
Ensiling Improved the Colonization and Degradation Ability of Irpex lacteus in Wheat Straw
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fungal Strain and Preparation of I. lacteus Spawn
2.2. Substrates and Ensiling Procedure
2.3. Fermentation with I. lacteus
2.4. Microbial Analysis
2.5. Chemical Analyses and Enzymatic Hydrolysis
2.6. Statistical Analyses
3. Results and Discussion
3.1. Chemical and Microbial Compositions of Raw Materials before Inoculation
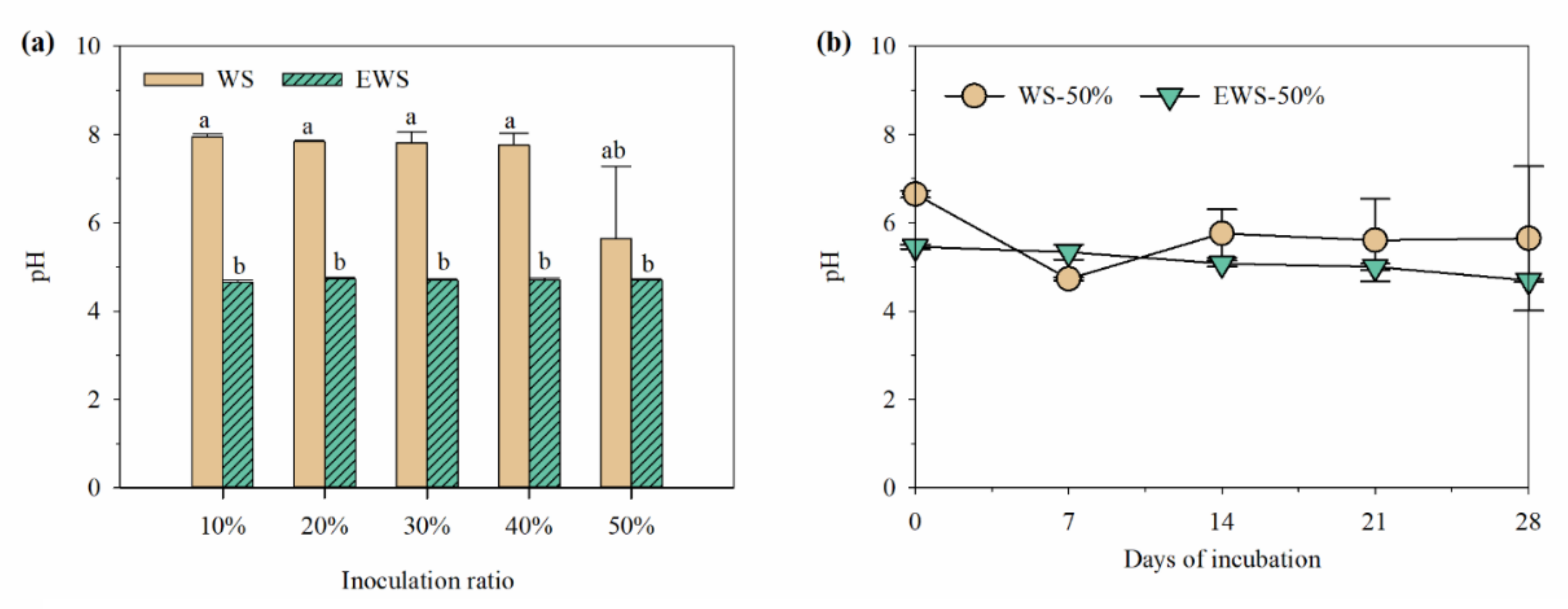
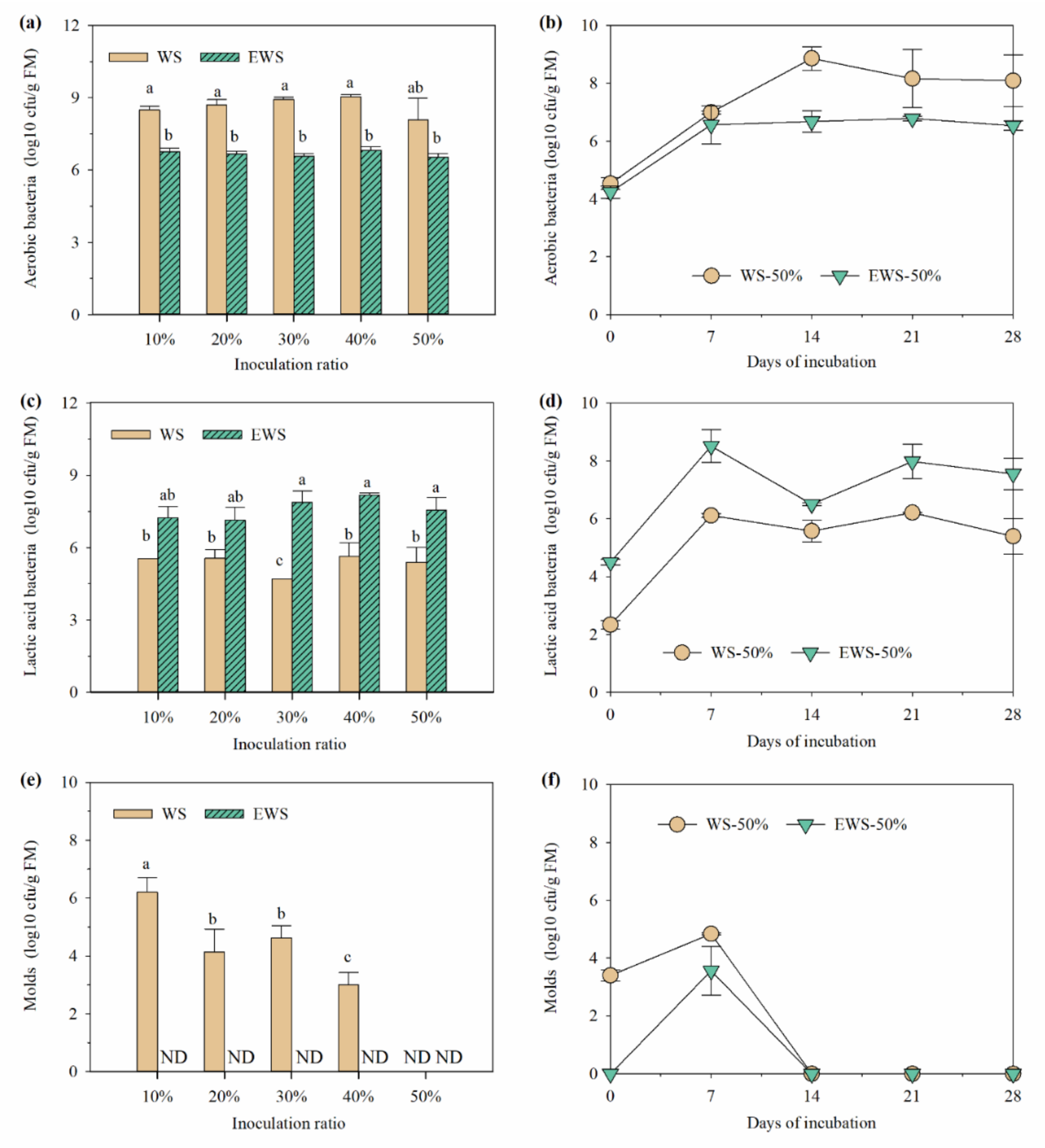
3.2. pH and Microbial Composition during Incubation with I. lacteus
3.3. Chemical Composition during Incubation with I. lacteus
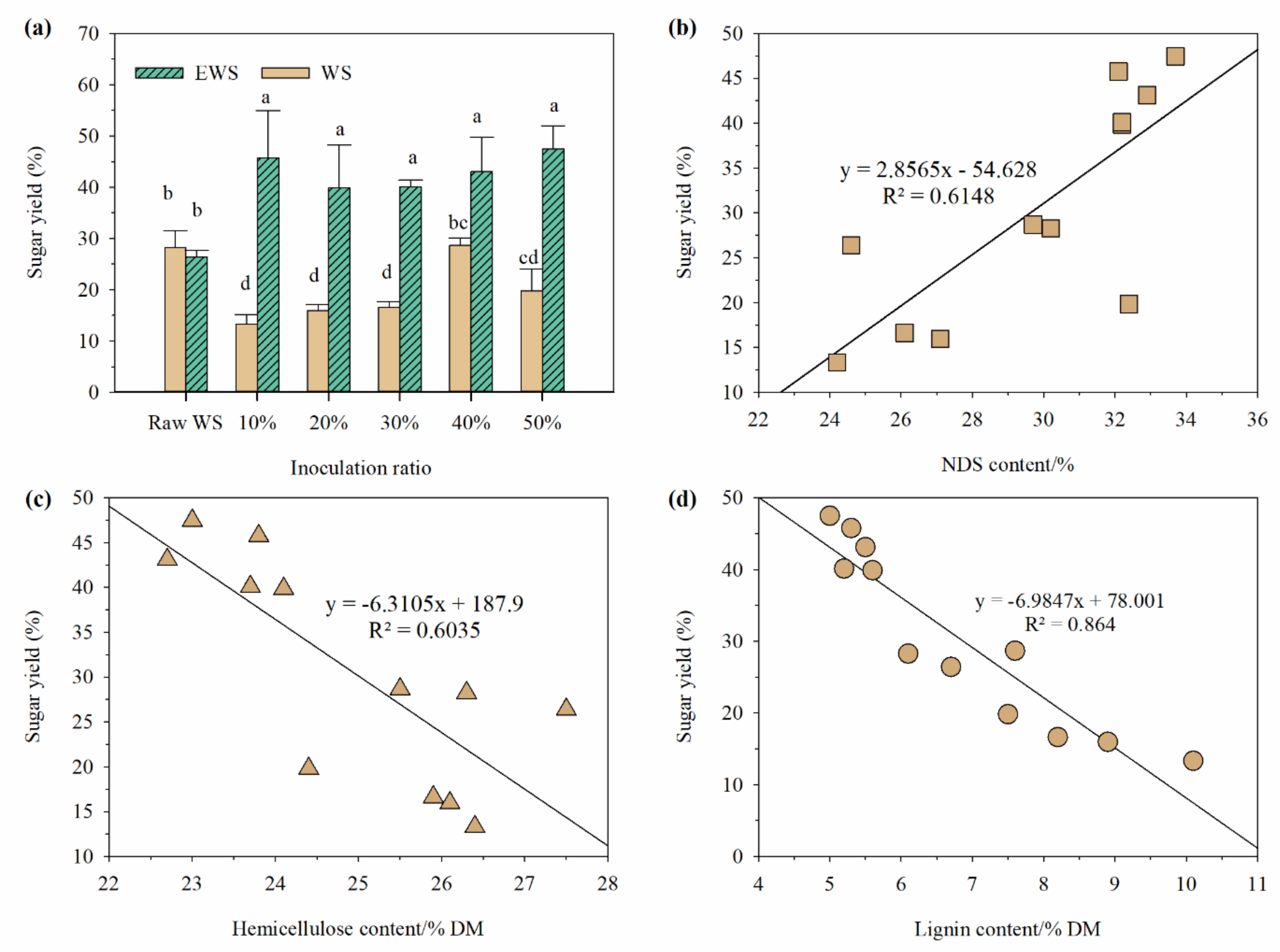
3.4. Sugar Yield of Enzymatic Hydrolysis before and after Incubation with I. lacteus
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Talebnia, F.; Karakashev, D.; Angelidaki, I. Production of bioethanol from wheat straw: An overview on pretreatment, hydrolysis and fermentation. Bioresour. Technol. 2010, 101, 4744–4753. [Google Scholar] [CrossRef] [PubMed]
- Niu, D.Z.; Zuo, S.S.; Jiang, D.; Tian, P.J.; Zheng, M.L.; Xu, C.C. Treatment using white rot fungi changed the chemical composition of wheat straw and enhanced digestion by rumen microbiota in vitro. Anim. Feed Sci. Technol. 2018, 237, 46–54. [Google Scholar] [CrossRef]
- Houfani, A.A.; Anders, N.; Spiess, A.C.; Baldrian, P.; Benallaoua, S. Insights from enzymatic degradation of cellulose and hemicellulose to fermentable sugars—A review. Biomass Bioenergy 2020, 134, 105481. [Google Scholar] [CrossRef]
- Severe, J.; ZoBell, D. Technical aspects for the utilization of small grain straws as feed energy sources for ruminants: Emphasis on beef cattle. Utah State Univ. Ext. Publ. 2012, 4, 95–104. [Google Scholar]
