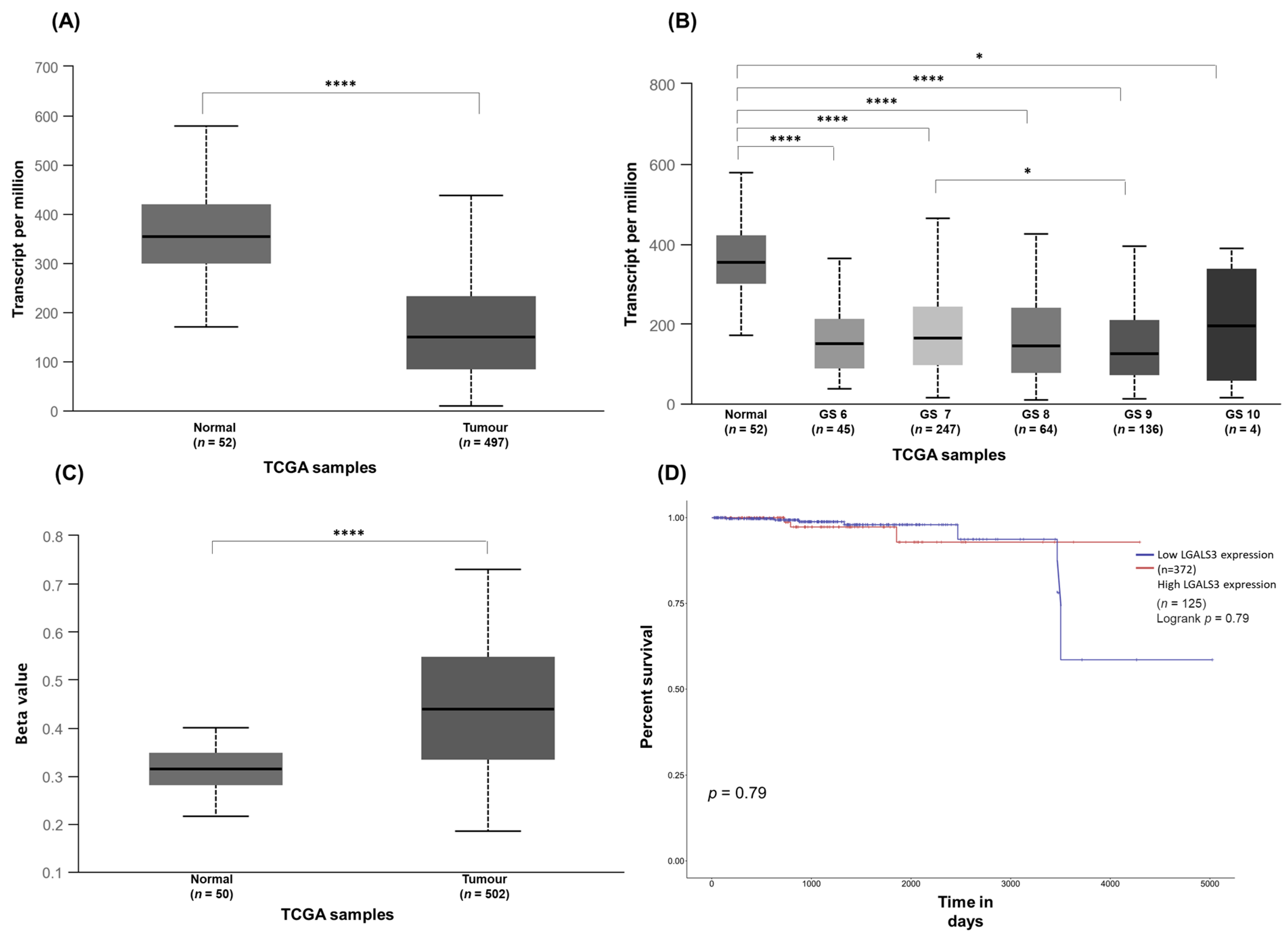
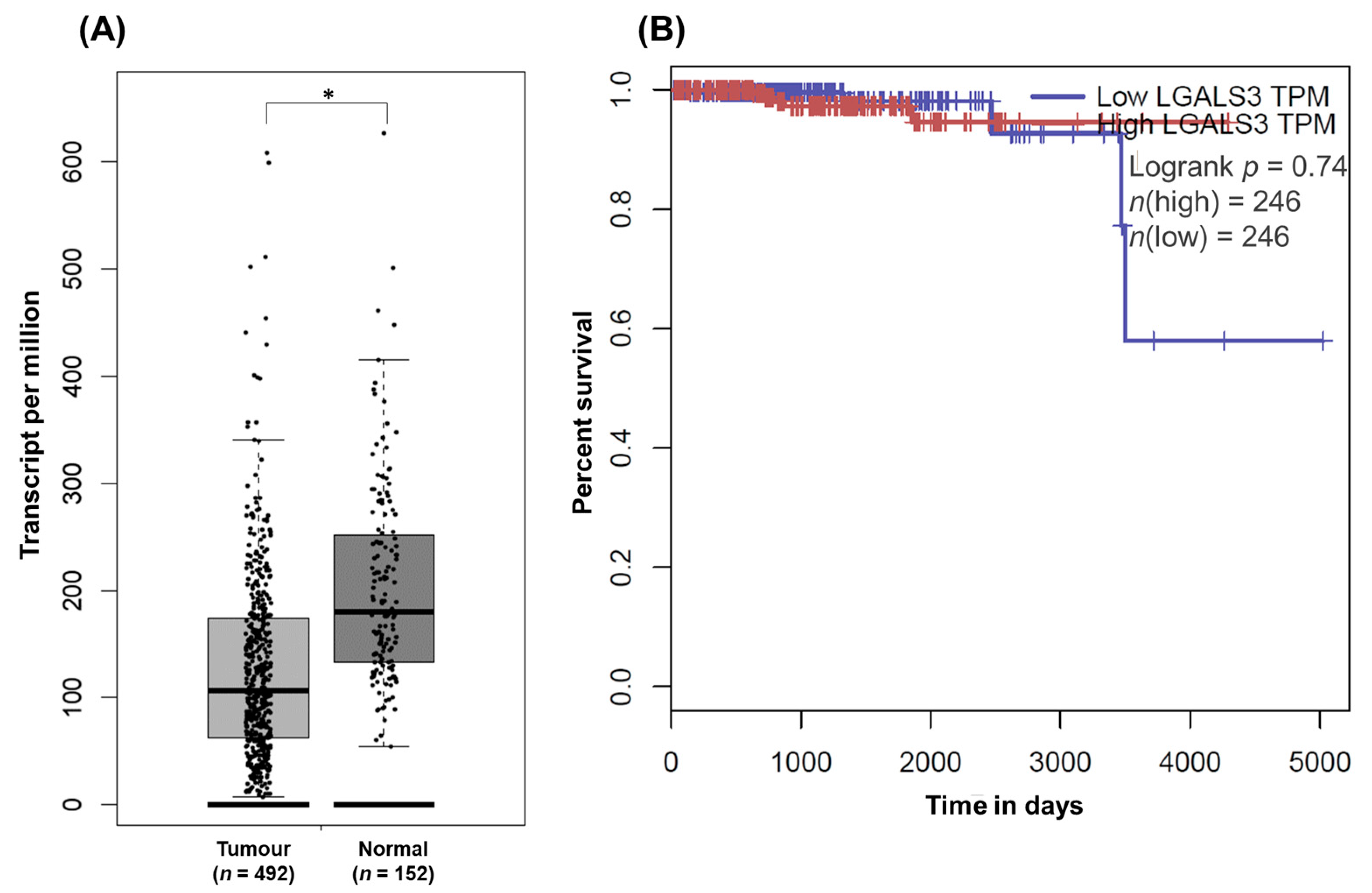
3.3. Exploration of Galectin-3 Expression in PCa and Its Impact on Patient Survival in Public Cancer Databases (In Silico Analysis)
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hara, A.; Niwa, M.; Noguchi, K.; Kanayama, T.; Niwa, A.; Matsuo, M.; Hatano, Y.; Tomita, H. Galectin-3 as a Next-Generation Biomarker for Detecting Early Stage of Various Diseases. Biomolecules 2020, 10, 389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gong, H.C.; Honjo, Y.; Nangia-Makker, P.; Hogan, V.; Mazurak, N.; Bresalier, R.; Raz, A. The NH2 terminus of galectin-3 governs cellular compartmentalization and functions in cancer cells. Cancer Res. 1999, 59, 6239–6245. [Google Scholar] [PubMed]
- Menon, R.P.; Hughes, R.C. Determinants in the N-terminal domains of galectin-3 for secretion by a novel pathway circumventing the endoplasmic reticulum-Golgi complex. JBIC J. Biol. Inorg. Chem. 1999, 264, 569–576. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balan, V.; Nangia-Makker, P.; Kho, D.H.; Wang, Y.; Raz, A. Tyrosine-phosphorylated Galectin-3 Protein Is Resistant to Prostate-specific Antigen (PSA) Cleavage. J. Biol. Chem. 2012, 287, 5192–5198. [Google Scholar] [CrossRef] [Green Version]
- Nangia-Makker, P.; Raz, T.; Tait, L.; Hogan, V.; Fridman, R.; Raz, A. Galectin-3 Cleavage: A Novel Surrogate Marker for Matrix Metalloproteinase Activity in Growing Breast Cancers. Cancer Res. 2007, 67, 11760–11768. [Google Scholar] [CrossRef] [Green Version]
- Akahani, S.; Nangia-Makker, P.; Inohara, H.; Kim, H.R.; Raz, A. Galectin-3: A novel antiapoptotic molecule with a functional BH1 (NWGR) domain of Bcl-2 family. Cancer Res. 1997, 57, 5272–5276. [Google Scholar]
- Popa, S.J.; Stewart, S.E.; Moreau, K. Unconventional secretion of annexins and galectins. Semin. Cell Dev. Biol. 2018, 83, 42–50. [Google Scholar] [CrossRef]
- Oka, N.; Nakahara, S.; Takenaka, Y.; Fukumori, T.; Hogan, V.; Kanayama, H.-O.; Yanagawa, T.; Raz, A. Galectin-3 Inhibits Tumor Necrosis Factor–Related Apoptosis-Inducing Ligand–Induced Apoptosis by Activating Akt in Human Bladder Carcinoma Cells. Cancer Res. 2005, 65, 7546–7553. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.R.; Lin, H.M.; Biliran, H.; Raz, A. Cell cycle arrest and inhibition of anoikis by galectin-3 in human breast epithelial cells. Cancer Res. 1999, 59, 4148–4154. [Google Scholar]
