
Secondary Terpenes in Cannabis sativa L.: Synthesis and Synergy

Abstract
:1. Introduction
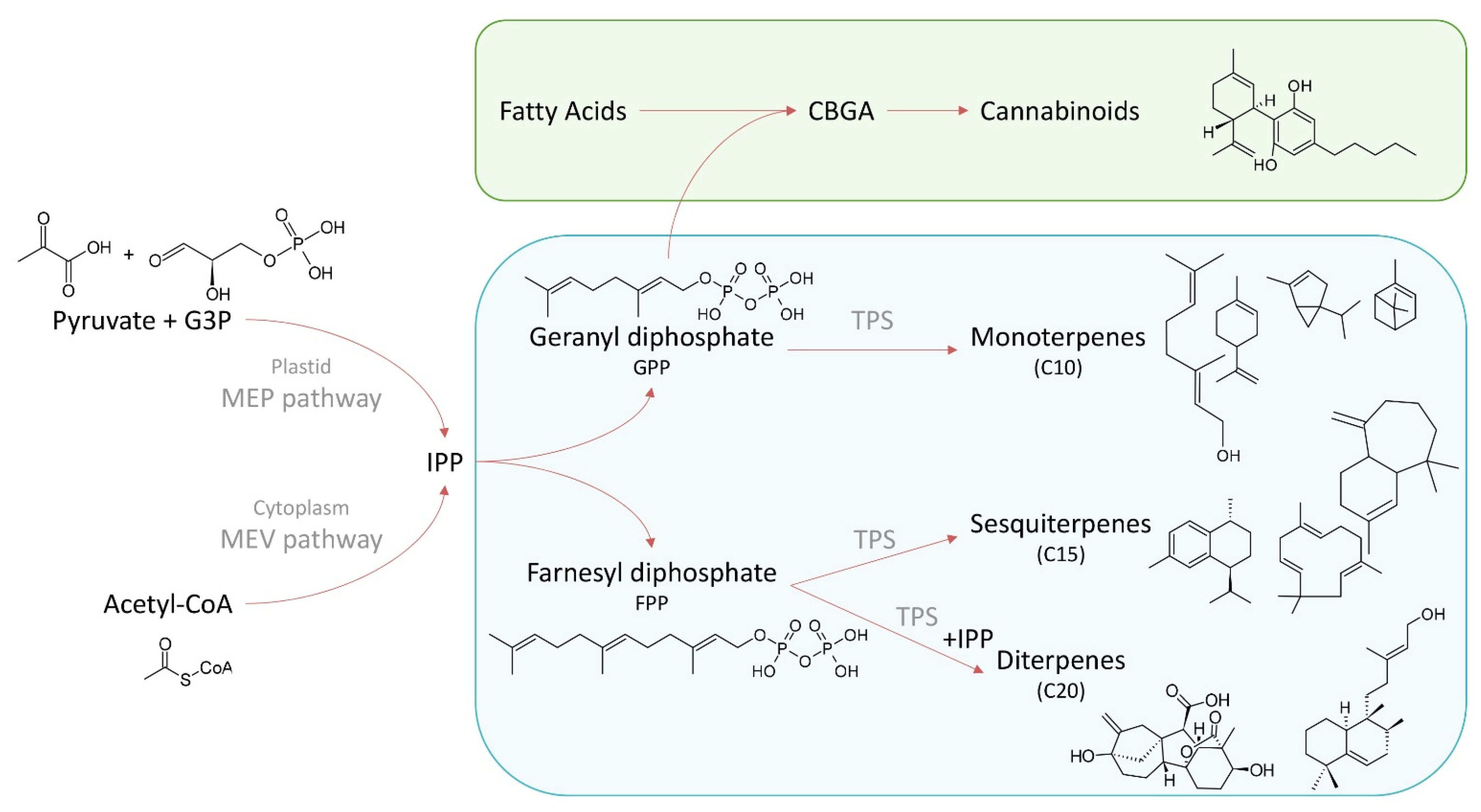
2. Terpene Biosynthesis
3. Terpene Diversity in Cannabis

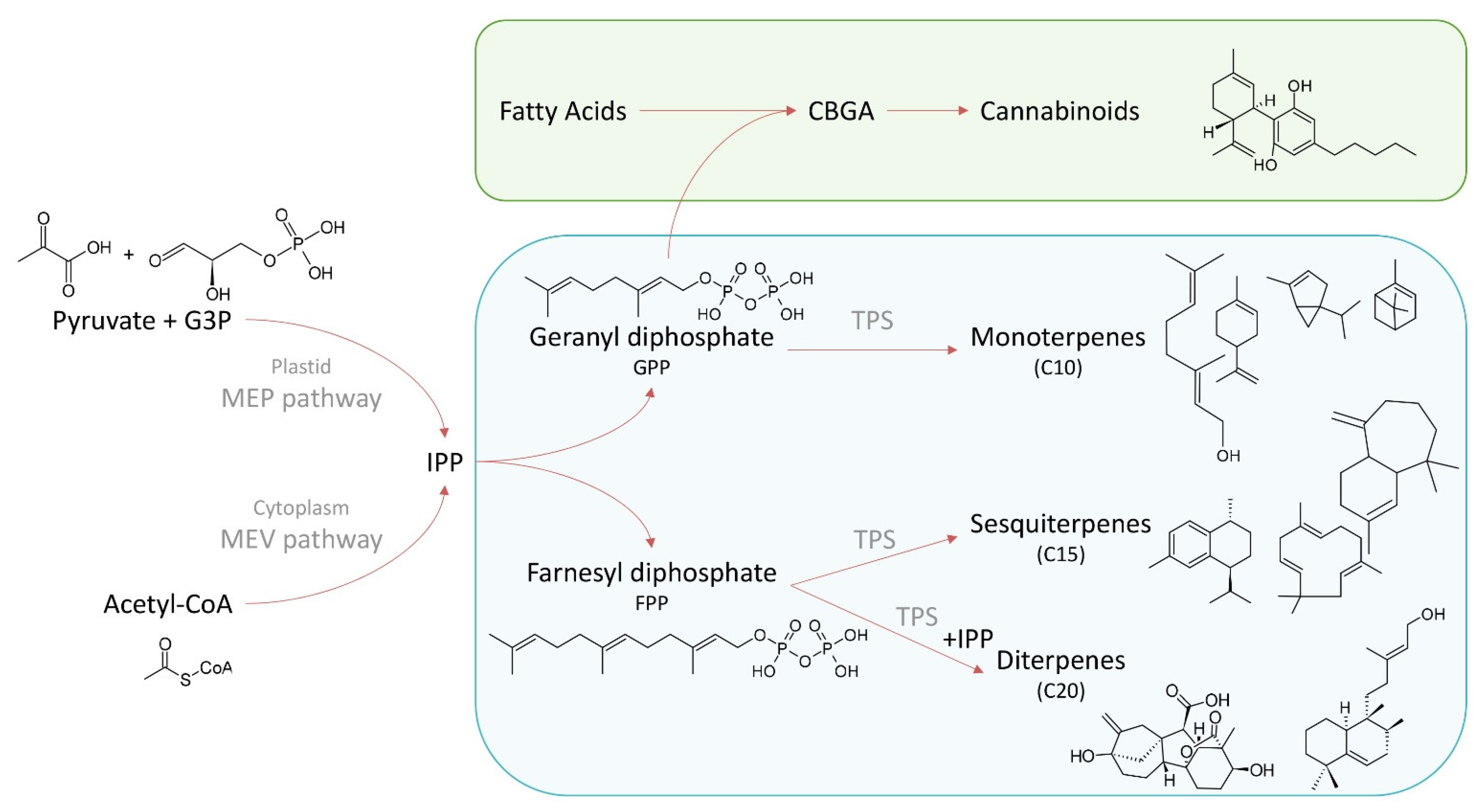{kind=link}
{kind=link}
{kind=link}
| Compound | Chemotypes | Rage of Average Concentrations Reported per Chemotype (mg/g Dry Weight) | Reference | |
|---|---|---|---|---|
| Agrospirol | I | I: | Tr–0.50 | [45] |
| Alloaromandrene | I, II, III | I: | 0.004–0.08 | [53,60] |
| II: | 0.08–0.10 | |||
| III: | 0.05–0.10 | |||
| Aromadendrene | I | I: | 0.02–0.13 | [61] |
| α-Bisabolol | I, II, III | I: | Tr–1.10 | [34,45,46,53,60,62,63,64] |
| II: | 0.57–1.22 | |||
| III: | 0.07–2.31 | |||
| α-Bisabolene | I, II, III | I: | 0.13–0.50 | [53,61] |
| II: | 0.11–0.29 | |||
| III: | 0.03–0.50 | |||
| β-Bisabolene | I, II, III | I: | 0.05–0.17 | [53] |
| II: | 0.18–0.51 | |||
| III: | 0.12–0.71 | |||
| Borneol | I, II, III | I: | 0.01–0.03 | [34,61,63,64] |
| II: | 0.05 | |||
| III: | 0.009–0.02 | |||
| α-bergamotene | I, II, III | I: | 0.024–1.18 | [34,53] |
| II: | 0.45–0.81 | |||
| III: | 0.018–0.68 | |||
| Cis-bergamotene | I, III | I: | 0.07–0.11 | [61] |
| III: | 0.21 | |||
| Trans-bergamotene | I, III | I: | 0.12–0.28 | [61] |
| III: | 0.04 | |||
| Bulnesol | I, II, III | I: | 0.10–0.50 | [34,45,53] |
| II: | 0.090–0.19 | |||
| III: | 0.070–0.49 | |||
| γ-cadinene | I, III | I: | 0.41–0.60 | [61] |
| III: | 0.02 | |||
| Camphene | I, III | I: | 0.002–0.09 | [34,60,63,64] |
| III: | 0.001–0.48 | |||
| Camphor | I | I: | 0.001–0.01 | [61,64] |
| P-Cimene | I, III | I: | 0.016 | [64] |
| III: | 0.01 | |||
| β-Caryophyllene | I, II, III | I: | 0.24–8.20 | [34,45,46,60,61,62,63,64,65] |
| II: | 0.86–3.90 | |||
| III: | 0.16–3.17 | |||
| β-Caryophyllene oxide | I, II, III | I: | 0.005–0.06 | [60,61,63] |
| II: | 0.02 | |||
| III: | 0.09 | |||
| Trans-β-caryophyllene | I, III | I: | 0.02–0.06 | [53,61] |
| III: | 0.06 | |||
| δ-3-carene | I, II, III | I: | Tr–0.60 | [45,46,61,64,65] |
| II: | Tr | |||
| III: | 0.065–0.070 | |||
| α-Cedrene | I, III | I: | 0.038 | [64] |
| III: | 0.023 | |||
| β-Citronellol | I, III | I: | 0.002 | [60,64] |
| III: | 0.001–0.003 | |||
| α-curcumene | I, III | I: | 0.008 | [60] |
| III: | 0.017 | |||
| β -Curcumene | I, II, III | I: | 0.014–0.61 | [53,60] |
| II: | 0.061–0.16 | |||
| III: | 0.016–0.09 | |||
| Cyclounatriene | I, III | I: | 0.02–0.13 | [34] |
| III: | 0.086 | |||
| Elemene | I, II | I: | Tr–2.70 | [45,65] |
| II: | Tr | |||
| γ-elemene | I, III | I: | 0.104–1.89 | [34,53,61] |
| III: | 0.04–0.068 | |||
| δ-elemene | I, III | I: | Tr–0.392 | [34] |
| III: | 0.005 | |||
| Eucalyptol | II, III | II: | 0.010–0.07 | [53,60,63] |
| III: | 0.052–0.14 | |||
| Eudesma-3,7(11)-diene | I, III | I: | Tr–0.80 | [34,61,65] |
| III: | 0.05 | |||
| Eudesmane | I, III | I: | 0.33–0.55 | [34] |
| III: | 0.04 | |||
| A-eudesmol | I, II | I: | 0.02 | [63] |
| II: | 0.26 | |||
| β-Eudesmol | I, II, III | I: | Tr–0.92 | [45,53,61,63,64] |
| II: | 0.23–0.65 | |||
| III: | 0.085–1.01 | |||
| γ-Eudesmol | I, III | I: | Tr–0.80 | [34,45,53,61] |
| II: | 0.30–0.78 | |||
| III: | 0.010–1.03 | |||
| α-farnesene | I, II, III | I: | 0.02–0.06 | [34,63] |
| II: | 0.24 | |||
| III: | 0.002 | |||
| β-farnesene | I, II, III | I: | 0.019–1.96 | [34,53,65] |
| II: | 0.73–1.6 | |||
| III: | 0.008–1.4 | |||
| Trans-β-farnesene | I, III | I: | 0.31–1.06 | [61,63] |
| II: | 0.35 | |||
| III: | 0.05 | |||
| Fenchone | I, II, III | I: | 0.005–0.03 | [60,63,64] |
| II: | 0.02 | |||
| III: | 0.007–0.008 | |||
| Fenchol | I, II, III | I: | 0.047–1.09 | [34,46,60,61,62,63,64] |
| II: | 0.09–0.31 | |||
| III: | 0.028–0.138 | |||
| Germacrene B | I, III | I: | 0.25–1.27 | [34] |
| III: | 0.34 | |||
| Geraniol | I, III | I: | 0.01 | [63,64] |
| III: | 0.004 | |||
| Geranyl Acetate | I | I: | Tr–0.70 | [46] |
| Guaiol | I, II, III | I: | Tr–1.09 | [34,45,53,61,63,65] |
| II: | 0.27–0.87 | |||
| III: | 0.010–1.21 | |||
| α-guaiene | I, III | I: | Tr–0.50 | [45,65] |
| II: | Tr | |||
| III: | Tr | |||
| δ-guaiene | I, II | I: | Tr–0.80 | [45,61,65] |
| II: | 0.8 | |||
| α-gurjunene | I | I: | 0.1–0.46 | [53] |
| Humulene | I, II, III | I: | Tr–4.00 | [45,46,53,64] |
| II: | 0.64–1.11 | |||
| III: | 0.26–0.93 | |||
| α-Humulene | I, II, III | I: | 0.09–1.93 | [34,60,62,63,65] |
| II: | 0.32–0.36 | |||
| III: | 0.14–0.27 | |||
| Isopulegol | I, II | I: | 0.02–0.04 | [63] |
| II: | 0.02 | |||
| Ledene | I, II | I: | 0.11–0.13 | [63] |
| II: | 0.05 | |||
| Limonene | I, II, III | I: | Tr–9.1 | [34,45,46,53,60,61,62,63,64] |
| II: | 0.079–1.14 | |||
| III: | 0.022–1.44 | |||
| Linalool | I, II, III | I: | Tr–3.10 | [34,45,46,53,60,61,62,63,64] |
| II: | 0.27–0.35 | |||
| III: | Tr–0.36 | |||
| Cis-linalool oxide | I, III | I: | 0.002 | [60] |
| III: | 0.005 | |||
| Trans-linalool oxide | I, III | I: | 0.002 | [60] |
| III: | 0.002 | |||
| Menthol | I, III | I: | 0.001 | [60] |
| III: | 0.001 | |||
| β-Myrcene | I, II, III | I: | 0.12–14.8 | [34,45,46,53,60,61,62,63,64,65] |
| II: | 0.20–3.02 | |||
| III: | 0.18–7.60 | |||
| Nerolidol | I, II, III | I: | 0.02 | [61] |
| III: | 0.01 | |||
| Trans-nerolidol | I, III | I: | 0.019–1.66 | [60,63,64] |
| II: | 0.09 | |||
| III: | 0.005–0.07 | |||
| β-Ocimene | I, III | I: | 0.21–1.38 | [34,53,63] |
| II: | 0.02 | |||
| III: | 0.19 | |||
| Cis-Ocimene | I, II, III | I: | 0.006–3.9 | [45,60,61,64,65] |
| II: | 1 | |||
| III: | 1 | |||
| Trans-Ocimene | I, III | I: | Tr–3.8 | [46,60,64] |
| III: | 0.007–0.01 | |||
| α-phellandrene | I, II, III | I: | Tr–0.60 | [65] |
| II: | Tr | |||
| III: | Tr | |||
| β-phellandrene | I, III | I: | Tr–2.1 | [34,65] |
| II: | 0.7 | |||
| III: | 0.097–0.50 | |||
| α-pinene | I, II, III | I: | Tr–6.70 | [34,45,46,53,60,61,62,63,64,65] |
| II: | 0.068–4.63 | |||
| III: | 0.004–1.40 | |||
| β-pinene | I, II, III | I: | Tr–2.00 | [34,45,46,53,60,61,62,63,64,65] |
| II: | 0.054–0.80 | |||
| III: | 0.001–0.50 | |||