- Dos Santos, A.C.; Ximenes, E.; Kim, Y.; Ladisch, M.R. Lignin-enzyme interactions in the hydrolysis of lignocellulosic biomass. Trends Biotechnol. 2019, 37, 518–531. [Google Scholar] [CrossRef] [PubMed]
- Kainthola, J.; Podder, A.; Fechner, M.; Goel, R. An overview of fungal pretreatment processes for anaerobic digestion: Applications, bottlenecks and future needs. Bioresour. Technol. 2021, 321, 124397. [Google Scholar] [CrossRef]
- Gu, X.; Fu, X.; Chen, S. Hydrothermal liquefaction conversion of lignocelluloses with enhanced fungal pretreatment. Ind. Crops Prod. 2021, 162, 113268. [Google Scholar] [CrossRef]
- González, C.; Wu, Y.; Zuleta-Correa, A.; Jaramillo, G.; Vasco-Correa, J. Biomass to value-added products using microbial consortia with white-rot fungi. Bioresour. Technol. Rep. 2021, 16, 100831. [Google Scholar] [CrossRef]
- Van Kuijk, S.J.A.; Sonnenberg, A.S.M.; Baars, J.J.P.; Hendriks, W.H.; Cone, J.W. Fungal treatment of lignocellulosic biomass: Importance of fungal species, colonization and time on chemical composition and in vitro rumen degradability. Anim. Feed Sci. Technol. 2015, 209, 40–50. [Google Scholar] [CrossRef]
- Zheng, M.; Zuo, S.; Niu, D.; Jiang, D.; Tao, Y.; Xu, C. Effect of four species of white rot fungi on the chemical composition and in vitro rumen degradability of naked oat straw. Waste Biomass Valori. 2020, 12, 435–443. [Google Scholar] [CrossRef]
- Zhao, X.; Wang, F.; Fang, Y.; Zhou, D.; Wang, S.; Wu, D.; Wang, L.; Zhong, R. High-potency white-rot fungal strains and duration of fermentation to optimize corn straw as ruminant feed. Bioresour. Technol. 2020, 312, 123512. [Google Scholar] [CrossRef] [PubMed]
- Vasco-Correa, J.; Ge, X.; Li, Y. Fungal pretreatment of non-sterile miscanthus for enhanced enzymatic hydrolysis. Bioresour. Technol. 2016, 203, 118–123. [Google Scholar] [CrossRef]
