
Health Functions of Egg Protein


Abstract
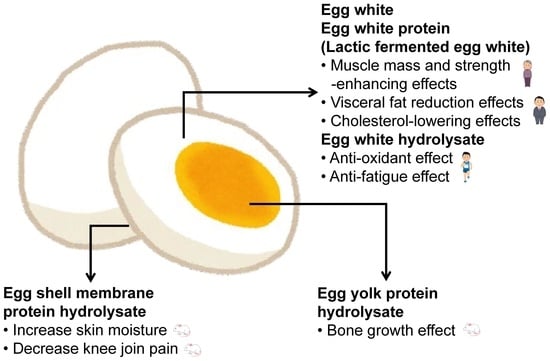
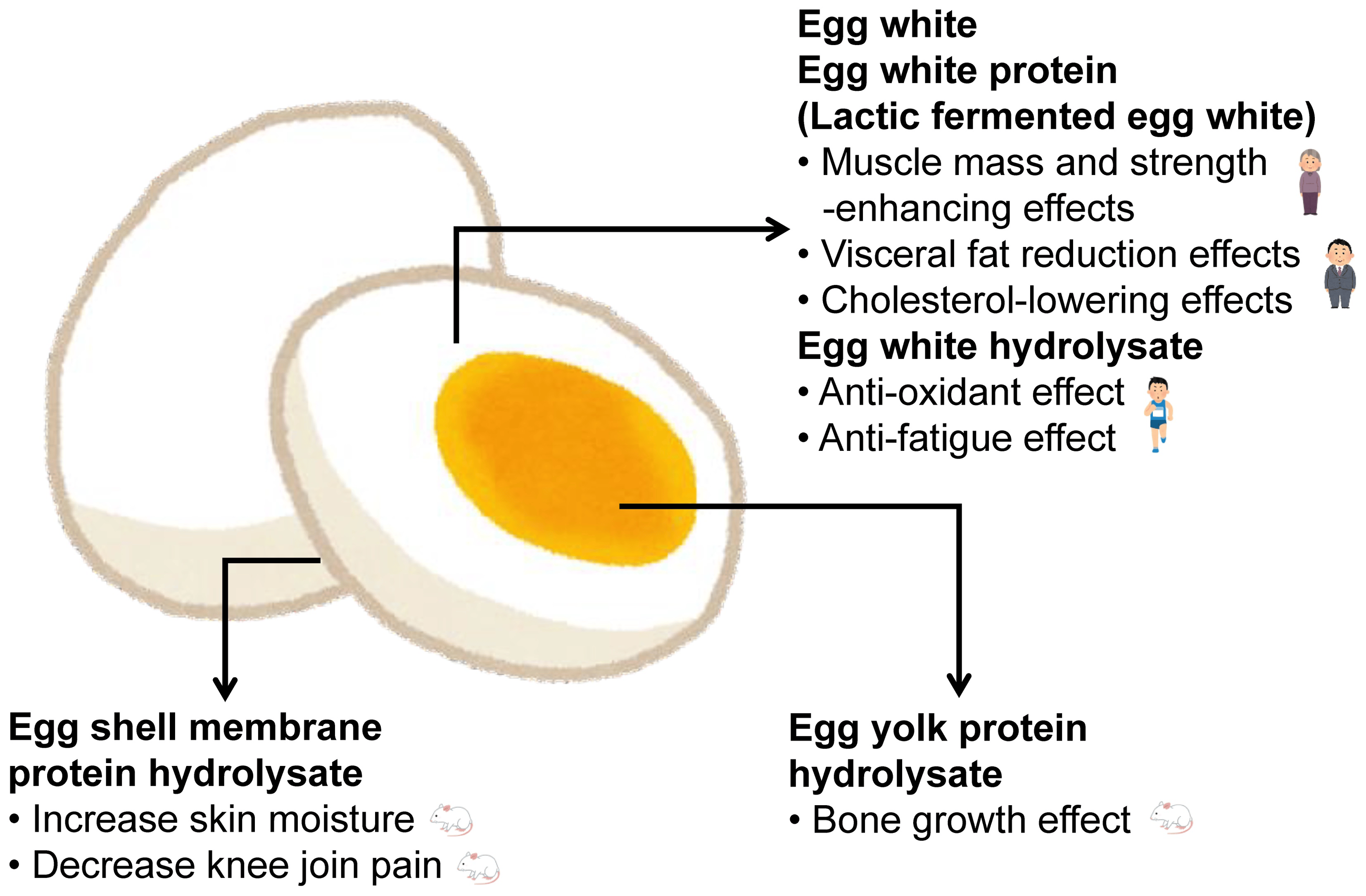
:
1. Introduction

| Casein | Dried Egg White | Soy Protein Isolate | |
|---|---|---|---|
| Ile | 5.0 | 4.4 | 4.0 |
| Leu | 8.5 | 7.3 | 7.0 |
| Lys | 7.2 | 6.1 | 5.5 |
| Met | 2.7 | 3.2 | 1.1 |
| Cys | 0.4 | 2.5 | 1.1 |
| Phe | 4.6 | 5.1 | 4.6 |
| Tyr | 5.2 | 3.9 | 3.5 |
| Thr | 4.0 | 4.0 | 3.7 |
| Trp | 1.1 | 1.3 | 1.2 |
| Val | 6.2 | 5.8 | 4.1 |
| His | 2,7 | 2.1 | 2.4 |
| Arg | 3.4 | 5.0 | 6.9 |
| Ala | 2.7 | 5.3 | 3.6 |
| Asp | 6.3 | 9.3 | 10.0 |
| Gul | 19.0 | 12.0 | 17.0 |
| Gly | 1.7 | 3.2 | 3.6 |
| Pro | 10.0 | 3.3 | 4.7 |
| Ser | 5.2 | 6.0 | 5.1 |

2. Muscle Mass and Strength-Enhancing Effects of Egg White Protein in Combination with Exercise
3. Egg White Protein Improves Lipid Metabolism
3.1. Visceral Fat Reduction Effects

| Time (Weeks) | |||
|---|---|---|---|
| 0 | 12 | ||
| Visceral Fat area (Δcm2) | Milk whey | 0 ± 0 | 1.71 ± 4.00 |
| LE | 0 ± 0 | −8.89 ± 2.75 * | |
| Visceral/Subcutaneous (Δ) | Milk Whey | 0 ± 0 | 0.0260 ± 0.0187 |
| LE | 0 ± 0 | −0.0876 ± 0.0128 * | |
3.2. Cholesterol-Lowering Effects
| Cholesterol Level | ||
|---|---|---|
| In vivo (Rats) | Serum | ↓ |
| Hepatic | ↓ | |
| Stomach | N.S. | |
| Intestinal contents (solid) | ↑ | |
| Intestinal contents (micelle) | ↓ | |
| Intestinal mucosa 1 (upper) | N.S. | |
| Intestinal mucosa 2 | ↓ | |
| Intestinal mucosa 3 | N.S. | |
| Intestinal mucosa 4 (lower) | N.S. | |
| Lymph | ↓ | |
| Fecal | ↑ | |
| In vitro | Micellar solubility | ↓ |
| Bile acid binding | N.S. | |
| Phospholipid binding | N.S. | |
| Transfer to triolein | ↓ | |