| α-phellandrene | I, II, III | I: | 0.003–0.7 | [46,60,61] |
| II: | Tr | |||
| III: | 0.001 | |||
| 2-pinanol | I, III | I: | 0.036–0.16 | [34] |
| III: | 0.047 | |||
| Sabinene | I, III | I: | 0.005 | [60] |
| III: | 0.001 | |||
| Cis-sabinene hydrate | I, II | I: | 0.015–0.08 | [60,61,63] |
| II: | 0.003–0.03 | |||
| α-selinene | I, II, III | I: | 0.04–1.36 | [34,53,63] |
| II: | 0.26–0.65 | |||
| III: | 0.094–0.79 | |||
| β-selinene | I, II, III | I: | 0.093–0.61 | [53,63] |
| II: | 0.09–0.34 | |||
| III: | 0.10–0.22 | |||
| γ-selinene | I, II, III | I: | 0.09–0.63 | [53,61,65] |
| II: | 0.06–0.09 | |||
| III: | 0.03–0.14 | |||
| δ-selinene | I, III | I: | 0.10–0.36 | [34] |
| III: | 0.09 | |||
| Selina-3.7 (11) diene | I, II, III | I: | 0.03–1.89 | [53] |
| II: | 0.05–0.07 | |||
| III: | 0.06–0.092 | |||
| β-Sesquiphellanderene | I, II, III | I: | 0.09–0.48 | [53] |
| II: | 0.14–0.23 | |||
| III: | 0.074–0.19 | |||
| α-Terpinene | I, II, III | I: | Tr–0.10 | [45,60,64] |
| II: | Tr | |||
| III: | Tr–0.068 | |||
| γ-Terpinene | I, III | I: | 0.02–0.06 | [46,60,61,64] |
| III: | 0.01–0.06 | |||
| Terpineol | I, II, III | I: | Tr–0.70 | [45] |
| II: | 0.6 | |||
| III: | Tr | |||
| Terpinen-4-ol | I, III | I: | 0.02 | [60] |
| III: | 0.01 | |||
| α-Terpineol | I, III | I: | 0.04–0.9 | [34,46,60,62,64,65] |
| II: | 0.29 | |||
| III: | 0.11–0.22 | |||
| Terpinolene | I, II, III | I: | Tr–13.9 | [34,45,46,53,60,63,64,65] |
| II: | 0.010–3.70 | |||
| III: | 0.019–2.90 | |||
| Valencene | I, II | I: | 0.001–0.06 | [34,60,63] |
| II: | 0.01 | |||
| III: | 0.16 |
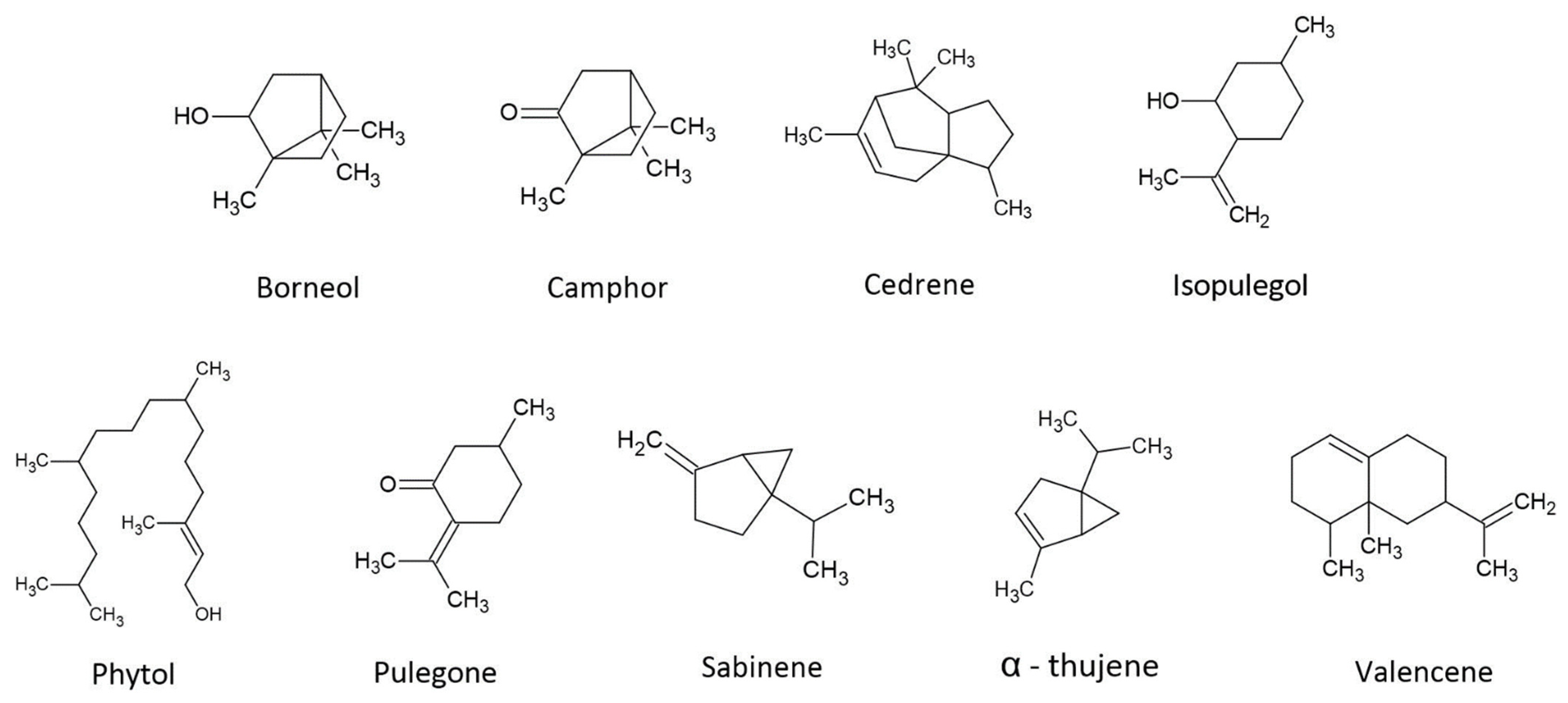
4. Potential Roles of Secondary Terpenes
4.1. Borneol
4.2. Camphor
4.3. Cedrene
4.4. Isopulegol
4.5. Phytol
4.6. Pulegone
4.7. Sabinene
4.8. Thujene
4.9. Valencene
5. Mechanism of Action for Terpenes—Pharmacologic Receptor Targets (TRPs)
6. Synergy and the Entourage Effect: Beyond Cannabinoids
7. Conclusions and Future Directions
Author Contributions
Funding
Conflicts of Interest
References
- Brand, E.J.; Zhao, Z. Cannabis in Chinese Medicine: Are Some Traditional Indications Referenced in Ancient Literature Related to Cannabinoids? Front. Pharmacol. 2017, 8, 108. [Google Scholar] [CrossRef] [Green Version]
- Ranade, A.; Tavhare, S.D.; Acharya, R.; Pawar, S.D. Cannabis (Bhanga) in Classical Text of Ayurveda: An Evidence-Based Rationale. In Cannabis/Marijuana for Healthcare; Agrawal, D.C., Kumar, R., Dhanasekaran, M., Eds.; Springer Nature: Singapore, 2022; pp. 235–251. ISBN 9789811688225. [Google Scholar]
- Bonini, S.A.; Premoli, M.; Tambaro, S.; Kumar, A.; Maccarinelli, G.; Memo, M.; Mastinu, A. Cannabis Sativa: A Comprehensive Ethnopharmacological Review of a Medicinal Plant with a Long History. J. Ethnopharmacol. 2018, 227, 300–315. [Google Scholar] [CrossRef]
- Kumar, P.; Mahato, D.K.; Kamle, M.; Borah, R.; Sharma, B.; Pandhi, S.; Tripathi, V.; Yadav, H.S.; Devi, S.; Patil, U.; et al. Pharmacological Properties, Therapeutic Potential, and Legal Status of Cannabis sativa L.: An Overview. Phytother. Res. 2021, 35, 6010–6029. [Google Scholar] [CrossRef]
- Hurgobin, B.; Tamiru-Oli, M.; Welling, M.T.; Doblin, M.S.; Bacic, A.; Whelan, J.; Lewsey, M.G. Recent Advances in Cannabis Sativa Genomics Research. New Phytol. 2021, 230, 73–89. [Google Scholar] [CrossRef]
- Monthony, A.S.; Page, S.R.; Hesami, M.; Jones, A.M.P. The Past, Present and Future of Cannabis Sativa Tissue Culture. Plants 2021, 10, 185. [Google Scholar] [CrossRef]
- Radwan, M.M.; Chandra, S.; Gul, S.; ElSohly, M.A. Cannabinoids, Phenolics, Terpenes and Alkaloids of Cannabis. Molecules 2021, 26, 2774. [Google Scholar] [CrossRef]
- Smith, T.; May, G.; Eckl, V.; Reynolds, C.M. US Sales of Herbal Supplements Increase by 8.6% in 2019. HerbalGram 2020, 54–69. [Google Scholar]
- Avins, D.; Kopf, J. New Data Show Americans Are Turning to CBD as a Cure-All for the Modern Condition. Available online: https://qz.com/1590765/survey-shows-americans-use-cbd-to-treat-anxiety-and-stress/ (accessed on 2 February 2022).
- The Brightfield Group. US CBD Market Industry Update; The Brightfield Group: Chicago, IL, USA, 2022. [Google Scholar]
- Hartsel, J.A.; Eades, J.; Hickory, B.; Makriyannis, A. Cannabis Sativa and Hemp. In Nutraceuticals; Elsevier: Amsterdam, The Netherlands, 2016; pp. 735–754. ISBN 978-0-12-802147-7. [Google Scholar]
- Oldfield, E.; Lin, F.-Y. Terpene Biosynthesis: Modularity Rules. Angew. Chem. Int. Ed. 2012, 51, 1124–1137. [Google Scholar] [CrossRef] [Green Version]
- Nuutinen, T. Medicinal Properties of Terpenes Found in Cannabis Sativa and Humulus Lupulus. Eur. J. Med. Chem. 2018, 157, 198–228. [Google Scholar] [CrossRef]
- Cox-Georgian, D.; Ramadoss, N.; Dona, C.; Basu, C. Therapeutic and Medicinal Uses of Terpenes. In Medicinal Plants: From Farm to Pharmacy; Joshee, N., Dhekney, S.A., Parajuli, P., Eds.; Springer International Publishing: Cham, Switzerland, 2019; pp. 333–359. ISBN 978-3-030-31269-5. [Google Scholar]
- Greay, S.J.; Hammer, K.A. Recent Developments in the Bioactivity of Mono- and Diterpenes: Anticancer and Antimicrobial Activity. Phytochem. Rev. 2015, 14, 1–6. [Google Scholar] [CrossRef]
- Masyita, A.; Mustika Sari, R.; Dwi Astuti, A.; Yasir, B.; Rahma Rumata, N.; Emran, T.B.; Nainu, F.; Simal-Gandara, J. Terpenes and Terpenoids as Main Bioactive Compounds of Essential Oils, Their Roles in Human Health and Potential Application as Natural Food Preservatives. Food Chem. X 2022, 13, 100217. [Google Scholar] [CrossRef]
- Koziol, A.; Stryjewska, A.; Librowski, T.; Salat, K.; Gawel, M.; Moniczewski, A.; Lochynski, S. An Overview of the Pharmacological Properties and Potential Applications of Natural Monoterpenes. Mini Rev. Med. Chem. 2014, 14, 1156–1168. [Google Scholar] [CrossRef]
- Lasekan, O.; Abbas, K.A. Distinctive Exotic Flavor and Aroma Compounds of Some Exotic Tropical Fruits and Berries: A Review. Crit. Rev. Food Sci. Nutr. 2012, 52, 726–735. [Google Scholar] [CrossRef]