- Sasaki, C.; Yamanaka, S. Novel sterilization method combining food preservative use and low temperature steaming for treatment of lignocellulosic biomass with white rot fungi. Ind. Crops Prod. 2020, 155, 112765. [Google Scholar] [CrossRef]
- Song, L.; Ma, F.; Zeng, Y.; Zhang, X.; Yu, H. The promoting effects of manganese on biological pretreatment with Irpex lacteus and enzymatic hydrolysis of corn stover. Bioresour. Technol. 2013, 135, 89–92. [Google Scholar] [CrossRef] [PubMed]
- Ni, K.; Wang, Y.; Pang, H.; Cai, Y. Effect of cellulase and lactic acid bacteria on fermentation quality and chemical composition of wheat straw silage. Am. J.Plant Sci. 2014, 05, 1877–1884. [Google Scholar] [CrossRef] [Green Version]
- Niu, D.; Zheng, M.; Zuo, S.; Jiang, D.; Xu, C. Effects of maize meal and limestone on the fermentation profile and aerobic stability of smooth bromegrass (Bromus inermis Leyss) silage. Grass Forage Sci. 2018, 73, 622–629. [Google Scholar] [CrossRef]
- Digman, M.F.; Shinners, K.J.; Casler, M.D.; Dien, B.S.; Hatfield, R.D.; Jung, H.-J.G.; Muck, R.E.; Weimer, P.J. Optimizing on-farm pretreatment of perennial grasses for fuel ethanol production. Bioresour. Technol. 2010, 101, 5305–5314. [Google Scholar] [CrossRef]
- Desta, S.T.; Yuan, X.; Li, J.; Shao, T. Ensiling characteristics, structural and nonstructural carbohydrate composition and enzymatic digestibility of Napier grass ensiled with additives. Bioresour. Technol. 2016, 221, 447–454. [Google Scholar] [CrossRef] [Green Version]
- Liu, S.; Ge, X.; Liu, Z.; Li, Y. Effect of harvest date on Arundo donax L.(giant reed) composition, ensilage performance, and enzymatic digestibility. Bioresour. Technol. 2016, 205, 97–103. [Google Scholar] [CrossRef] [Green Version]
- Chen, Q.; Marshall, M.N.; Geib, S.M.; Tien, M.; Richard, T.L. Effects of laccase on lignin depolymerization and enzymatic hydrolysis of ensiled corn stover. Bioresour. Technol. 2012, 117, 186–192. [Google Scholar] [CrossRef]