| Water holding capacity | ↑ | |
| Settling volume | ↑ | |
| Viscosity | ↓ |
| Egg White Protein | Time (Weeks) | |||
|---|---|---|---|---|
| Intake/Day | 0 | 4 | 8 | |
| Total Cholesterol (Δmg/dL) | 4 g | 0 ± 0 | −4.46 ± 3.36 | 3.11 ± 3.38 |
| 6 g | 0 ± 0 | −7.42 ± 3.76 | −5.97 ± 3.53 | |
| 8 g | 0 ± 0 | −11.3 ± 3.63 | −11.0 ± 3.74 * | |
| LDL-Cholesterol (Δmg/dL) | 4 g | 0 ± 0 | −7.61 ± 2.98 | −2.07 ± 2.91 |
| 6 g | 0 ± 0 | −8.00 ± 3.51 | −9.48 ± 3.12 | |
| 8 g | 0 ± 0 | −10.0 ± 3.00 | −13.7 ± 3.74* | |
4. Antifatigue Effects of Egg White Protein Hydrolysate

{kind=link}
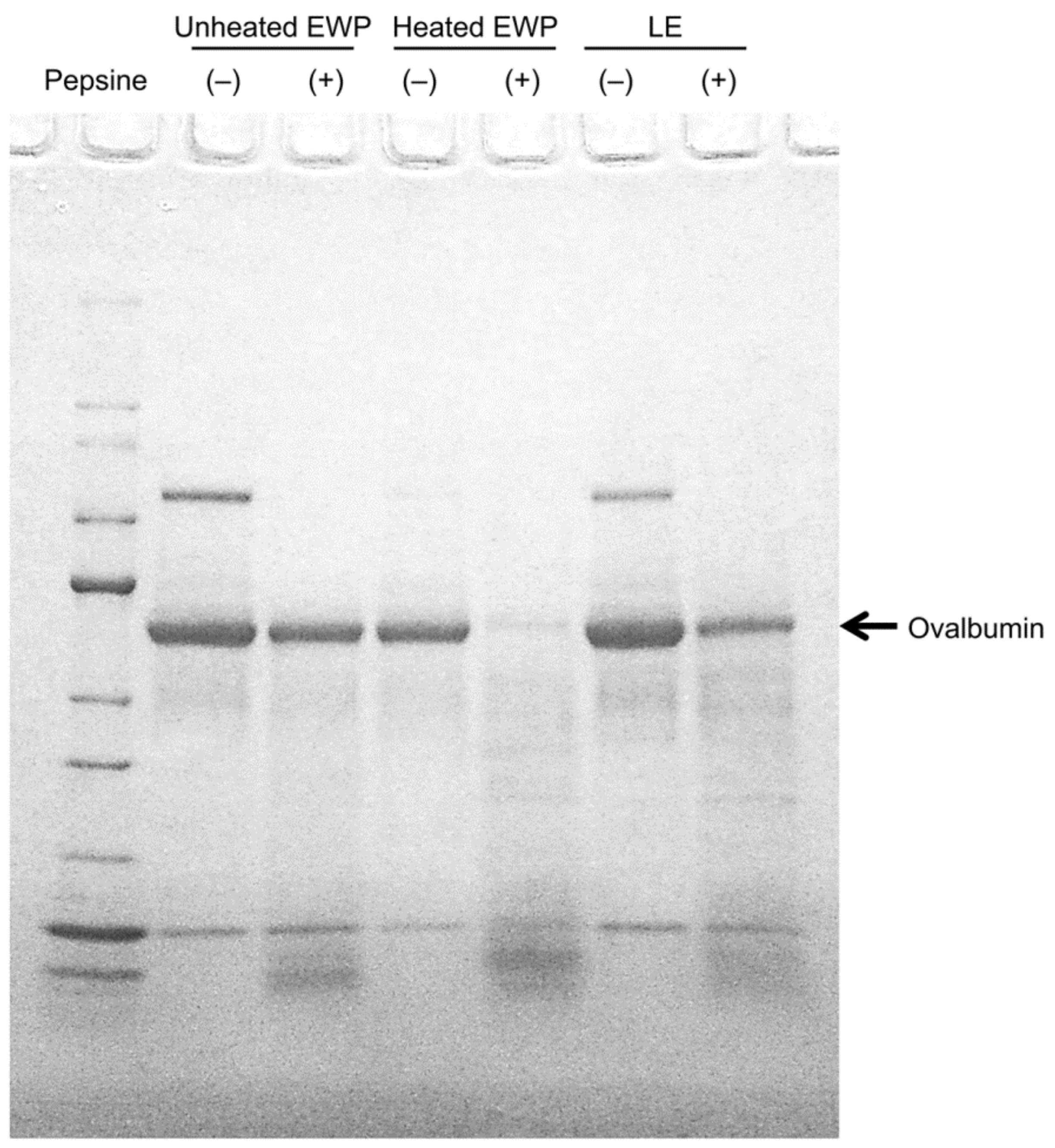{kind=link}
{kind=link}
{kind=link}
5. Absorption and Physiological Functions of Eggshell Membrane Proteins and Hydrolysate
| Casein | ESM-P | ESM-H | |
|---|---|---|---|
| Ile | 4.7 | 3.2 | 2.5 |
| Leu | 8.4 | 4.5 | 3.6 |
| Lys | 7.2 | 3.3 | 2.4 |
| Met | 2.5 | 3.7 | 2.9 |
| Cys | 0.4 | 9.7 | 6.0 |
| Phe | 4.6 | 1.5 | 1.2 |
| Tyr | 5.0 | 1.7 | 1.5 |
| Thr | 3.8 | 5.5 | 4.2 |
| Trp | 1.1 | 3.3 | 2.4 |
| Val | 5.8 | 6.9 | 5.3 |
| His | 2.7 | 4.0 | 2.9 |
| Arg | 3.3 | 7.1 | 5.4 |
| Ala | 2.7 | 2.6 | 2.2 |
| Asp | 6.4 | 8.2 | 6.4 |
| Gul | 19.7 | 12.4 | 9.5 |
| Gly | 1.7 | 5.8 | 4.5 |
| Pro | 9.5 | 9.4 | 7.5 |
| Ser | 5.1 | 5.2 | 4.0 |
6. Health Function of Egg Protein in Basal Research
7. Discussion
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| ALT | alanine aminotransferase |
| AST | aspartate aminotransferase |
| BCAA | branched-chain amino acids |
| DIAAS | digestible indispensable amino acid scores |
| ESM-P | egg shell membrane protein |
| ESM-H | egg shell membrane protein hydrolysate |
| EWH | egg white protein hydrolysates |