- Marais, J. Terpenes in the Aroma of Grapes and Wines: A Review. SAJEV 2017, 4, 49–58. [Google Scholar] [CrossRef]
- Mele, M.A.; Kang, H.-M.; Lee, Y.-T.; Islam, M.Z. Grape Terpenoids: Flavor Importance, Genetic Regulation, and Future Potential. Crit. Rev. Food Sci. Nutr. 2021, 61, 1429–1447. [Google Scholar] [CrossRef]
- Roell, M.-S. Terpenes in Cannabis: Solving the Puzzle of How to Predict Taste and Smell. Plant Physiol. 2020, 184, 8–9. [Google Scholar] [CrossRef]
- McPartland, J.M.; Russo, E.B. Cannabis and Cannabis Extracts. J. Cannabis Ther. 2001, 1, 103–132. [Google Scholar] [CrossRef]
- Russo, E.B. Taming THC: Potential Cannabis Synergy and Phytocannabinoid-Terpenoid Entourage Effects. Br. J. Pharmacol. 2011, 163, 1344–1364. [Google Scholar] [CrossRef]
- Efferth, T.; Koch, E. Complex Interactions between Phytochemicals. The Multi-Target Therapeutic Concept of Phytotherapy. Curr. Drug Targets 2011, 12, 122–132. [Google Scholar] [CrossRef]
- Wagner, H.; Ulrich-Merzenich, G. Synergy Research: Approaching a New Generation of Phytopharmaceuticals. Phytomedicine 2009, 16, 97–110. [Google Scholar] [CrossRef]
- Ben-Shabat, S.; Fride, E.; Sheskin, T.; Tamiri, T.; Rhee, M.-H.; Vogel, Z.; Bisogno, T.; De Petrocellis, L.; Di Marzo, V.; Mechoulam, R. An Entourage Effect: Inactive Endogenous Fatty Acid Glycerol Esters Enhance 2-Arachidonoyl-Glycerol Cannabinoid Activity. Eur. J. Pharmacol. 1998, 353, 23–31. [Google Scholar] [CrossRef]
- Rohmer, M. From Molecular Fossils of Bacterial Hopanoids to the Formation of Isoprene Units: Discovery and Elucidation of the Methylerythritol Phosphate Pathway. Lipids 2008, 43, 1095–1107. [Google Scholar] [CrossRef]
- Miziorko, H.M. Enzymes of the Mevalonate Pathway of Isoprenoid Biosynthesis. Arch. Biochem. Biophys. 2011, 505, 131–143. [Google Scholar] [CrossRef] [Green Version]
- Jiang, S.-Y.; Jin, J.; Sarojam, R.; Ramachandran, S. A Comprehensive Survey on the Terpene Synthase Gene Family Provides New Insight into Its Evolutionary Patterns. Genome Biol. Evol. 2019, 11, 2078–2098. [Google Scholar] [CrossRef] [Green Version]
- Degenhardt, F.; Stehle, F.; Kayser, O. Chapter 2—The Biosynthesis of Cannabinoids. In Handbook of Cannabis and Related Pathologies; Preedy, V.R., Ed.; Academic Press: San Diego, CA, USA, 2017; pp. 13–23. ISBN 978-0-12-800756-3. [Google Scholar]
- Hanuš, L.O.; Hod, Y. Terpenes/Terpenoids in Cannabis: Are They Important? MCA 2020, 3, 25–60. [Google Scholar] [CrossRef]
- McGarvey, D.J.; Croteau, R. Terpenoid Metabolism. Plant Cell 1995, 7, 1015–1026. [Google Scholar] [CrossRef] [Green Version]
- Chen, F.; Tholl, D.; Bohlmann, J.; Pichersky, E. The Family of Terpene Synthases in Plants: A Mid-Size Family of Genes for Specialized Metabolism That Is Highly Diversified throughout the Kingdom. Plant J. 2011, 66, 212–229. [Google Scholar] [CrossRef]
- Booth, J.K.; Yuen, M.M.S.; Jancsik, S.; Madilao, L.L.; Page, J.E.; Bohlmann, J. Terpene Synthases and Terpene Variation in Cannabis Sativa. Plant Physiol. 2020, 184, 130–147. [Google Scholar] [CrossRef]
- Booth, J.K.; Page, J.E.; Bohlmann, J. Terpene Synthases from Cannabis Sativa. PLoS ONE 2017, 12, e0173911. [Google Scholar] [CrossRef] [Green Version]
- Livingston, S.J.; Quilichini, T.D.; Booth, J.K.; Wong, D.C.J.; Rensing, K.H.; Laflamme-Yonkman, J.; Castellarin, S.D.; Bohlmann, J.; Page, J.E.; Samuels, A.L. Cannabis Glandular Trichomes Alter Morphology and Metabolite Content during Flower Maturation. Plant J. 2020, 101, 37–56. [Google Scholar] [CrossRef]
- Zager, J.J.; Lange, I.; Srividya, N.; Smith, A.; Lange, B.M. Gene Networks Underlying Cannabinoid and Terpenoid Accumulation in Cannabis. Plant Physiol. 2019, 180, 1877–1897. [Google Scholar] [CrossRef]
- Günnewich, N.; Page, J.E.; Köllner, T.G.; Degenhardt, J.; Kutchan, T.M. Functional Expression and Characterization of Trichome-Specific (-)-Limonene Synthase and (+)-α-Pinene Synthase from Cannabis Sativa. Nat. Prod. Commun. 2007, 2, 223–232. [Google Scholar] [CrossRef] [Green Version]
- Watts, S.; McElroy, M.; Migicovsky, Z.; Maassen, H.; van Velzen, R.; Myles, S. Cannabis Labelling Is Associated with Genetic Variation in Terpene Synthase Genes. Nat. Plants 2021, 7, 1330–1334. [Google Scholar] [CrossRef]
- Ross, S.A.; ElSohly, M.A. The Volatile Oil Composition of Fresh and Air-Dried Buds of Cannabis Sativa. J. Nat. Prod. 1996, 59, 49–51. [Google Scholar] [CrossRef]
- Tholl, D. Terpene Synthases and the Regulation, Diversity and Biological Roles of Terpene Metabolism. Curr. Opin. Plant Biol. 2006, 9, 297–304. [Google Scholar] [CrossRef]
- Lewis, M.A.; Russo, E.B.; Smith, K.M. Pharmacological Foundations of Cannabis Chemovars. Planta Med. 2018, 84, 225–233. [Google Scholar] [CrossRef]
- Brown, A.K.; Xia, Z.; Bulloch, P.; Idowu, I.; Francisco, O.; Stetefeld, J.; Stout, J.; Zimmer, J.; Marvin, C.; Letcher, R.J.; et al. Validated Quantitative Cannabis Profiling for Canadian Regulatory Compliance—Cannabinoids, Aflatoxins, and Terpenes. Anal. Chim. Acta 2019, 1088, 79–88. [Google Scholar] [CrossRef]
- Pacifico, D.; Miselli, F.; Carboni, A.; Moschella, A.; Mandolino, G. Time Course of Cannabinoid Accumulation and Chemotype Development during the Growth of Cannabis sativa L. Euphytica 2008, 160, 231–240. [Google Scholar] [CrossRef]
- Fischedick, J.T.; Hazekamp, A.; Erkelens, T.; Choi, Y.H.; Verpoorte, R. Metabolic Fingerprinting of Cannabis Sativa L., Cannabinoids and Terpenoids for Chemotaxonomic and Drug Standardization Purposes. Phytochemistry 2010, 71, 2058–2073. [Google Scholar] [CrossRef]
- Fischedick, J.T. Identification of Terpenoid Chemotypes Among High (−)-Trans-Δ9- Tetrahydrocannabinol-Producing Cannabis sativa L. Cultivars. Cannabis Cannabinoid Res. 2017, 2, 34–47. [Google Scholar] [CrossRef] [Green Version]
- Richins, R.D.; Rodriguez-Uribe, L.; Lowe, K.; Ferral, R.; O’Connell, M.A. Accumulation of Bioactive Metabolites in Cultivated Medical Cannabis. PLoS ONE 2018, 13, e0201119. [Google Scholar] [CrossRef]