- Shimura, T.; Takenaka, Y.; Fukumori, T.; Tsutsumi, S.; Okada, K.; Hogan, V.; Kikuchi, A.; Kuwano, H.; Raz, A. Implication of Galectin-3 in Wnt Signaling. Cancer Res. 2005, 65, 3535–3537. [Google Scholar] [CrossRef] [Green Version]
- Dagher, S.F.; Wang, J.L.; Patterson, R.J. Identification of galectin-3 as a factor in pre-mRNA splicing. Proc. Natl. Acad. Sci. USA 1995, 92, 1213–1217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Inohara, H.; Akahani, S.; Koths, K.; Raz, A. Interactions between galectin-3 and Mac-2-binding protein mediate cell-cell adhesion. Cancer Res. 1996, 56, 4530–4534. [Google Scholar] [PubMed]
- Sedlář, A.; Trávníčková, M.; Bojarová, P.; Vlachová, M.; Slámová, K.; Křen, V.; Bačáková, L. Interaction between Galectin-3 and Integrins Mediates Cell-Matrix Adhesion in Endothelial Cells and Mesenchymal Stem Cells. Int. J. Mol. Sci. 2021, 22, 5144. [Google Scholar] [CrossRef] [PubMed]
- Ippel, H.; Miller, M.C.; Vértesy, S.; Zheng, Y.; Cañada, F.J.; Suylen, D.; Umemoto, K.; Romanò, C.; Hackeng, T.; Tai, G.; et al. Intra- and intermolecular interactions of human galectin-3: Assessment by full-assignment-based NMR. Glycobiology 2016, 26, 888–903. [Google Scholar] [CrossRef] [Green Version]
- Liu, F.-T.; Hsu, D.K.; Zuberi, R.I.; Hill, P.N.; Shenhav, A.; Kuwabara, I.; Chen, S.-S. Modulation of Functional Properties of Galectin-3 by Monoclonal Antibodies Binding to the Non-Lectin Domains. Biochemistry 1996, 35, 6073–6079. [Google Scholar] [CrossRef]
- Lepur, A.; Salomonsson, E.; Nilsson, U.J.; Leffler, H. Ligand Induced Galectin-3 Protein Self-association. J. Biol. Chem. 2012, 287, 21751–21756. [Google Scholar] [CrossRef] [Green Version]
- Nabi, I.R.; Shankar, J.; Dennis, J.W. The galectin lattice at a glance. J. Cell Sci. 2015, 128, 2213–2219. [Google Scholar] [CrossRef] [Green Version]
- Nangia-Makker, P.; Wang, Y.; Raz, T.; Tait, L.; Balan, V.; Hogan, V.; Raz, A. Cleavage of galectin-3 by matrix metalloproteases induces angiogenesis in breast cancer. Int. J. Cancer 2010, 127, 2530–2541. [Google Scholar] [CrossRef] [Green Version]