| EWP | egg white protein |
| LE | lactic-fermented egg whites |
| PDCAAS | protein digestibility corrected amino acid scores |
| PPAR | peroxisome proliferator-responsive receptor |
References
- Ruxto, C. Recommendation for the use of egg in the diet. Nurs. Stand. 2010, 24, 47–55. [Google Scholar] [CrossRef]
- Sugano, M.; Matsuoka, R. Nutritional viewpoint on egg and cholesterol. Food 2021, 10, 494. [Google Scholar] [CrossRef] [PubMed]
- Ezaki, O.; Sakono, M.; Miyake, M.; Mito, N. Concepts for Reference Intake of Cholesterol in the Japanese Population. J. Jpn. Soc. Nutr. Food Sci. 2005, 58, 69–83. (In Japanese) [Google Scholar] [CrossRef]
- Masuda, Y.; Kokubu, T.; Yamashita, M.; Ikeda, H.; Inoue, S. Egg phosphatidylcholine combined with vitamin B12 improved memory impairment following lesioning nucleus basalis in rats. Life Sci. 1998, 62, 813–822. [Google Scholar] [CrossRef]
- Yin, M.; Matsuoka, R.; Xi, Y.; Wang, X. Comparison of egg yolk and soybean phospholipids on hepatic fatty acid profile and liver protection in rats fed a high fructose diets. Foods 2021, 10, 1569. [Google Scholar] [CrossRef]
- Sakai, S.; Hien, V.T.T.; Tuyen, L.D.; Duc, H.A.; Masuda, Y.; Yamamoto, S. Effects of Eggshell Calcium Supplementation on Bone Mass in Postmenopausal Vietnamese Women. J. Nutr. Sci. Vitaminol. 2017, 63, 120–124. [Google Scholar] [CrossRef] [Green Version]
- Ministry of Education, Culture, Sports, Science and Technology, Standard Tables of Food Composition in Japan (Seventh Revised Version). 2015; Available online: https://www.mext.go.jp/en/policy/science_technology/policy/title01/detail01/1374030.htm (accessed on 9 June 2022).