- Reimann-Philipp, U.; Speck, M.; Orser, C.; Johnson, S.; Hilyard, A.; Turner, H.; Stokes, A.J.; Small-Howard, A.L. Cannabis Chemovar Nomenclature Misrepresents Chemical and Genetic Diversity; Survey of Variations in Chemical Profiles and Genetic Markers in Nevada Medical Cannabis Samples. Cannabis Cannabinoid Res. 2020, 5, 215–230. [Google Scholar] [CrossRef] [Green Version]
- Birenboim, M.; Chalupowicz, D.; Maurer, D.; Barel, S.; Chen, Y.; Fallik, E.; Paz-Kagan, T.; Rapaport, T.; Sadeh, A.; Kengisbuch, D.; et al. Multivariate Classification of Cannabis Chemovars Based on Their Terpene and Cannabinoid Profiles. Phytochemistry 2022, 200, 113215. [Google Scholar] [CrossRef]
- Lemay, J.; Zheng, Y.; Scott-Dupree, C. Factors Influencing the Efficacy of Biological Control Agents Used to Manage Insect Pests in Indoor Cannabis (Cannabis sativa) Cultivation. Front. Agron. 2022, 4, 795989. [Google Scholar] [CrossRef]
- Whipker, B.E. Put Your Fertilizer Program to the Test. Available online: https://www.cannabisbusinesstimes.com/article/cultivation-matters-ncsu-north-carolina-cannabis-fertilization-program-management-npk-nitrogen-phosphorus-potassium/ (accessed on 28 September 2022).
- Hawley, D.; Graham, T.; Stasiak, M.; Dixon, M. Improving Cannabis Bud Quality and Yield with Subcanopy Lighting. HortScience 2018, 53, 1593–1599. [Google Scholar] [CrossRef]
- Aizpurua-Olaizola, O.; Soydaner, U.; Öztürk, E.; Schibano, D.; Simsir, Y.; Navarro, P.; Etxebarria, N.; Usobiaga, A. Evolution of the Cannabinoid and Terpene Content during the Growth of Cannabis Sativa Plants from Different Chemotypes. J. Nat. Prod. 2016, 79, 324–331. [Google Scholar] [CrossRef]
- AL Ubeed, H.M.S.; Wills, R.B.H.; Chandrapala, J. Post-Harvest Operations to Generate High-Quality Medicinal Cannabis Products: A Systemic Review. Molecules 2022, 27, 1719. [Google Scholar] [CrossRef]
- Pagnani, G.; Pellegrini, M.; Galieni, A.; D’Egidio, S.; Matteucci, F.; Ricci, A.; Stagnari, F.; Sergi, M.; Lo Sterzo, C.; Pisante, M.; et al. Plant Growth-Promoting Rhizobacteria (PGPR) in Cannabis Sativa ‘Finola’ Cultivation: An Alternative Fertilization Strategy to Improve Plant Growth and Quality Characteristics. Ind. Crops Prod. 2018, 123, 75–83. [Google Scholar] [CrossRef]
- Hudson, J.R. Humulones, Lupulones and Other Constituents of Hops. In Modern Methods of Plant Analysis/Moderne Methoden der Pflanzenanalyse; Linskens, H.F., Tracey, M.V., Beiss, U., Bendall, F., Björk, W., Bohlmann, F., Boman, H.G., Braun, R., Heinen, W., Hesse, M., et al., Eds.; Springer: Berlin/Heidelberg, Germany, 1963; pp. 135–154. ISBN 978-3-642-94878-7. [Google Scholar]
- Desaulniers Brousseau, V.; Wu, B.-S.; MacPherson, S.; Morello, V.; Lefsrud, M. Cannabinoids and Terpenes: How Production of Photo-Protectants Can Be Manipulated to Enhance Cannabis sativa L. Phytochemistry. Front. Plant Sci. 2021, 12, 620021. [Google Scholar] [CrossRef]
- Lydon, J.; Teramura, A.H.; Coffman, C.B. UV-B RADIATION EFFECTS ON PHOTOSYNTHESIS, GROWTH and CANNABINOID PRODUCTION OF TWO Cannabis sativa CHEMOTYPES. Photochem. Photobiol. 1987, 46, 201–206. [Google Scholar] [CrossRef] [PubMed]
- Eichhorn Bilodeau, S.; Wu, B.-S.; Rufyikiri, A.-S.; MacPherson, S.; Lefsrud, M. An Update on Plant Photobiology and Implications for Cannabis Production. Front. Plant Sci. 2019, 10, 296. [Google Scholar] [CrossRef] [PubMed]
- Milay, L.; Berman, P.; Shapira, A.; Guberman, O.; Meiri, D. Metabolic Profiling of Cannabis Secondary Metabolites for Evaluation of Optimal Postharvest Storage Conditions. Front. Plant Sci. 2020, 11, 583605. [Google Scholar] [CrossRef] [PubMed]
- Hazekamp, A.; Tejkalová, K.; Papadimitriou, S. Cannabis: From Cultivar to Chemovar II—A Metabolomics Approach to Cannabis Classification. Cannabis Cannabinoid Res. 2016, 1, 202–215. [Google Scholar] [CrossRef]
- Sexton, M.; Shelton, K.; Haley, P.; West, M. Evaluation of Cannabinoid and Terpenoid Content: Cannabis Flower Compared to Supercritical CO2 Concentrate. Planta Med. 2018, 84, 234–241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, D.; Dai, K.; Xie, Z.; Chen, J. Secondary Metabolites Profiled in Cannabis Inflorescences, Leaves, Stem Barks, and Roots for Medicinal Purposes. Sci. Rep. 2020, 10, 3309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dei Cas, M.; Arnoldi, S.; Monguzzi, L.; Casagni, E.; Morano, C.; Vieira de Manincor, E.; Bolchi, C.; Pallavicini, M.; Gambaro, V.; Roda, G. Characterization of Chemotype-Dependent Terpenoids Profile in Cannabis by Headspace Gas-Chromatography Coupled to Time-of-Flight Mass Spectrometry. J. Pharm. Biomed. Anal. 2021, 203, 114180. [Google Scholar] [CrossRef]
- Hazekamp, A.; Fischedick, J.T. Cannabis—From Cultivar to Chemovar. Drug Test. Anal. 2012, 4, 660–667. [Google Scholar] [CrossRef]
- Russo, E.B.; Marcu, J. Cannabis Pharmacology: The Usual Suspects and a Few Promising Leads. In Advances in Pharmacology; Elsevier: Amsterdam, The Netherlands, 2017; Volume 80, pp. 67–134. ISBN 978-0-12-811232-8. [Google Scholar]
- Baron, E.P. Medicinal Properties of Cannabinoids, Terpenes, and Flavonoids in Cannabis, and Benefits in Migraine, Headache, and Pain: An Update on Current Evidence and Cannabis Science. Headache J. Head Face Pain 2018, 58, 1139–1186. [Google Scholar] [CrossRef]
- Sommano, S.R.; Chittasupho, C.; Ruksiriwanich, W.; Jantrawut, P. The Cannabis Terpenes. Molecules 2020, 25, 5792. [Google Scholar] [CrossRef]
- Weston-Green, K.; Clunas, H.; Jimenez Naranjo, C. A Review of the Potential Use of Pinene and Linalool as Terpene-Based Medicines for Brain Health: Discovering Novel Therapeutics in the Flavours and Fragrances of Cannabis. Front. Psychiatry 2021, 12, 583211. [Google Scholar] [CrossRef]
- Lowe, H.; Steele, B.; Bryant, J.; Toyang, N.; Ngwa, W. Non-Cannabinoid Metabolites of Cannabis sativa L. with Therapeutic Potential. Plants 2021, 10, 400. [Google Scholar] [CrossRef]