- Saraswati, S.; Block, A.S.; Davidson, M.K.; Rank, R.G.; Mahadevan, M.; Diekman, A.B. Galectin-3 is a substrate for prostate specific antigen (PSA) in human seminal plasma. Prostate 2011, 71, 197–208. [Google Scholar] [CrossRef] [Green Version]
- Gao, X.; Liu, J.; Liu, X.; Li, L.; Zheng, J. Cleavage and phosphorylation: Important post-translational modifications of galectin-3. Cancer Metastasis Rev. 2017, 36, 367–374. [Google Scholar] [CrossRef]
- Takenaka, Y.; Fukumori, T.; Yoshii, T.; Oka, N.; Inohara, H.; Kim, H.-R.C.; Bresalier, R.S.; Raz, A. Nuclear Export of Phosphorylated Galectin-3 Regulates Its Antiapoptotic Activity in Response to Chemotherapeutic Drugs. Mol. Cell. Biol. 2004, 24, 4395–4406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, Y.; Shen, R.; Yu, L.; Zheng, X.; Cui, R.; Song, Y.; Wang, D. Roles of galectin-3 in the tumor microenvironment and tumor metabolism (Review). Oncol. Rep. 2020, 44, 1799–1809. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Lee, J.; Hyun, J.W.; Park, J.W.; Joo, H.; Shin, T. Expression and immunohistochemical localization of galectin-3 in various mouse tissues. Cell Biol. Int. 2007, 31, 655–662. [Google Scholar] [CrossRef] [PubMed]
- Al-Maghrabi, J.; Abdelrahman, A.S.; Ghabrah, T.; Butt, N.S.; Al-Maghrabi, B.; Khabaz, M.N. Immunohistochemical expression of galectin-3 is significantly associated with grade, stage and differentiation of endometrial carcinomas. Pathol. Res. Pract. 2017, 213, 348–352. [Google Scholar] [CrossRef]
- Merseburger, A.S.; Kramer, M.W.; Hennenlotter, J.; Simon, P.; Knapp, J.; Hartmann, J.T.; Stenzl, A.; Serth, J.; Kuczyk, M.A. Involvement of decreased Galectin-3 expression in the pathogenesis and progression of prostate cancer. Prostate 2008, 68, 72–77. [Google Scholar] [CrossRef]
- Larsen, L.; Chen, H.-Y.; Saegusa, J.; Liu, F.-T. Galectin-3 and the skin. J. Dermatol. Sci. 2011, 64, 85–91. [Google Scholar] [CrossRef]
- Setayesh, T.; Colquhoun, S.D.; Wan, Y.-J.Y. Overexpression of Galectin-1 and Galectin-3 in hepatocellular carcinoma. Liver Res. 2020, 4, 173–179. [Google Scholar] [CrossRef]
- Blair, B.B.; Funkhouser, A.T.; Goodwin, J.L.; Strigenz, A.M.; Chaballout, B.H.; Martin, J.C.; Arthur, C.M.; Funk, C.R.; Edenfield, W.J.; Blenda, A.V. Increased Circulating Levels of Galectin Proteins in Patients with Breast, Colon, and Lung Cancer. Cancers 2021, 13, 4819. [Google Scholar] [CrossRef]