- Matsuoka, R.; Takahashi, Y.; Kimura, M.; Masuda, Y.; Kunou, M. Heating has no effect on the net protein utilisation from egg whites in rats. Sci. World J. 2017, 2017, 6817196. [Google Scholar] [CrossRef] [Green Version]
- Matsuoka, R.; Shirouchi, B.; Umegatani, M.; Fukuda, M.; Muto, A.; Masuda, Y.; Kunou, M.; Sato, M. Dietary egg white protein increases body protein mass and reduces body fat mass through an acceleration of hepatic β-oxidation in rats. Br. J. Nutr. 2017, 118, 423–430. [Google Scholar] [CrossRef] [Green Version]
- Yoo, J.H.; Kim, J.K.; Yang, H.J.; Park, K.M. Egg shell membrane hydrolysates on the UVB-radiation-induced wrinkle formation in SKH-1 hairless mice. Korean J. Food. Sci. 2015, 35, 58–70. [Google Scholar] [CrossRef] [Green Version]
- Wedekind, K.J.; Ruff, K.J.; Atwell, C.A.; Evans, J.L.; Bendele, A.M. Beneficial effects of natural eggshell membrane (NEM) on m-ultiple indices of arthritis in collagen-induced arthritic rats. Mod. Rheumatol. 2017, 27, 838–848. [Google Scholar] [CrossRef] [Green Version]
- Liao, W.; Jahandideh, F.; Fan, H.; Son, M.; Wu, J. Egg Protein-Derived Bioactive Peptides: Preparation, Efficacy, and Absorption. Adv. Food Nutr. Res. 2018, 85, 1–58. [Google Scholar]
- Legros, J.; Jan, S.; Bonnassie, S.; Gautier, M.; Croguennec, T.; Pezennec, S.; Cochet, M.F.; Nau, F.; Andrews, S.C.; Baron, F. The Role of Ovotransferrin in Egg-White Antimicrobial Activity: A Review. Foods 2021, 10, 823. [Google Scholar] [CrossRef]
- Lee, J.H.; Paik, H.D. Anticancer and immunomodulatory activity of egg proteins and peptides: A review. Poult. Sci. 2019, 98, 6505–6516. [Google Scholar] [CrossRef]
- Mine, Y. Egg proteins and peptides in human health--chemistry, bioactivity and production. Curr. Pharm. Des. 2007, 13, 875–884. [Google Scholar] [CrossRef]
- Sanlier, N.; Üstün, D. Egg consumption and health effects: A narrative review. J. Food Sci. 2021, 86, 4250–4261. [Google Scholar] [CrossRef]
- Marventano, S.; Godos, J.; Tieri, M.; Ghelfi, F.; Titta, L.; Lafranconi, A.; Gambera, A.; Alonzo, E.; Sciacca, S.; Buscemi, S.; et al. Egg consumption and human health: An umbrella review of observational studies. Int. J. Food Sci. Nutr. 2020, 71, 325–331. [Google Scholar] [CrossRef]
- Shirouchi, B.; Matsuoka, R. Alleviation of Metabolic Syndrome with Dietary Egg White Protein. J. Oleo Sci. 2019, 68, 517–524. [Google Scholar] [CrossRef] [Green Version]
- Puglist, M.J.; Luz Fernandez, M. The Health Benefits of Egg Protein. Nutrients 2022, 14, 2904. [Google Scholar] [CrossRef]
- Arimitsu, K.; Kojima, T.; Matsuoka, R.; Narita, K.; Idei, A. The good function of lactic fermented egg white “Lacty Egg”. Jpn. J. Food. Eng. 2015, 16, 79–82. (In Japanese) [Google Scholar] [CrossRef] [Green Version]
- Matsuoka, R.; Takahashi, Y.; Muto, A.; Kimura, M. Heated egg white has no effect, but lactic fermented and unheated egg white reduces abdominal fat in rats. Lipids Health Dis. 2019, 18, 187. [Google Scholar] [CrossRef] [Green Version]
- Matsuoka, R.; Kurihara, H.; Nishijima, N.; Oda, Y.; Handa, A. Egg white hydrolysate retains the nutritional values and is quickly absorbed in rats. Sci. World J. 2019, 2019, 5475302. [Google Scholar] [CrossRef]
- Kato, Y.; Sawada, A.; Numao, S.; Suzuki, M. Chronic effects of light resistance exercise after ingestion of a high-protein snack on increase of skeletal muscle mass and strength in young adults. J. Nutr. Sci. Vitaminol. 2011, 57, 233–238. [Google Scholar] [CrossRef] [Green Version]
- Takanami, Y.; Kawai, Y.; Nakata, C.; Nishiyama, H.; Matsuoka, R. The combined effect of excise and lactic fermented egg white on the skeletal muscle function and muscle mass in middle-aged and elderly women. In Proceedings of the 70th Annual Meeting of the Japanese Society of Physical Fitness and Sports Medicine, Wakayama, Japan, 18–20 September 2015; Tajima, H., Ed.; The Japanese Society of Physical Fitness and Sports Medicine: Tokyo, Japan, 2015. [Google Scholar]
- Van Vliet, S.; Shy, E.L.; Abou Sawan, S.; Beals, J.W.; West, D.W.; Skinner, S.K.; Ulanov, A.V.; Li, Z.; Paluska, S.A.; Parsons, C.M.; et al. Consumption of whole eggs promotes greater stimulation of postexercise muscle protein synthesis than consumption of isonitrogenous amounts of egg whites in young men. Am. J. Clin. Nutr. 2017, 106, 1401–1412. [Google Scholar] [CrossRef]
- Bagheri, R.; Hooshmand Moghadam, B.; Ashtary-Larky, D.; Forbes, S.C.; Candow, D.G.; Galpin, A.J.; Eskandari, M.; Kreider, R.B.; Wong, A. Whole Egg vs. Egg White Ingestion during 12 weeks of Resistance Training in Trained Young Males: A Randomized Controlled Trial. J. Strength Cond. Res. 2021, 35, 411–419. [Google Scholar] [CrossRef]
- Fritzen, A.M.; Lundsgaard, A.M.; Kiens, B. Tuning fatty acid oxidation in skeletal muscle with dietary fat and exercise. Nat. Rev. Endocrinol. 2020, 16, 683–696. [Google Scholar] [CrossRef]