- Pattnaik, F.; Nanda, S.; Mohanty, S.; Dalai, A.K.; Kumar, V.; Ponnusamy, S.K.; Naik, S. Cannabis: Chemistry, Extraction and Therapeutic Applications. Chemosphere 2022, 289, 133012. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.-P.; Feng, J.-F.; Hu, K.-L. Progress in regulation effect of aromatic refreshing traditional Chinese medicine on BBB permeability and its mechanism. Zhongguo Zhong Yao Za Zhi 2014, 39, 949–954. [Google Scholar]
- Zhang, Q.-L.; Fu, B.M.; Zhang, Z.-J. Borneol, a Novel Agent That Improves Central Nervous System Drug Delivery by Enhancing Blood–Brain Barrier Permeability. Drug Deliv. 2017, 24, 1037–1044. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiong, Z.-Y.; Xiao, F.-M.; Xu, X.; Wu, Y.-F.; Jiang, X.-M. Studies on pharmacological activity of borneol. Zhongguo Zhong Yao Za Zhi 2013, 38, 786–790. [Google Scholar] [PubMed]
- da Silva Almeida, J.R.G.; Souza, G.R.; Silva, J.C.; de Lima Saraiva, S.R.G.; de Oliveira Júnior, R.G.; de Souza Siqueira Quintans, J.; de Souza Siqueira Barreto, R.; Bonjardim, L.R.; de Holanda Cavalcanti, S.C.; Junior, L.J.Q. Borneol, a Bicyclic Monoterpene Alcohol, Reduces Nociceptive Behavior and Inflammatory Response in Mice. Sci. World J. 2013, 2013, e808460. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.-H.; Sun, X.-P.; Zhang, Y.-Q.; Wang, N.-S. The Antithrombotic Effect of Borneol Related to Its Anticoagulant Property. Am. J. Chin. Med. 2008, 36, 719–727. [Google Scholar] [CrossRef]
- Liu, R.; Zhang, L.; Lan, X.; Li, L.; Zhang, T.-T.; Sun, J.-H.; Du, G.-H. Protection by Borneol on Cortical Neurons against Oxygen-Glucose Deprivation/Reperfusion: Involvement of Anti-Oxidation and Anti-Inflammation through Nuclear Transcription Factor ΚappaB Signaling Pathway. Neuroscience 2011, 176, 408–419. [Google Scholar] [CrossRef]
- Chen, X.; Lin, Z.; Liu, A.; Ye, J.; Luo, Y.; Luo, Y.; Mao, X.; Liu, P.; Pi, R. The Orally Combined Neuroprotective Effects of Sodium Ferulate and Borneol against Transient Global Ischaemia in C57 BL/6J Mice. J. Pharm. Pharmacol. 2010, 62, 915–923. [Google Scholar] [CrossRef]
- Horvathova, E.; Mastihubova, M.; Karnisova Potocka, E.; Kis, P.; Galova, E.; Sevcovicova, A.; Klapakova, M.; Hunakova, L.; Mastihuba, V. Comparative Study of Relationship between Structure of Phenylethanoid Glycopyranosides and Their Activities Using Cell-Free Assays and Human Cells Cultured in Vitro. Toxicol. Vitr. 2019, 61, 104646. [Google Scholar] [CrossRef]
- Corrêa, P.R.C.; Miranda, R.R.S.; Duarte, L.P.; Silva, G.D.F.; Filho, S.A.V.; Okuma, A.A.; Carazza, F.; Morgado-Díaz, J.A.; Pinge-Filho, P.; Yamauchi, L.M.; et al. Antimicrobial Activity of Synthetic Bornyl Benzoates against Trypanosoma Cruzi. Pathog. Glob. Health 2012, 106, 107–112. [Google Scholar] [CrossRef] [Green Version]
- Vasconcelos, R.M.C.; Leite, F.C.; Leite, J.A.; Rodrigues Mascarenhas, S.; Rodrigues, L.C.; Piuvezam, M.R. Synthesis, Acute Toxicity and Anti-Inflammatory Effect of Bornyl Salicylate, a Salicylic Acid Derivative. Immunopharmacol. Immunotoxicol. 2012, 34, 1028–1038. [Google Scholar] [CrossRef]
- PubChem Camphor. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/2537 (accessed on 4 October 2022).
- Chen, W.; Vermaak, I.; Viljoen, A. Camphor—A Fumigant during the Black Death and a Coveted Fragrant Wood in Ancient Egypt and Babylon—A Review. Molecules 2013, 18, 5434–5454. [Google Scholar] [CrossRef] [Green Version]
- Hamidpour, R.; Hamidpour, S.; Hamidpour, M.; Shahlari, M. Camphor (Cinnamomum Camphora), a Traditional Remedy with the History of Treating Several Diseases. Int. J. Case Rep. Images (IJCRI) 2013, 4, 86–89. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Zhang, K.; Zhang, K.; Zhang, J.; Fu, J.; Li, J.; Wang, G.; Qiu, Z.; Wang, X.; Li, J. Antibacterial Activity of Cinnamomum Camphora Essential Oil on Escherichia Coli During Planktonic Growth and Biofilm Formation. Front. Microbiol. 2020, 11, 561002. [Google Scholar] [CrossRef]
- Zarubaev, V.V.; Garshinina, A.V.; Tretiak, T.S.; Fedorova, V.A.; Shtro, A.A.; Sokolova, A.S.; Yarovaya, O.I.; Salakhutdinov, N.F. Broad Range of Inhibiting Action of Novel Camphor-Based Compound with Anti-Hemagglutinin Activity against Influenza Viruses in Vitro and in Vivo. Antivir. Res. 2015, 120, 126–133. [Google Scholar] [CrossRef]
- Kumar, N.; Nepali, K.; Sapra, S.; Bijjem, K.R.V.; Kumar, R.; Suri, O.P.; Dhar, K.L. Effect of Nitrogen Insertion on the Antitussive Properties of Menthol and Camphor. Med. Chem. Res. 2012, 21, 531–537. [Google Scholar] [CrossRef]
- Nikolić, B.; Vasilijević, B.; Mitić-Ćulafić, D.; Vuković-Gačić, B.; Knežević-Vukćević, J. Comparative Study of Genotoxic, Antigenotoxic and Cytotoxic Activities of Monoterpenes Camphor, Eucalyptol and Thujone in Bacteria and Mammalian Cells. Chem.-Biol. Interact. 2015, 242, 263–271. [Google Scholar] [CrossRef]
- Moayedi, Y.; Greenberg, S.A.; Jenkins, B.A.; Marshall, K.L.; Dimitrov, L.V.; Nelson, A.M.; Owens, D.M.; Lumpkin, E.A. Camphor White Oil Induces Tumor Regression through Cytotoxic T Cell-Dependent Mechanisms. Mol. Carcinog. 2019, 58, 722–734. [Google Scholar] [CrossRef]
- Shata, F.Y.H.; Eldebaky, H.A.A. Effects of Camphor on Hepatic Enzymes, Steroids and Antioxidant Capacity of Male Rats Intoxicated with Atrazine. Middle-East J. of Sci. Res. 2014, 22, 553–560. [Google Scholar]
- Drikvandi, P.; Bahramikia, S.; Alirezaei, M. Modulation of the Antioxidant Defense System in Liver, Kidney, and Pancreas Tissues of Alloxan-Induced Diabetic Rats by Camphor. J. Food Biochem. 2020, 44, e13527. [Google Scholar] [CrossRef]
- Poudel, D.K.; Rokaya, A.; Ojha, P.K.; Timsina, S.; Satyal, R.; Dosoky, N.S.; Satyal, P.; Setzer, W.N. The Chemical Profiling of Essential Oils from Different Tissues of Cinnamomum Camphora L. and Their Antimicrobial Activities. Molecules 2021, 26, 5132. [Google Scholar] [CrossRef]
- Baker, B.P.; Grant, J.A.; Malakar-Kuenen, R. Cedarwood Oil Profile. Cornell Cooperative Extension Integrated Pest Management Program; Cornell University: Ithaca, NY, USA, 2018; pp. 1–8. Available online: https://ecommons.cornell.edu/handle/1813/56116 (accessed on 15 September 2022).