- Li, Y.; Cheng, D.; Liang, B. Serum Galectin-3 as a Potential Marker for Gastric Cancer. Med. Sci. Monit. Int. Med. J. Exp. Clin. Res. 2015, 21, 755–760. [Google Scholar] [CrossRef] [Green Version]
- Waltregny, D.; Liu, F.-T.; Castronovo, V. Alteration of the cytoplasmic/nuclear expression pattern of galectin-3 correlates with prostate carcinoma progression. Int. J. Cancer 2000, 89, 361–367. [Google Scholar] [CrossRef]
- Sanjuan, X.; Fernandez, P.; Castells, A.; Castronovo, V.; Brule, F.V.D.; Liu, F.; Cardesa, A.; Campo, E. Differential expression of galectin 3 and galectin 1 in colorectal cancer progression. Gastroenterology 1997, 113, 1906–1915. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.K.; Lih, T.M.; Wang, Y.; Hu, Y.; Höti, N.; Chan, D.W.; Zhang, H. Improving the detection of aggressive prostate cancer using immunohistochemical staining of protein marker panels. Am. J. Cancer Res. 2022, 12, 1323–1336. [Google Scholar] [PubMed]
- Araújo-Filho, J.L.; Melo-Junior, M.R.; Beltrão, E.I.C.; De Lima, L.R.A.; Antunes, C.B.L.; De Carvalho, L.B. Immunochemiluminescent detection of galectin-3 in tumoral tissue from prostate. Int. J. Clin. Exp. Pathol. 2013, 6, 1861–1867. [Google Scholar] [PubMed]
- de Melo-Júnior, M.R.; Araújo-Filho, J.L.S.; Lins, C.A.B.; de Pontes-Filho, N.T.; de Carvalho, L.B. Immobilization of Anti-Galectin-3 onto Polysiloxane–Polyvinyl Alcohol Disks for Tumor Prostatic Diseases Diagnosis. Appl. Biochem. Biotechnol. 2010, 160, 2198–2207. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, H.; Banerjee, P.P.; Vasta, G.R. Differential expression of galectins in normal, benign and malignant prostate epithelial cells: Silencing of galectin-3 expression in prostate cancer by its promoter methylation. Biochem. Biophys. Res. Commun. 2007, 358, 241–246. [Google Scholar] [CrossRef]
- Gao, J.; Li, T.; Mo, Z.; Hu, Y.; Yi, Q.; He, R.; Zhu, X.; Zhou, X.; She, S.; Chen, Y. Overexpression of the galectin-3 during tumor progression in prostate cancer and its clinical implications. Int. J. Clin. Exp. Pathol. 2018, 11, 839–846. [Google Scholar]
- Ahmed, H.; Cappello, F.; Rodolico, V.; Vasta, G.R. Evidence of Heavy Methylation in the Galectin 3 Promoter in Early Stages of Prostate Adenocarcinoma: Development and Validation of a Methylated Marker for Early Diagnosis of Prostate Cancer. Transl. Oncol. 2009, 2, 146–156. [Google Scholar] [CrossRef] [Green Version]