- Tian, P.; Vyas, D.; Niu, D.; Zuo, S.; Jiang, D.; Xu, C. Effects of calcium carbonate on the fermentation quality and aerobic stability of total mixed ration silage. J. Anim. Feed Sci. 2020, 29, 167–174. [Google Scholar] [CrossRef]
- He, L.; Zhou, W.; Xing, Y.; Pian, R.; Chen, X.; Zhang, Q. Improving the quality of rice straw silage with Moringa oleifera leaves and propionic acid: Fermentation, nutrition, aerobic stability and microbial communities. Bioresour. Technol. 2020, 299, 122579. [Google Scholar] [CrossRef] [PubMed]
- Niu, D.; Zuo, S.; Ren, J.; Huhetaoli; Zheng, M.; Jiang, D.; Xu, C. Novel strategy to improve the colonizing ability of Irpex lacteus in non-sterile wheat straw for enhanced rumen and enzymatic digestibility. Appl. Microbiol. Biotechnol. 2020, 104, 1347–1355. [Google Scholar] [CrossRef] [PubMed]
- Zuo, S.; Jiang, D.; Niu, D.; Zheng, M.; Tao, Y.; Xu, C. A lower cost method of preparing corn stover for Irpex lacteus treatment by ensiling with lactic acid bacteria. Asian-Australas J. Anim. Sci. 2020, 33, 1273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, M.; Li, R.; Wang, Y.; Cui, X.; Niu, D.; Yang, F.; Xu, C. Ensiling with rumen fluid promoted Irpex lacteus colonization on the non-sterile naked oat straw for enhanced lignocellulose degradation and enzymatic hydrolysis. Biochem. Eng. J. 2022, 183, 108462. [Google Scholar] [CrossRef]
- Villa, R.; Ortega Rodriguez, L.; Fenech, C.; Anika, O.C. Ensiling for anaerobic digestion: A review of key considerations to maximise methane yields. Renew. Sust. Energ. Rev. 2020, 134, 110401. [Google Scholar] [CrossRef]
- Andrade, E.; Mendes-Ferreira, A.; Botelho, S.; Marques, G.; Cone, J.W.; Rodrigues, M.; Ferreira, L. Preservation of fungal-treated cowpea straw in association with discarded apple by ensilage process. Waste Biomass Valori. 2021, 12, 5533–5543. [Google Scholar] [CrossRef]