- Johnston, W.H.; Karchesy, J.J.; Constantine, G.H.; Craig, A.M. Antimicrobial Activity of Some Pacific Northwest Woods against Anaerobic Bacteria and Yeast. Phytother. Res. 2001, 15, 586–588. [Google Scholar] [CrossRef] [PubMed]
- National Toxicology Program (NTP). NTP Technical Report on the Toxicity Studies of Cedarwood Oil (Virginia) (CASRN 8000-27-9) Administered Dermally to F344/N Rats and B6C3F1/N Mice; National Toxicology Program: Research Triangle Park, NC, USA, 2019; p. 86.
- Al-Daghri, N.M.; Alokail, M.S.; Alkharfy, K.M.; Mohammed, A.K.; Abd-Alrahman, S.H.; Yakout, S.M.; Amer, O.E.; Krishnaswamy, S. Fenugreek Extract as an Inducer of Cellular Death via Autophagy in Human T Lymphoma Jurkat Cells. BMC Complement. Altern. Med. 2012, 12, 202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Rahmah, A.N.; Mostafa, A.A.; Abdel-Megeed, A.; Yakout, S.M.; Hussein, S.A. Fungicidal Activities of Certain Methanolic Plant Extracts against Tomato Phytopathogenic Fungi. Afr. J. Microbiol. Res. 2013, 7, 517–524. [Google Scholar]
- Su, Y.-C.; Hsu, K.-P.; Wang, E.I.-C.; Ho, C.-L. Composition, Anticancer, and Antimicrobial Activities in Vitro of the Heartwood Essential Oil of Cunninghamia Lanceolata Var. Konishii from Taiwan. Nat. Prod. Commun. 2012, 7, 1245–1247. [Google Scholar] [CrossRef] [Green Version]
- Kim, T.H.; Yoo, S.D.; Lee, H.S.; Lee, K.M.; Seok, S.H.; Kim, M.G.; Jung, B.H.; Kim, M.G.; Shin, B.S. In Vivo Absorption and Disposition of α-Cedrene, a Sesquiterpene Constituent of Cedarwood Oil, in Female and Male Rats. Drug Metab. Pharmacokinet. 2015, 30, 168–173. [Google Scholar] [CrossRef]
- Tong, T.; Yu, R.; Park, T. α-Cedrene Protects Rodents from High-Fat Diet-Induced Adiposity via Adenylyl Cyclase 3. Int. J. Obes. 2019, 43, 202–216. [Google Scholar] [CrossRef]
- Haque, A.N.M.A.; Remadevi, R.; Naebe, M. Lemongrass (Cymbopogon): A Review on Its Structure, Properties, Applications and Recent Developments. Cellulose 2018, 25, 5455–5477. [Google Scholar] [CrossRef]
- Spencer, J.S.; Dowd, E.; Faas, W.; Company, A.T. The Genuineness of Two Mint Essential Oils. Perfum. Flavorist 1997, 22, 37–45. [Google Scholar]
- Rajeswara Rao, B.R.; Kaul, P.N.; Syamasundar, K.V.; Ramesh, S. Comparative Composition of Decanted and Recovered Essential Oils of Eucalyptus Citriodora Hook. Flavour Fragr. J. 2003, 18, 133–135. [Google Scholar] [CrossRef]
- Leafly Cannabis Glossary. Available online: https://www.leafly.com/learn/cannabis-glossary (accessed on 15 September 2022).
- Silva, M.I.G.; de Aquino Neto, M.R.; Teixeira Neto, P.F.; Moura, B.A.; do Amaral, J.F.; de Sousa, D.P.; Vasconcelos, S.M.M.; de Sousa, F.C.F. Central Nervous System Activity of Acute Administration of Isopulegol in Mice. Pharmacol. Biochem. Behav. 2007, 88, 141–147. [Google Scholar] [CrossRef]
- Silva, M.I.G.; Silva, M.A.G.; de Aquino Neto, M.R.; Moura, B.A.; de Sousa, H.L.; de Lavor, E.P.H.; de Vasconcelos, P.F.; Macêdo, D.S.; de Sousa, D.P.; Vasconcelos, S.M.M.; et al. Effects of Isopulegol on Pentylenetetrazol-Induced Convulsions in Mice: Possible Involvement of GABAergic System and Antioxidant Activity. Fitoterapia 2009, 80, 506–513. [Google Scholar] [CrossRef] [PubMed]
- Silva, M.I.G.; Moura, B.A.; de Aquino Neto, M.R.; da Rocha Tomé, A.; Rocha, N.F.M.; de Carvalho, A.M.R.; Macêdo, D.S.; Vasconcelos, S.M.M.; de Sousa, D.P.; de Barros Viana, G.S.; et al. Gastroprotective Activity of Isopulegol on Experimentally Induced Gastric Lesions in Mice: Investigation of Possible Mechanisms of Action. Naunyn-Schmied Arch Pharm. 2009, 380, 233–245. [Google Scholar] [CrossRef] [PubMed]
- Bounihi, A.; Hajjaj, G.; Alnamer, R.; Cherrah, Y.; Zellou, A. In Vivo Potential Anti-Inflammatory Activity of Melissa Officinalis L. Essential Oil. Adv. Pharmacol. Pharm. Sci. 2013, 2013, e101759. [Google Scholar] [CrossRef]
- Guo, X.; Ho, C.-T.; Schwab, W.; Wan, X. Aroma Profiles of Green Tea Made with Fresh Tea Leaves Plucked in Summer. Food Chem. 2021, 363, 130328. [Google Scholar] [CrossRef]
- Hossain, M.A.; Al-Hdhrami, S.S.; Weli, A.M.; Al-Riyami, Q.; Al-Sabahi, J.N. Isolation, Fractionation and Identification of Chemical Constituents from the Leaves Crude Extracts of Mentha Piperita L Grown in Sultanate of Oman. Asian Pac. J. Trop. Biomed. 2014, 4, S368–S372. [Google Scholar] [CrossRef] [Green Version]
- Kim, C.-W.; Lee, H.J.; Jung, J.H.; Kim, Y.H.; Jung, D.-B.; Sohn, E.J.; Lee, J.H.; Woo, H.J.; Baek, N.-I.; Kim, Y.C.; et al. Activation of Caspase-9/3 and Inhibition of Epithelial Mesenchymal Transition Are Critically Involved in Antitumor Effect of Phytol in Hepatocellular Carcinoma Cells. Phytother. Res. 2015, 29, 1026–1031. [Google Scholar] [CrossRef]
- Santos, C.C.d.M.P.; Salvadori, M.S.; Mota, V.G.; Costa, L.M.; de Almeida, A.A.C.; de Oliveira, G.A.L.; Costa, J.P.; de Sousa, D.P.; de Freitas, R.M.; de Almeida, R.N. Antinociceptive and Antioxidant Activities of Phytol In Vivo and In Vitro Models. Neurosci. J. 2013, 2013, 949452. [Google Scholar] [CrossRef] [Green Version]
- Silva, R.O.; Sousa, F.B.M.; Damasceno, S.R.B.; Carvalho, N.S.; Silva, V.G.; Oliveira, F.R.M.A.; Sousa, D.P.; Aragão, K.S.; Barbosa, A.L.R.; Freitas, R.M.; et al. Phytol, a Diterpene Alcohol, Inhibits the Inflammatory Response by Reducing Cytokine Production and Oxidative Stress. Fundam. Clin. Pharmacol. 2014, 28, 455–464. [Google Scholar] [CrossRef] [Green Version]
- Pejin, B.; Kojic, V.; Bogdanovic, G. An Insight into the Cytotoxic Activity of Phytol at in Vitro Conditions. Nat. Prod. Res. 2014, 28, 2053–2056. [Google Scholar] [CrossRef] [PubMed]
- Costa, J.P.; de Oliveira, G.A.L.; de Almeida, A.A.C.; Islam, M.T.; de Sousa, D.P.; de Freitas, R.M. Anxiolytic-like Effects of Phytol: Possible Involvement of GABAergic Transmission. Brain Res. 2014, 1547, 34–42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Costa, J.P.; Ferreira, P.B.; De Sousa, D.P.; Jordan, J.; Freitas, R.M. Anticonvulsant Effect of Phytol in a Pilocarpine Model in Mice. Neurosci. Lett. 2012, 523, 115–118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moniruzzaman, M.; Mannan, M.A.; Hossen Khan, M.F.; Abir, A.B.; Afroze, M. The Leaves of Crataeva Nurvala Buch-Ham. Modulate Locomotor and Anxiety Behaviors Possibly through GABAergic System. BMC Complement. Altern. Med. 2018, 18, 283. [Google Scholar] [CrossRef] [PubMed]
- de Moraes, J.; de Oliveira, R.N.; Costa, J.P.; Junior, A.L.G.; de Sousa, D.P.; Freitas, R.M.; Allegretti, S.M.; Pinto, P.L.S. Phytol, a Diterpene Alcohol from Chlorophyll, as a Drug against Neglected Tropical Disease Schistosomiasis Mansoni. PLOS Negl. Trop. Dis. 2014, 8, e2617. [Google Scholar] [CrossRef]
- Chowdhury, R.; Ghosh, S. Phytol-Derived Novel Isoprenoid Immunostimulants. Front. Immunol. 2012, 3, 49. [Google Scholar] [CrossRef] [Green Version]
- Aachoui, Y.; Chowdhury, R.R.; Fitch, R.W.; Ghosh, S.K. Molecular Signatures of Phytol-Derived Immunostimulants in the Context of Chemokine–Cytokine Microenvironment and Enhanced Immune Response. Cell. Immunol. 2011, 271, 227–238. [Google Scholar] [CrossRef]
- Liu, C.; Srividya, N.; Parrish, A.N.; Yue, W.; Shan, M.; Wu, Q.; Lange, B.M. Morphology of Glandular Trichomes of Japanese Catnip (Schizonepeta Tenuifolia Briquet) and Developmental Dynamics of Their Secretory Activity. Phytochemistry 2018, 150, 23–30. [Google Scholar] [CrossRef]
- Farley, D.R.; Howland, V. The Natural Variation of the Pulegone Content in Various Oils of Peppermint. J. Sci. Food Agric. 1980, 31, 1143–1151. [Google Scholar] [CrossRef]
- Turner, C.E.; Elsohly, M.A.; Boeren, E.G. Constituents of Cannabis Sativa L. XVII. A Review of the Natural Constituents. J. Nat. Prod. 1980, 43, 169–234. [Google Scholar] [CrossRef]