- Knapp, J.S.; Lokeshwar, S.D.; Vogel, U.; Hennenlotter, J.; Schwentner, C.; Kramer, M.W.; Stenzl, A.; Merseburger, A.S. Galectin-3 expression in prostate cancer and benign prostate tissues: Correlation with biochemical recurrence. World J. Urol. 2013, 31, 351–358. [Google Scholar] [CrossRef]
- Lima, T.; Ferreira, R.; Freitas, M.; Henrique, R.; Vitorino, R.; Fardilha, M. Integration of Automatic Text Mining and Genomic and Proteomic Analysis to Unravel Prostate Cancer Biomarkers. J. Proteome Res. 2022, 21, 447–458. [Google Scholar] [CrossRef]
- Moch, H.; Amin, M.B.; Berney, D.M.; Compérat, E.M.; Gill, A.J.; Hartmann, A.; Menon, S.; Raspollini, M.R.; Rubin, M.A.; Srigley, J.R.; et al. The 2022 World Health Organization Classification of Tumours of the Urinary System and Male Genital Organs—Part A: Renal, Penile, and Testicular Tumours. Eur. Urol. 2022, 82, 458–468. [Google Scholar] [CrossRef]
- Uhlén, M.; Fagerberg, L.; Hallström, B.M.; Lindskog, C.; Oksvold, P.; Mardinoglu, A.; Sivertsson, Å.; Kampf, C.; Sjöstedt, E.; Asplund, A.; et al. Proteomics. Tissue-Based Map of the Human Proteome. Science 2015, 347, 1260419. [Google Scholar] [CrossRef] [PubMed]
- Tang, Z.; Li, C.; Kang, B.; Gao, G.; Li, C.; Zhang, Z. GEPIA: A web server for cancer and normal gene expression profiling and interactive analyses. Nucleic Acids Res. 2017, 45, W98–W102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chandrashekar, D.S.; Karthikeyan, S.K.; Korla, P.K.; Patel, H.; Shovon, A.R.; Athar, M.; Netto, G.J.; Qin, Z.S.; Kumar, S.; Manne, U.; et al. UALCAN: An update to the integrated cancer data analysis platform. Neoplasia 2022, 25, 18–27. [Google Scholar] [CrossRef]
- Pacis, R.A.; Pilat, M.J.; Pienta, K.J.; Wojno, K.; Raz, A.; Hogan, V.; Cooper, C.R. Decreased galectin-3 expression in prostate cancer. Prostate 2000, 44, 118–123. [Google Scholar] [CrossRef] [PubMed]
- Ellerhorst, J.; Troncoso, P.; Xu, X.-C.; Lee, J.; Lotan, R. Galectin-1 and galectin-3 expression in human prostate tissue and prostate cancer. Urol. Res. 1999, 27, 362–367. [Google Scholar] [CrossRef]
- Slaughter, D.P.; Southwick, H.W.; Smejkal, W. Field cancerization in oral stratified squamous epithelium; clinical implications of multicentric origin. Cancer 1953, 6, 963–968. [Google Scholar] [CrossRef]