- Matsuoka, R.; Shirouchi, B.; Kawamura, S.; Baba, S.; Shiratake, S.; Nagata, K.; Imaizumi, K.; Sato, M. Dietary egg white protein inhibits lymphatic lipid transport in thoracic lymph duct-cannulated rats. J. Agric. Food Chem. 2014, 62, 10694–10700. [Google Scholar] [CrossRef]
- Matsuoka, R. Health and nutritional function of egg protein. Amino Acid Res. 2019, 13, 27–32. (In Japanese) [Google Scholar]
- Matsuoka, R.; Kimura, M.; Arimitsu, K.; Kojima, T. New Health Functions of Egg White Protein (Lactic Fermented Egg White) and Its Application. J. Jpn. Soc. Nutr. Food Sci. 2021, 74, 147–154. (In Japanese) [Google Scholar] [CrossRef]
- Sakai, K.; Ushiyama, Y.; Manabe, S. Peptic and pancreatic digestibility of raw and heat-treated hen’s egg white protein. Jpn. J. Pediatr. Allegy Clin. Immunol. 1999, 13, 36–42. (In Japanese) [Google Scholar] [CrossRef]
- Handa, A.; Gennadios, A.; Hanna, M.A.; Weller, C.L.; Kuroda, N. Physical and molecular properties of egg-white lipid films. J. Food Sci. 1999, 64, 860–864. [Google Scholar] [CrossRef]
- Takahashi, Y.; Matsuoka, R.; Kimura, M.; Masuda, Y.; Kunou, M.; Wada, Y.; Takahashi, Y.; Utsunomiya, K. Health function of egg white protein (No. 3: Mechanism for visceral fat lowering effects). In Proceedings of the 66th Annual Meeting of Japan Society of Nutrition and Food Science, Sendai, Japan, 18–20 May 2012; Komai, M., Ed.; Japan Society of Nutrition and Food Science: Tokyo, Japan, 2012. [Google Scholar]
- Rao, P.S.; Nolasco, E.; Handa, A.; Naldrett, M.J.; Alvarez, S.; Majumder, K. Effect of pH and heat treatment on the antioxidant activity of egg white protein-derived peptides after simulated in-vitro gastrointestinal digestion. Antioxidants 2020, 9, 1114. [Google Scholar] [CrossRef] [PubMed]
- Yamada, Y.; Muraki, A.; Oie, M.; Kanegawa, N.; Oda, A.; Sawashi, Y.; Kaneko, K.; Yoshikawa, M.; Goto, T.; Takahashi, N.; et al. Soymorphin-5, a soy-derived mu-opioid peptide, decreases glucose and triglyceride levels through activating adiponectin and PPARalpha systems in diabetic KKAy mice. Am. J. Physiol. Endocrinol. Metab. 2012, 302, E433–E440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goto, T.; Mori, A.; Nagaoka, S. Soluble soy protein peptic hydrolysate stimulates adipocyte differentiation in 3T3-L1 cells. Mol. Nutr. Food Res. 2013, 57, 1435–1445. [Google Scholar] [CrossRef] [PubMed]
- Kohno, M.; Hirotsuka, M.; Kito, M.; Matsuzawa, Y. Decreases in serum triacylglycerol and visceral fat mediated by dietary soybean beta-conglycinin. J. Atheroscler. Thromb. 2006, 13, 247–255. [Google Scholar] [CrossRef] [Green Version]
- Ono, T.; Murakoshi, M.; Suzuki, N.; Iida, N.; Ohdera, M.; Iigo, M.; Yoshida, T.; Sugiyama, K.; Nishino, H. Potent anti-obesity effect of enteric-coated lactoferrin: Decrease in visceral fat accumulation in Japanese men and women with abdominal obesity after 8-week administration of enteric coated lactofferin tablets. Br. J. Nutr. 2010, 104, 1688–1695. [Google Scholar] [CrossRef] [Green Version]
- Moriyama, T.; Kishimoto, K.; Nagai, K.; Urade, R.; Ogawa, T.; Utsumi, S.; Maruyama, N.; Maebashi, M. Soybean beta-conglycinin diet suppresses serum triglyceride levels in normal and genetically obese mice by induction of beta-oxidation, downregulation of fatty acid synthase, and inhibition of triglyceride absorption. Biosci. Biotechnol. Biochem. 2004, 68, 352–359. [Google Scholar] [CrossRef]
- Matsuoka, R.; Kimura, M.; Muto, A.; Masuda, Y.; Sato, M.; Imaizumi, K. Mechanism for the cholesterol-lowering action of egg white protein in rats. Biosci. Biotechnol. Biochem. 2008, 72, 1506–1512. [Google Scholar] [CrossRef] [Green Version]
- Matsuoka, R.; Kamachi, K.; Usuda, M.; Wang, W.; Masuda, Y.; Kunou, M.; Tanaka, A.; Utsunomiya, K. Lactic-Fermented Egg White Improved Visceral Fat Obesity in Japanese Subjects: A Double-Blind, Placebo-Controlled study. Lipids Health Dis. 2017, 16, 237. [Google Scholar] [CrossRef] [Green Version]
- Ballesteros, M.N.; Cabrera, R.M.; Saucedo, M.S.; Fernandez, M.L. Dietary cholesterol does not increase biomarkers for chronic disease in a pediatric population from northern Mexico. Am. J. Clin. Nutr. 2005, 80, 855–861. [Google Scholar] [CrossRef] [Green Version]
- Vega-Lopez, S.; Conde, K.; Ramjiganesh, T.; Shachter, N.S.; Lus Fernandez, M. Men classified as hypo- or hyperresponders to dietary cholesterol feeding exhibit differences in lipoprotein metabolism. J. Nutr. 2003, 133, 1036–1042. [Google Scholar]
- Matsuoka, R. Property of Phytosterols and Development of Its Containing Mayonnaise-Type Dressing. Foods 2022, 11, 1141. [Google Scholar] [CrossRef]