- Formisano, C.; Rigano, D.; Napolitano, F.; Senatore, F.; Arnold, N.A.; Piozzi, F.; Rosselli, S. Volatile Constituents of Calamintha Origanifolia Boiss. Growing Wild in Lebanon. Nat. Prod. Commun. 2007, 2, 1253–1256. [Google Scholar] [CrossRef] [Green Version]
- da Silveira, N.S.; de Oliveira-Silva, G.L.; de Freitas Lamanes, B.; da Silva Prado, L.C.; Bispo-da-Silva, L.B. The Aversive, Anxiolytic-Like, and Verapamil-Sensitive Psychostimulant Effects of Pulegone. Biol. Pharm. Bull. 2014, 37, 771–778. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ortiz de Urbina, A.V.; Martín, M.L.; Montero, M.J.; Morán, A.; San Román, L. Sedating and Antipyretic Activity of the Essential Oil of Calamintha Sylvatica Subsp. Ascendens. J. Ethnopharmacol. 1989, 25, 165–171. [Google Scholar] [CrossRef]
- Božović, M.; Ragno, R. Calamintha nepeta (L.) Savi and Its Main Essential Oil Constituent Pulegone: Biological Activities and Chemistry. Molecules 2017, 22, 290. [Google Scholar] [CrossRef] [Green Version]
- Hilfiger, L.; Triaux, Z.; Marcic, C.; Héberlé, E.; Emhemmed, F.; Darbon, P.; Marchioni, E.; Petitjean, H.; Charlet, A. Anti-Hyperalgesic Properties of Menthol and Pulegone. Front. Pharmacol. 2021, 12, 753873. [Google Scholar] [CrossRef]
- Valente, J.; Zuzarte, M.; Gonçalves, M.J.; Lopes, M.C.; Cavaleiro, C.; Salgueiro, L.; Cruz, M.T. Antifungal, Antioxidant and Anti-Inflammatory Activities of Oenanthe Crocata L. Essential Oil. Food Chem. Toxicol. 2013, 62, 349–354. [Google Scholar] [CrossRef]
- Quiroga, P.R.; Asensio, C.M.; Nepote, V. Antioxidant Effects of the Monoterpenes Carvacrol, Thymol and Sabinene Hydrate on Chemical and Sensory Stability of Roasted Sunflower Seeds. J. Sci. Food Agric. 2015, 95, 471–479. [Google Scholar] [CrossRef]
- Sela, F.; Karapandzova, M.; Stefkov, G.; Cvetkovikj, I.; Kulevanova, S. Chemical Composition and Antimicrobial Activity of Essential Oils of Juniperus Excelsa Bieb. (Cupressaceae) Grown in R. Macedonia. Pharmacogn. Res. 2015, 7, 74–80. [Google Scholar] [CrossRef] [Green Version]
- Maghsoodlou, M.T.; Kazemipoor, N.; Valizadeh, J.; Falak Nezhad Seifi, M.; Rahneshan, N. Essential Oil Composition of Eucalyptus Microtheca and Eucalyptus Viminalis. Avicenna J. Phytomed. 2015, 5, 540–552. [Google Scholar]
- Johnson, S.; DeCarlo, A.; Satyal, P.; Dosoky, N.S.; Sorensen, A.; Setzer, W.N. The Chemical Composition of Single-Tree Boswellia Frereana Resin Samples. Nat. Prod. Commun. 2021, 16, 1934578X211043727. [Google Scholar] [CrossRef]
- Hao, Y.; Kang, J.; Guo, X.; Yang, R.; Chen, Y.; Li, J.; Shi, L. Comparison of Nutritional Compositions and Essential Oil Profiles of Different Parts of a Dill and Two Fennel Cultivars. Foods 2021, 10, 1784. [Google Scholar] [CrossRef]
- Aros, D.; Garrido, N.; Rivas, C.; Medel, M.; Müller, C.; Rogers, H.; Úbeda, C. Floral Scent Evaluation of Three Cut Flowers Through Sensorial and Gas Chromatography Analysis. Agronomy 2020, 10, 131. [Google Scholar] [CrossRef] [Green Version]
- Verghese, J.; Joy, M.T.; Retamar, J.A.; Malinskas, G.G.; Catalán, C.A.N.; Gros, E.G. A Fresh Look at the Constituents of Indian Olibanum Oil. Flavour Fragr. J. 1987, 2, 99–102. [Google Scholar] [CrossRef]
- Ali, N.A.A.; Wurster, M.; Arnold, N.; Teichert, A.; Schmidt, J.; Lindequist, U.; Wessjohann, L. Chemical Composition and Biological Activities of Essential Oils from the Oleogum Resins of Three Endemic Soqotraen Boswellia Species. Rec. Nat. Prod. 2008, 8, 6–12. [Google Scholar]
- Siddiqui, M.Z. Boswellia Serrata, a Potential Antiinflammatory Agent: An Overview. Indian J. Pharm. Sci. 2011, 73, 255–261. [Google Scholar] [CrossRef]
- Sadhasivam, S.; Palanivel, S.; Ghosh, S. Synergistic Antimicrobial Activity of Boswellia Serrata Roxb. Ex Colebr. (Burseraceae) Essential Oil with Various Azoles against Pathogens Associated with Skin, Scalp and Nail Infections. Lett. Appl. Microbiol. 2016, 63, 495–501. [Google Scholar] [CrossRef]
- Prabhavathi, K.; Chandra, U.S.J.; Soanker, R.; Rani, P.U. A Randomized, Double Blind, Placebo Controlled, Cross over Study to Evaluate the Analgesic Activity of Boswellia Serrata in Healthy Volunteers Using Mechanical Pain Model. Indian J. Pharmacol. 2014, 46, 475. [Google Scholar] [CrossRef]
- Elston, A.; Lin, J.; Rouseff, R. Determination of the Role of Valencene in Orange Oil as a Direct Contributor to Aroma Quality. Flavour Fragr. J. 2005, 20, 381–386. [Google Scholar] [CrossRef]
- Marques, F.M.; Figueira, M.M.; Schmitt, E.F.P.; Kondratyuk, T.P.; Endringer, D.C.; Scherer, R.; Fronza, M. In Vitro Anti-Inflammatory Activity of Terpenes via Suppression of Superoxide and Nitric Oxide Generation and the NF-ΚB Signalling Pathway. Inflammopharmacology 2019, 27, 281–289. [Google Scholar] [CrossRef]
- Sánchez-Martínez, J.D.; Alvarez-Rivera, G.; Gallego, R.; Fagundes, M.B.; Valdés, A.; Mendiola, J.A.; Ibañez, E.; Cifuentes, A. Neuroprotective Potential of Terpenoid-Rich Extracts from Orange Juice by-Products Obtained by Pressurized Liquid Extraction. Food Chem. X 2022, 13, 100242. [Google Scholar] [CrossRef]
- Jin, J.H.; Lee, D.-U.; Kim, Y.S.; Kim, H.P. Anti-Allergic Activity of Sesquiterpenes from the Rhizomes of Cyperus Rotundus. Arch. Pharm. Res. 2011, 34, 223–228. [Google Scholar] [CrossRef]
- Muthaiyan, A.; Biswas, D.; Crandall, P.G.; Wilkinson, B.J.; Ricke, S.C. Application of Orange Essential Oil as an Antistaphylococcal Agent in a Dressing Model. BMC Complement. Altern. Med. 2012, 12, 125. [Google Scholar] [CrossRef] [Green Version]
- Rao, V.S.N.; Menezes, A.M.S.; Viana, G.S.B. Effect of Myrcene on Nociception in Mice. J. Pharm. Pharmacol. 1990, 42, 877–878. [Google Scholar] [CrossRef] [PubMed]
- Gertsch, J.; Leonti, M.; Raduner, S.; Racz, I.; Chen, J.-Z.; Xie, X.-Q.; Altmann, K.-H.; Karsak, M.; Zimmer, A. Beta-Caryophyllene Is a Dietary Cannabinoid. Proc. Natl. Acad. Sci. USA 2008, 105, 9099–9104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bento, A.F.; Marcon, R.; Dutra, R.C.; Claudino, R.F.; Cola, M.; Pereira Leite, D.F.; Calixto, J.B. β-Caryophyllene Inhibits Dextran Sulfate Sodium-Induced Colitis in Mice through CB2 Receptor Activation and PPARγ Pathway. Am. J. Pathol. 2011, 178, 1153–1166. [Google Scholar] [CrossRef]
- Jansen, C.; Shimoda, L.M.N.; Kawakami, J.K.; Ang, L.; Bacani, A.J.; Baker, J.D.; Badowski, C.; Speck, M.; Stokes, A.J.; Small-Howard, A.L.; et al. Myrcene and Terpene Regulation of TRPV1. Channels 2019, 13, 344–366. [Google Scholar] [CrossRef] [Green Version]
- Chen, G.-L.; Lei, M.; Zhou, L.-P.; Zeng, B.; Zou, F. Borneol Is a TRPM8 Agonist That Increases Ocular Surface Wetness. PLoS ONE 2016, 11, e0158868. [Google Scholar] [CrossRef]
- Lai, H.; Liu, C.; Hou, L.; Lin, W.; Chen, T.; Hong, A. TRPM8-Regulated Calcium Mobilization Plays a Critical Role in Synergistic Chemosensitization of Borneol on Doxorubicin. Theranostics 2020, 10, 10154–10170. [Google Scholar] [CrossRef]
- Granger, R.E.; Campbell, E.L.; Johnston, G.A.R. (+)- And (-)-Borneol: Efficacious Positive Modulators of GABA Action at Human Recombinant Alpha1beta2gamma2L GABA(A) Receptors. Biochem. Pharmacol. 2005, 69, 1101–1111. [Google Scholar] [CrossRef]
- Jiang, J.; Shen, Y.Y.; Li, J.; Lin, Y.H.; Luo, C.X.; Zhu, D.Y. (+)-Borneol Alleviates Mechanical Hyperalgesia in Models of Chronic Inflammatory and Neuropathic Pain in Mice. Eur. J. Pharmacol. 2015, 757, 53–58. [Google Scholar] [CrossRef]
- Vogt-Eisele, A.K.; Weber, K.; Sherkheli, M.A.; Vielhaber, G.; Panten, J.; Gisselmann, G.; Hatt, H. Monoterpenoid Agonists of TRPV3. Br. J. Pharmacol. 2007, 151, 530–540. [Google Scholar] [CrossRef] [Green Version]
- Takaishi, M.; Uchida, K.; Fujita, F.; Tominaga, M. Inhibitory Effects of Monoterpenes on Human TRPA1 and the Structural Basis of Their Activity. J. Physiol. Sci. 2014, 64, 47–57. [Google Scholar] [CrossRef] [Green Version]