- Aran, D.; Camarda, R.; Odegaard, J.; Paik, H.; Oskotsky, B.; Krings, G.; Goga, A.; Sirota, M.; Butte, A.J. Comprehensive analysis of normal adjacent to tumor transcriptomes. Nat. Commun. 2017, 8, 1077. [Google Scholar] [CrossRef] [Green Version]
- Chandran, U.R.; Dhir, R.; Ma, C.; Michalopoulos, G.; Becich, M.; Gilbertson, J. Differences in gene expression in prostate cancer, normal appearing prostate tissue adjacent to cancer and prostate tissue from cancer free organ donors. BMC Cancer 2005, 5, 45. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Nangia-Makker, P.; Tait, L.; Balan, V.; Hogan, V.; Pienta, K.J.; Raz, A. Regulation of Prostate Cancer Progression by Galectin-3. Am. J. Pathol. 2009, 174, 1515–1523. [Google Scholar] [CrossRef] [Green Version]
- Califice, S.; Castronovo, V.; Bracke, M.; van den Brûle, F. Dual activities of galectin-3 in human prostate cancer: Tumor suppression of nuclear galectin-3 vs tumor promotion of cytoplasmic galectin-3. Oncogene 2004, 23, 7527–7536. [Google Scholar] [CrossRef] [Green Version]
- Geisler, C.; Gaisa, N.T.; Pfister, D.; Fuessel, S.; Kristiansen, G.; Braunschweig, T.; Gostek, S.; Beine, B.; Diehl, H.C.; Jackson, A.M.; et al. Identification and Validation of Potential New Biomarkers for Prostate Cancer Diagnosis and Prognosis Using 2D-DIGE and MS. BioMed Res. Int. 2015, 2015, 454256. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristic | PCa n = 48 | Normal Prostate n = 8 | Adjacent-Normal Prostaten = 14 |
|---|---|---|---|
| Age (Years) | 64.10 ± 6.52 | 68.50 ± 16.72 | 65.14 ± 6.26 |
| Serum PSA (NG/ML) | 10.08 ± 4.54 | _ | 9.64 ± 4.27 |
| Combined Gleason Score | 6.79 ± 1.29 | _ | 7.21 ± 0.97 |
| GS5: 10 (20.83%) GS6: 9 (18.75%) GS7: 16 (33.33%) GS8: 7 (14.59%) GS9: 6 (12.5%) | GS6: 3 (21.43%) GS7: 7 (50.00%) GS8: 2 (14.29%) GS9: 2 (14.29%) | ||
| CLINICAL STAGE | pT2a: 5 (10.42%) pT2b: 20 (41.67%) pT2c: 1 (2.08%) pT3a: 15 (31.25%) pT3b: 6 (12.5%) pT4: 1 (2.08%) | pT2b: 6 (42.87%) pT3: 1 (7.14%) pT3a: 5 (35.71%) pT3b: 1 (7.14%) pT4: 1 (7.14%) | |
| Other Pathologies | |||
| Yes | Bladder tumour: 1 (2.08%) Colon Adenocarcinoma: 1 (2.08%) | Papillary carcinoma: 1 (12.50%) Urothelial carcinoma: 4 (50.00%) Bladder carcinoma: 3 (37.50%) | _ |