- Matsuoka, R.; Masuda, Y.; Takeuchi, A.; Marushima, R.; Onuki, M. Minimal effective dose of plant sterol on serum cholesterol concentration in Japanese subjects and safety evaluation of plant sterol supplemented in mayonnaise. J. Oleo Sci. 2004, 53, 17–27. [Google Scholar] [CrossRef] [Green Version]
- Høie, L.H.; Morgenstern, E.C.; Gruenwald, J.; Graubaum, H.J.; Busch, R.; Lüder, W.; Zunft, H.J. A double-blind placebo-controlled clinical trial compares the cholesterol-lowering effects of two different soy protein preparations in hypercholesterolemic subjects. Eur. J. Nutr. 2005, 44, 65–71. [Google Scholar] [CrossRef]
- Anderson, J.W.; Bush, H.M. Soy protein effects on serum lipoproteins: A quality assessment and meta-analysis of randomized, controlled studies. J. Am. Coll. Nutr. 2011, 30, 79–91. [Google Scholar] [CrossRef]
- Soliman, G.A. Dietary Fiber, Atherosclerosis, and Cardiovascular Disease. Nutrients 2019, 11, 1155. [Google Scholar] [CrossRef] [Green Version]
- Lin, S.M.; Li, F.J.; Yuangsoi, B.; Doolgindachbaporn, S. Effect of dietary phospholipid levels on growth, lipid metabolism, and antioxidative status of juvenile hybrid snakehead (Channa argus × Channa maculata). Fish Physiol. Biochem. 2018, 44, 401–410. [Google Scholar] [CrossRef]
- Asato, L.; Wang, M.F.; Chan, Y.C.; Yeh, S.H.; Chung, H.M.; Chung, S.Y.; Chida, S.; Uezato, T.; Suzuki, I.; Yamagata, N.; et al. Effect of egg white on serum cholesterol concentration in young women. J. Nutr. Sci. Vitaminol. 1996, 42, 87–96. [Google Scholar] [CrossRef]
- Oda, H. Functions of sulfur-containing amino acids in lipid metabolism. J. Nutr. 2006, 136, 1666S–1669S. [Google Scholar] [CrossRef] [Green Version]
- Aoyama, Y.; Amano, N.; Yoshida, A. Cholesterol synthesis and degradation in normal rats fed a cholesterol-free diet with excess cystine. Lipids 1999, 34, 583–589. [Google Scholar] [CrossRef]
- Nagaoka, S.; Masaoka, M.; Zhang, Q.; Hasegawa, M.; Watanabe, K. Egg ovomucin attenuates hypercholesterolemia in rats and inhibits cholesterol absorption in Caco-2 cells. Lipids 2002, 37, 267–272. [Google Scholar] [CrossRef]
- Nakamura, R.; Takeyama, M.; Nakamura, K.; Umemura, O. Constituent proteins of globulin fraction obtained from egg white. Agric. Biol. Chem. 1980, 44, 2357–2363. [Google Scholar]
- Zhu, H.; He, Z.; Kwek, E.; Liu, J.; Hao, W.; Liang, N.; Zhao, Y.; Ma, K.Y.; He, W.S.; Chen, Z.Y. Dose-dependent increase in liver cholesterol but not plasma cholesterol from consumption of one to five eggs and no effects from egg whites on liver or plasma cholesterol in hamsters. J. Agric. Food. Chem. 2018, 66, 12805–12814. [Google Scholar] [CrossRef]
- Matsuoka, R.; Usuda, M.; Masuda, Y.; Kunou, M.; Utsunomiya, K. Lactic-Fermented Egg White Reduced Serum Cholesterol Concentrations in Mildly Hypercholesterolemic Japanese Men: A Double-Blind, Parallel-Arm Design. Lipids Health Dis. 2017, 16, 101. [Google Scholar] [CrossRef] [Green Version]
- Mizuno, S.; Matsuura, K.; Gotou, T.; Nishimura, S.; Kajimoto, O.; Yabune, M.; Kajimoto, Y.; Yamamoto, N. Antihypertensive effect of casein hydrolysate in a placebo-controlled study in subjects with high-normal blood pressure and mild hypertension. Br. J. Nutr. 2005, 94, 84–91. [Google Scholar] [CrossRef] [Green Version]
- Moro, T.; Brightwell, C.R.; Velarde, B.; Fry, C.S.; Nakayama, K.; Sanbongi, C.; Volpi, E.; Rasmussen, B.B. Whey Protein Hydrolysate Increases Amino Acid Uptake, mTORC1 Signaling, and Protein Synthesis in Skeletal Muscle of Healthy Young Men in a Randomized Crossover Trial. J. Nutr. 2019, 149, 1149–1158. [Google Scholar] [CrossRef]
- Kwak, J.H.; Kim, M.; Lee, E.; Lee, S.H.; Ahn, C.W.; Lee, J.H. Effects of black soy peptide supplementation on blood pressure and oxidative stress: A randomized controlled trial. Hypertens. Res. 2013, 36, 1060–1066. [Google Scholar] [CrossRef] [Green Version]
- Watabe, K. Function of egg white peptide “Peptifine®”. Jpn. Food Sci. 2013, 52, 17–24. (In Japanese) [Google Scholar]
- WHO. Energy and protein requirements (WHO technical report series, No. 724). In Report of a Joint FAO/WHO/UNU Expert Consultation; World Health Organization: Geneva, Switzerland, 1985. [Google Scholar]
- Seligson, F.H.; Rotruck, J.T. Tissue nonprotein sulfhydryl content and weight gain of rats as affected by dietary methionine level. J. Nutr. 1983, 13, 98–104. [Google Scholar] [CrossRef]
- Steinberg, D.; Parthasarathy, S.; Carew, T.E.; Khoo, J.C.; Witztum, J.L. Beyond cholesterol. Modifications of low-density lipoprotein that increase its atherogenicity. N. Engl. J. Med. 1989, 320, 915–924. [Google Scholar]
- Matsumoto, K.; Koba, T.; Hamada, K.; Sakurai, M.; Higuchi, T.; Miyata, H. Branched-chain amino acid supplementation attenuates muscle soreness, muscle damage and inflammation during an intensive training program. J. Sports Med. Phys. Fitness 2009, 49, 424–431. [Google Scholar]