- Liaud, N.; Giniés, C.; Navarro, D.; Fabre, N.; Crapart, S.; Gimbert, I.H.; Levasseur, A.; Raouche, S.; Sigoillot, J.C. Exploring fungal biodiversity: Organic acid production by 66 strains of filamentous fungi. Fungal Biol. Biotechnol. 2014, 1, 1. [Google Scholar] [CrossRef] [Green Version]
- Iimura, Y.; Abe, H.; Otsuka, Y.; Sato, Y.; Habe, H. Bacterial community coexisting with white-rot fungi in decayed wood in nature. Curr. Microbiol. 2021, 78, 3212–3217. [Google Scholar] [CrossRef]
- Mao, L.; Sonnenberg, A.S.; van Arkel, J.; Cone, J.W.; de Vos, R.C.; Marchal, J.L.M.; Hendriks, W.H. Storage temperature and time and its influence on feed quality of fungal treated wheat straw. Anim. Feed Sci. Technol. 2021, 272, 114749. [Google Scholar] [CrossRef]
- Wiberth, C.C.; Casandra, A.Z.C.; Zhiliang, F.; Gabriela, H. Oxidative enzymes activity and hydrogen peroxide production in white-rot fungi and soil-borne micromycetes co-cultures. Ann. Microbiol. 2019, 69, 171–181. [Google Scholar] [CrossRef]
- Isroi, I.; Millati, R.; Niklasson, C.; Cayanto, C.; Taherzadeh, M.J.; Lundquist, K. Biological treatment of Lignocelluloses with white-rot funghi and its applications. BioResources 2011, 6, 5224–5259. [Google Scholar] [CrossRef]
- Wang, J.; Chen, X.; Chio, C.; Yang, C.; Su, E.; Jin, Y.; Cao, F.; Qin, W. Delignification overmatches hemicellulose removal for improving hydrolysis of wheat straw using the enzyme cocktail from Aspergillus niger. Bioresour. Technol. 2019, 274, 459–467. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Item | Wheat Straw | Ensiled Wheat Straw |
|---|---|---|
| WSC (% DM) | 5.15 ± 0.243 | 0.59 ± 0.038 |
| NDS (% DM) | 28.4 ± 0.23 | 25.2 ± 0.36 |
| Hemicellulose (% DM) | 27.9 ± 0.28 | 29.7 ± 0.31 |