| No | 46 (95.84%) | _ | 14 (100%) |
| Relapse | _ | ||
| Yes | 17 (35.41%) | 6 (42.86%) | |
| GS5 (n = 2) | |||
| GS6 (n = 4) | |||
| GS7 (n = 6) | |||
| GS8 (n = 3) | |||
| GS9 (n = 2) | |||
| No | 29 (60.42%) | 7 (50%) | |
| GS5 (n = 8) | |||
| GS6 (n = 5) | |||
| GS7 (n = 9) | |||
| GS8 (n = 4) | |||
| Unknown | 2 (4.17%) | 1 (7.14%) | |
| GS7 (n = 1) | |||
| GS9 (n = 1) | |||
| Survival | |||
| Alive | 2 (4.17%) | 1 (7.14%) | |
| GS7 (n = 2) | |||
| Alive with Disease | 7 (14.58%) | 2 (14.29%) | |
| GS5 (n = 1) | |||
| GS6 (n = 3) | |||
| GS7 (n = 2) | |||
| GS8 (n = 1) | |||
| Alive without Disease | 19 (39.58%) | 6 (42.86%) | |
| GS5 (n = 5) | |||
| GS6 (n = 5) | |||
| GS7 (n = 4) | |||
| GS8 (n = 2) | |||
| GS9 (n = 3) | |||
| Death unknown Cause | 3 (6.25%) | 1 (7.14%) | |
| GS7 (n = 1) | |||
| GS8 (n = 2) | |||
| Death with Disease | 10 (20.83%) | 3 (21.43%) | |
| GS7 (n = 6) | |||
| GS8 (n = 2) | |||
| GS9 (n = 2) | |||
| Death without Disease | 5 (10.42%) | 0 | |
| GS5 (n = 4) | |||
| GS6 (n = 1) | |||
| Unknown | 2 (4.17%) | 1 (7.14%) | |
| GS7 (n = 1) | |||
| GS9 (n = 1) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lima, T.; Macedo-Silva, C.; Felizardo, D.; Fraga, J.; Carneiro, I.; Jerónimo, C.; Henrique, R.; Fardilha, M.; Vitorino, R. Gal-3 Protein Expression and Localization in Prostate Tumours. Curr. Oncol. 2023, 30, 2729-2742. https://doi.org/10.3390/curroncol30030206
Lima T, Macedo-Silva C, Felizardo D, Fraga J, Carneiro I, Jerónimo C, Henrique R, Fardilha M, Vitorino R. Gal-3 Protein Expression and Localization in Prostate Tumours. Current Oncology. 2023; 30(3):2729-2742. https://doi.org/10.3390/curroncol30030206
Chicago/Turabian StyleLima, Tânia, Catarina Macedo-Silva, Diana Felizardo, João Fraga, Isa Carneiro, Carmen Jerónimo, Rui Henrique, Margarida Fardilha, and Rui Vitorino. 2023. "Gal-3 Protein Expression and Localization in Prostate Tumours" Current Oncology 30, no. 3: 2729-2742. https://doi.org/10.3390/curroncol30030206
APA StyleLima, T., Macedo-Silva, C., Felizardo, D., Fraga, J., Carneiro, I., Jerónimo, C., Henrique, R., Fardilha, M., & Vitorino, R. (2023). Gal-3 Protein Expression and Localization in Prostate Tumours. Current Oncology, 30(3), 2729-2742. https://doi.org/10.3390/curroncol30030206