- Kato, Y.; Sawada, A.; Numao, S.; Miyauchi, R.; Imaizumi, K.; Sakamoto, S.; Suzuki, M. Effect of light resistance exercise after ingestion of a high-protein snack on plasma branched-chain amino acid concentrations in young adult females. J. Nutr. Sci. Vitaminol. 2009, 55, 106–111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harada, R.; Taguchi, Y.; Urashima, K.; Sato, M.; Ohmori, T.; Morimatsu, F. Enhancement of Swimming Endurance in Mice by Chicken Breast Extract. Jpn. Soc. Nutr. Food. Sci. 2002, 55, 73–78. (In Japanese) [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Wang, X.; Zhao, Z. Effect of whey protein hydrolysates with different molecular weight on fatigue induced by swimming exercise in mice. J. Sci. Food. Agr. 2014, 94, 126–130. [Google Scholar] [CrossRef] [PubMed]
- Davalos, A.; Miguel, M.; Bartolome, B.; Lopez-Fandino, R. Antioxidant activity of peptides derived from egg white proteins by enzymatic hydrolysis. J. Food Prot. 2004, 67, 1939–1944. [Google Scholar] [CrossRef]
- Sun, S.; Niu, H.; Yang, T.; Lin, Q.; Luo, F.; Ma, M. Antioxidant and anti-fatigue activities of egg white peptides prepared by pepsin digestion. J. Sci. Food Agr. 2014, 94, 3195–3200. [Google Scholar] [CrossRef]
- Matsuoka, R.; Kimura, M.; Uno, S.; Shidara, H.; Kunou, M. Egg white hydrolysate improves fatigue due to short-term swimming load test in mice. Food Sci. Nutr. 2018, 6, 2314–2320. [Google Scholar] [CrossRef]
- Sugiyama, K.; Kuriki, D.; Matsuoka, R.; Masuda, Y.; Kunou, M.; Daigo, E. Evaluation of the Functionality of Egg White Peptide as a Dietary Supplement for Sports. Nippon. Shokuhin Kagaku Kogaku Kaishi 2020, 67, 245–256. (In Japanese) [Google Scholar] [CrossRef]
- Oe, M.; Sakamoto, H.; Nishiyama, H.; Sasahara, R.; Masuda, Y.; Adachi, M.; Nishiyama, T. Egg white hydrolyzate reduces mental fatigue: Randomized, double-blind, controlled study. BMC Res. Notes 2020, 13, 443. [Google Scholar] [CrossRef]
- Banergee, A.; Panda, S.; Sidhantha, M.; Chakravati, S.; Chaudhuri, B.; Bhattaharjee, S. Utilisation of egg shell membrane as an adsorbent for car bondioxide. Int. J. Global Warm. 2010, 2, 252–261. [Google Scholar] [CrossRef]
- Suyama, K.; Fukazawa, Y.; Umetsu, Y. A new biomaterial, hen egg shell membrane, to eliminate heavy metal ion from their dilute waste solution. Appl. Biochem. Biotechnol. 1994, 45–46, 871–879. [Google Scholar] [CrossRef]
- Horiike, S. Efficacy of health food, Feature and usage of Ransyo. New Food Ind. 2000, 42, 44–48. (In Japanese) [Google Scholar]
- Matsuoka, R.; Kurihara, H.; Yukawa, H.; Sasahara, R. Eggshell membrane protein can be absorbed and utilised in the bodies of rats. BMC Res. Notes 2019, 12, 258. [Google Scholar] [CrossRef] [Green Version]
- Lee, D.E.; Jung, T.H.; Jo, Y.N.; Yun, S.S.; Han, K.S. Enzymatic hydrolysis of egg white protein exerts a hypotensive effect in spontaneously hypertensive rats. Food. Sci. Anim. Resour. 2019, 39, 980–987. [Google Scholar] [CrossRef] [Green Version]
- Miguel, M.; López-Fandiño, R.; Ramos, M.; Aleixandre, A. Long-term intake of egg white hydrolysate attenuates the development of hypertension in spontaneously hypertensive rats. Life Sci. 2006, 78, 2960–2966. [Google Scholar] [CrossRef]
- Manso, M.A.; Miguel, M.; Even, J.; Hernández, R.; Aleixandre, A.; López-Fandiño, R. Effect of the long-term intake of an egg white hydrolysate on the oxidative status and blood lipid profile of spontaneously hypertensive rats. Food Chem. 2008, 109, 361–367. [Google Scholar] [CrossRef]
- Garcés-Rimón, M.; González, C.; Vera, G.; Uranga, J.A.; López-Fandiño, R.; López-Miranda, V.; Miguel, M. Pepsin Egg White Hydrolysate Improves Glucose Metabolism Complications Related to Metabolic Syndrome in Zucker Fatty Rats. Nutrients 2018, 10, 441. [Google Scholar] [CrossRef] [Green Version]
- Ochiai, M.; Azuma, Y. Egg White Hydrolysate Improves Glucose Tolerance in Type-2 Diabetic NSY Mice. J. Nutr. Sci. Vitaminol. 2017, 63, 422–429. [Google Scholar] [CrossRef] [Green Version]
- Jahandideh, F.; Zani, S.C.C.; Son, M.; Proctor, S.D.; Davidge, S.T.; Chan, C.B.; Wu, J. Egg white hydrolysate enhances insulin sensitivity in high-fat diet-induced insulin-resistant rats via Akt activation. Br. J. Nutr. 2019, 122, 14–24. [Google Scholar] [CrossRef]
- Yamamoto, S.; Kita, T.; Yamagata, N.; Kokubu, T.; Shinjo, S.; Asato, L. Favorable effects of EWP on lipid metabolism in rats and mice. Nutr. Res. 1993, 13, 1453–1457. [Google Scholar] [CrossRef]
- Martinez, C.S.; Piagette, J.T.; Escobar, A.G.; Martín, Á.; Palacios, R.; Peçanha, F.M.; Vassallo, D.V.; Exley, C.; Alonso, M.J.; Salaices, M.; et al. Egg White Hydrolysate: A new putative agent to prevent vascular dysfunction in rats following long-term exposure to aluminum. Food. Chem. Toxicol. 2019, 133, 110799. [Google Scholar] [CrossRef]