| Cellulose (% DM) | 37.9 ± 0.33 | 41.3 ± 0.27 |
| ADL (% DM) | 6.81 ± 0.17 | 6.99 ± 0.23 |
| pH value | 6.88 ± 0.076 | 5.55 ± 0.043 |
| Lactic acid (% DM) | ND | 2.48 ± 0.137 |
| Acetic acid (% DM) | ND | 3.40 ± 0.211 |
| Butyric acid (% DM) | ND | 2.67 ± 0.173 |
| LAB (log10 CFU/g FM) | 3.52 ± 0.127 | 6.87 ± 0.393 |
| AB (log10 CFU/g FM) | 6.86 ± 0.251 | 6.13 ± 0.174 |
| Molds (log10 CFU/g FM) | 5.12 ± 0.181 | ND |
| Treatment | Items | NDS | Hemicellulose | Cellulose | Lignin |
|---|---|---|---|---|---|
| Wheat straw | NDS | 1 | −0.527 | −0.828 ** | −0.666 * |
| Hemicellulose | −0.527 | 1 | 0.061 | −0.138 | |
| Cellulose | −0.828 ** | 0.061 | 1 | 0.642 * | |
| Lignin | −0.666 * | −0.138 | 0.642 * | 1 | |
| Ensiled wheat straw | NDS | 1 | −0.928 ** | −0.720 * | −0.918 ** |
| Hemicellulose | −0.928 ** | 1 | 0.422 | 0.920 ** | |
| Cellulose | −0.720 * | 0.422 | 1 | 0.467 | |
| Lignin | −0.918 ** | 0.920 ** | 0.467 | 1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Niu, D.; Zhu, P.; Pan, T.; Yu, C.; Li, C.; Ren, J.; Xu, C. Ensiling Improved the Colonization and Degradation Ability of Irpex lacteus in Wheat Straw. Int. J. Environ. Res. Public Health 2022, 19, 13668. https://doi.org/10.3390/ijerph192013668
Niu D, Zhu P, Pan T, Yu C, Li C, Ren J, Xu C. Ensiling Improved the Colonization and Degradation Ability of Irpex lacteus in Wheat Straw. International Journal of Environmental Research and Public Health. 2022; 19(20):13668. https://doi.org/10.3390/ijerph192013668
Chicago/Turabian StyleNiu, Dongze, Peng Zhu, Tingting Pan, Changyong Yu, Chunyu Li, Jianjun Ren, and Chuncheng Xu. 2022. "Ensiling Improved the Colonization and Degradation Ability of Irpex lacteus in Wheat Straw" International Journal of Environmental Research and Public Health 19, no. 20: 13668. https://doi.org/10.3390/ijerph192013668