- Garcés-Rimón, M.; González, C.; Uranga, J.A.; López-Miranda, V.; López-Fandiño, R.; Miguel, M. Pepsin Egg White hydrolysate ameliorates obesity-related oxidative stress, inflammation and steatosis in zucker fatty rats. PLoS ONE 2016, 11, e0151193. [Google Scholar] [CrossRef] [Green Version]
- Jiang, Z.; Kimura, Y.; Shirouchi, B.; Tanaka, Y.; Tsai, W.T.; Yuan, X.; Sato, M. Dietary egg white protein hydrolysate improves orotic acid-induced fatty liver in rats by promoting hepatic phospholipid synthesis and microsomal triglyceride transfer protein expression. J. Nutr. Biochem. 2021, 98, 108820. [Google Scholar] [CrossRef]
- Rizzetti, D.A.; Fernandez, F.; Moreno, S.; Uranga, O.J.A.; Peçanha, F.M.; Vera, G.; Vassallo, D.V.; Castro, M.M.; Wiggers, G.A. Egg white hydrolysate promotes neuroprotection for neuropathic disorders induced by chronic exposure to low concentrations of mercury. Brain. Res. 2016, 1646, 482–489. [Google Scholar] [CrossRef]
- Martinez, C.S.; Alterman, C.D.C.; Vera, G.; Márquez, A.; Uranga, J.A.; Peçanha, F.M.; Vassallo, D.V.; Exley, C.; Mello-Carpes, P.B.; Miguel, M.; et al. Egg white hydrolysate as a functional food ingredient to prevent cognitive dysfunction in rats following long-term exposure to aluminum. Sci. Rep. 2019, 9, 1868. [Google Scholar] [CrossRef] [Green Version]
- Kobayashi, T.; Koie, H.; Watanabe, A.; Ino, A.; Watabe, K.; Kim, M.; Kanayama, K.; Otsuji, K. Effects of food enriched with egg yolk hydrolysate (bone peptide) on bone metabolism in orchidectomized dogs. J. Vet. Med. Sci. 2015, 77, 503–506. [Google Scholar] [CrossRef] [Green Version]
- World Health Organization. Life Expectancy and Healthy Life Expectancy Data by Country. Available online: https://apps.who.int/gho/data/view.main.SDG2016LEXv?lang=en (accessed on 9 June 2022).
- Lee, J.H.; Kim, H.J.; Ahn, D.U.; Paik, H.D. Improved immune-enhancing activity of egg white protein ovotransferrin after enzyme hydrolysis. J. Anim. Sci. Technol. 2021, 63, 1159–1168. [Google Scholar] [CrossRef]
- Van den Heuvel, E.; Murphy, J.L.; Appleton, K.M. Towards a food-based intervention to increase protein intakes in older adults: Challenges to and facilitators of egg consumption. Nutrients 2018, 10, 1409. [Google Scholar] [CrossRef] [Green Version]
- Sivixay, S.; Bai, G.; Tsuruta, T.; Nishino, N. Cecum microbiota in rats fed soy, milk, meat, fish, and egg proteins with prebiotic oligosaccharides. AIMS Microbiol. 2021, 7, 1. [Google Scholar] [CrossRef]
- Devries, M.C.; Phillips, S.M. Supplemental protein in support of muscle mass and health: Advantage whey. J. Food Sci. 2015, 80 (Suppl. S1), A8–A15. [Google Scholar] [CrossRef]
- Messina, M.; Lynch, H.; Dickinson, J.M.; Reed, K.E. No Difference between the Effects of Supplementing with Soy Protein Versus Animal Protein on Gains in Muscle Mass and Strength in Response to Resistance Exercise. Int. J. Sport Nutr. Exerc. Metab. 2018, 28, 674–685. [Google Scholar] [CrossRef] [Green Version]
- Aoyama, T.; Nosaka, N.; Kasai, M. Research on the nutritional characteristics of medium-chain fatty acids. J. Med. Investig. 2007, 54, 385–388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kobayashi, M.; Kawano, T.; Ukawa, Y.; Sagesaka, Y.M.; Fukuhara, I. Green tea beverages enriched with catechins with a galloyl moiety reduce body fat in moderately obese adults: A randomized double-blind placebo-controlled trial. Food Funct. 2016, 7, 498–507. [Google Scholar] [CrossRef] [PubMed]
- Nagatomo, A.; Nishida, N.; Fukuhara, I.; Noro, A.; Kozai, Y.; Sato, H.; Matsuura, Y. Daily intake of rosehip extract decreases abdominal visceral fat in preobese subjects: A randomized, double-blind, placebo-controlled clinical trial. Diabetes Metab. Syndr. Obes. 2015, 8, 147–156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsuzuki, K.; Asao, H.; Ban, S.; Ikematsu, H.; Shionoya, K. Evaluation of the effect of prepared soymilk containing soy protein on serum lipids and the safety of its excessive intake. J. Nutr. Food. 2003, 6, 25–38. [Google Scholar]
- Sirtori, C.R.; Lovati, M.R.; Manzoni, C.; Monetti, M.; Pazzucconi, F.; Gatti, E. Soy and cholesterol reduction: Clinical experience. J. Nutr. 1995, 125, 598S–605S. [Google Scholar]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Matsuoka, R.; Sugano, M. Health Functions of Egg Protein. Foods 2022, 11, 2309. https://doi.org/10.3390/foods11152309
Matsuoka R, Sugano M. Health Functions of Egg Protein. Foods. 2022; 11(15):2309. https://doi.org/10.3390/foods11152309
Chicago/Turabian StyleMatsuoka, Ryosuke, and Michihiro Sugano. 2022. "Health Functions of Egg Protein" Foods 11, no. 15: 2309. https://doi.org/10.3390/foods11152309
APA StyleMatsuoka, R., & Sugano, M. (2022). Health Functions of Egg Protein. Foods, 11(15), 2309. https://doi.org/10.3390/foods11152309