- Sherkheli, M.A.; Schreiner, B.; Haq, R.; Werner, M.; Hatt, H. Borneol Inhibits TRPA1, a Proinflammatory and Noxious Pain-Sensing Cation Channel. Pak. J. Pharm. Sci. 2015, 28, 1357–1363. [Google Scholar]
- Selescu, T.; Ciobanu, A.C.; Dobre, C.; Reid, G.; Babes, A. Camphor Activates and Sensitizes Transient Receptor Potential Melastatin 8 (TRPM8) to Cooling and Icilin. Chem. Senses 2013, 38, 563–575. [Google Scholar] [CrossRef]
- Marsakova, L.; Touska, F.; Krusek, J.; Vlachova, V. Pore Helix Domain Is Critical to Camphor Sensitivity of Transient Receptor Potential Vanilloid 1 Channel. Anesthesiology 2012, 116, 903–917. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, T.H.D.; Itoh, S.G.; Okumura, H.; Tominaga, M. Structural Basis for Promiscuous Action of Monoterpenes on TRP Channels. Commun. Biol. 2021, 4, 293. [Google Scholar] [CrossRef]
- Freitas, M.M.; Cavalcante, P.M.; Duarte-Filho, L.A.M.S.; Macedo, C.A.F.; Brito, M.C.; Menezes, P.M.N.; Ribeiro, T.F.; Costa, S.M.; Carvalho, B.A.G.; Ribeiro, F.P.R.A.; et al. Investigation of the Relaxing Effect of a Camphor Nanoemulsion on Rat Isolated Trachea. Chem. Biol. Interact. 2021, 348, 109656. [Google Scholar] [CrossRef]
- Tong, T.; Ryu, S.E.; Min, Y.; de March, C.A.; Bushdid, C.; Golebiowski, J.; Moon, C.; Park, T. Olfactory Receptor 10J5 Responding to α-Cedrene Regulates Hepatic Steatosis via the CAMP–PKA Pathway. Sci. Rep. 2017, 7, 9471. [Google Scholar] [CrossRef] [PubMed]
- Tong, T.; Kim, M.; Park, T. α-Cedrene, a Newly Identified Ligand of MOR23, Increases Skeletal Muscle Mass and Strength. Mol. Nutr. Food Res. 2018, 62, e1800173. [Google Scholar] [CrossRef] [PubMed]
- Kessler, A.; Sahin-Nadeem, H.; Lummis, S.C.R.; Weigel, I.; Pischetsrieder, M.; Buettner, A.; Villmann, C. GABAA Receptor Modulation by Terpenoids from Sideritis Extracts. Mol. Nutr. Food Res. 2014, 58, 851–862. [Google Scholar] [CrossRef] [Green Version]
- Chuang, H.; Neuhausser, W.M.; Julius, D. The Super-Cooling Agent Icilin Reveals a Mechanism of Coincidence Detection by a Temperature-Sensitive TRP Channel. Neuron 2004, 43, 859–869. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andrade Próspero, D.F.; Reis Filho, A.C.; Piauilino, C.A.; Lopes, E.M.; de Sousa, D.P.; de Castro Almeida, F.R. Effects of Isopulegol in Acute Nociception in Mice: Possible Involvement of Muscarinic Receptors, Opioid System and l-Arginine/NO/CGMP Pathway. Chem. Biol. Interact. 2018, 293, 55–60. [Google Scholar] [CrossRef] [PubMed]
- Ellinghaus, P.; Wolfrum, C.; Assmann, G.; Spener, F.; Seedorf, U. Phytanic Acid Activates the Peroxisome Proliferator-Activated Receptor Alpha (PPARalpha) in Sterol Carrier Protein 2-/ Sterol Carrier Protein x-Deficient Mice. J. Biol. Chem. 1999, 274, 2766–2772. [Google Scholar] [CrossRef] [Green Version]
- Zomer, A.W.; van Der Burg, B.; Jansen, G.A.; Wanders, R.J.; Poll-The, B.T.; van Der Saag, P.T. Pristanic Acid and Phytanic Acid: Naturally Occurring Ligands for the Nuclear Receptor Peroxisome Proliferator-Activated Receptor Alpha. J. Lipid Res. 2000, 41, 1801–1807. [Google Scholar] [CrossRef]
- Kitareewan, S.; Burka, L.T.; Tomer, K.B.; Parker, C.E.; Deterding, L.J.; Stevens, R.D.; Forman, B.M.; Mais, D.E.; Heyman, R.A.; McMorris, T.; et al. Phytol Metabolites Are Circulating Dietary Factors That Activate the Nuclear Receptor RXR. Mol. Biol. Cell 1996, 7, 1153–1166. [Google Scholar] [CrossRef] [Green Version]
- Thakor, P.; Subramanian, R.B.; Thakkar, S.S.; Ray, A.; Thakkar, V.R. Phytol Induces ROS Mediated Apoptosis by Induction of Caspase 9 and 3 through Activation of TRAIL, FAS and TNF Receptors and Inhibits Tumor Progression Factor Glucose 6 Phosphate Dehydrogenase in Lung Carcinoma Cell Line (A549). Biomed. Pharmacother. 2017, 92, 491–500. [Google Scholar] [CrossRef]
- Kruska, N.; Reiser, G. Phytanic Acid and Pristanic Acid, Branched-Chain Fatty Acids Associated with Refsum Disease and Other Inherited Peroxisomal Disorders, Mediate Intracellular Ca2+ Signaling through Activation of Free Fatty Acid Receptor GPR40. Neurobiol. Dis. 2011, 43, 465–472. [Google Scholar] [CrossRef]
- Elmazar, M.M.; El-Abhar, H.S.; Schaalan, M.F.; Farag, N.A. Phytol/Phytanic Acid and Insulin Resistance: Potential Role of Phytanic Acid Proven by Docking Simulation and Modulation of Biochemical Alterations. PLoS ONE 2013, 8, e45638. [Google Scholar] [CrossRef]
- Sadgrove, N.J.; Oblong, J.E.; Simmonds, M.S.J. Inspired by Vitamin A for Anti-ageing: Searching for Plant-derived Functional Retinoid Analogues. Skin Health Dis. 2021, 1, e36. [Google Scholar] [CrossRef]
- Shayiranbieke, A.; Liang, Q.; Wang, T.; Ma, J.; Li, G.; Du, X.; Zhang, G.; Wang, C.; Zhao, X. Development of Immobilized Beta1-Adrenoceptor Chromatography for Rapid Discovery of Ligands Specifically Binding to the Receptor from Herbal Extract. J. Chromatogr. A 2022, 1677, 463298. [Google Scholar] [CrossRef]
- Majikina, A.; Takahashi, K.; Saito, S.; Tominaga, M.; Ohta, T. Involvement of Nociceptive Transient Receptor Potential Channels in Repellent Action of Pulegone. Biochem. Pharmacol. 2018, 151, 89–95. [Google Scholar] [CrossRef]
- Vimal, A.; Pal, D.; Tripathi, T.; Kumar, A. Eucalyptol, Sabinene and Cinnamaldehyde: Potent Inhibitors of Salmonella Target Protein l-Asparaginase. 3 Biotech 2017, 7, 258. [Google Scholar] [CrossRef]
- İstİflİ, E.S.; ŞihoĞlu Tepe, A.; SarikÜrkcÜ, C.; Tepe, B. Interaction of Certain Monoterpenoid Hydrocarbons with the Receptor Binding Domain of 2019 Novel Coronavirus (2019-NCoV), Transmembrane Serine Protease 2 (TMPRSS2), Cathepsin B, and Cathepsin L (CatB/L) and Their Pharmacokinetic Properties. Turk. J. Biol. 2020, 44, 242–264. [Google Scholar] [CrossRef]
- Lakhera, S.; Devlal, K.; Ghosh, A.; Chowdhury, P.; Rana, M. Modelling the DFT Structural and Reactivity Study of Feverfew and Evaluation of Its Potential Antiviral Activity against COVID-19 Using Molecular Docking and MD Simulations. Chem. Zvesti 2022, 76, 2759–2776. [Google Scholar] [CrossRef]
- Shervin Prince, S.; Stanely Mainzen Prince, P.; Berlin Grace, V.M. Valencene Post-Treatment Exhibits Cardioprotection via Inhibiting Cardiac Hypertrophy, Oxidative Stress, Nuclear Factor- ΚB Inflammatory Pathway, and Myocardial Infarct Size in Isoproterenol-Induced Myocardial Infarcted Rats; A Molecular Study. Eur. J. Pharmacol. 2022, 927, 174975. [Google Scholar] [CrossRef]
- Nam, J.H.; Nam, D.-Y.; Lee, D.-U. Valencene from the Rhizomes of Cyperus Rotundus Inhibits Skin Photoaging-Related Ion Channels and UV-Induced Melanogenesis in B16F10 Melanoma Cells. J. Nat. Prod. 2016, 79, 1091–1096. [Google Scholar] [CrossRef]
- Yang, I.J.; Lee, D.-U.; Shin, H.M. Inhibitory Effect of Valencene on the Development of Atopic Dermatitis-Like Skin Lesions in NC/Nga Mice. Evid. -Based Complement. Altern. Med. 2016, 2016, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Bonn-Miller, M.O.; ElSohly, M.A.; Loflin, M.J.E.; Chandra, S.; Vandrey, R. Cannabis and Cannabinoid Drug Development: Evaluating Botanical versus Single Molecule Approaches. Int. Rev. Psychiatry 2018, 30, 277–284. [Google Scholar] [CrossRef]
- Caesar, L.K.; Cech, N.B. Synergy and Antagonism in Natural Product Extracts: When 1 + 1 Does Not Equal 2. Nat. Prod. Rep. 2019, 36, 869–888. [Google Scholar] [CrossRef] [Green Version]
- Russo, E.B.; McPartland, J.M. Cannabis Is More than Simply Δ9-Tetrahydrocannabinol. Psychopharmacology 2003, 165, 431–432. [Google Scholar] [CrossRef]
- Wilkinson, J.D.; Whalley, B.J.; Baker, D.; Pryce, G.; Constanti, A.; Gibbons, S.; Williamson, E.M. Medicinal Cannabis: Is Δ9–Tetrahydrocannabinol Necessary for All Its Effects? J. Pharm. Pharmacol. 2003, 55, 1687–1694. [Google Scholar] [CrossRef] [PubMed]
- McPartland, J.M.; Pruitt, P.L. Side Effects of Pharmaceuticals Not Elicited By Comparable Herbal Medicines: The Case of Tetrahydrocannabinol and Marijuana. Altern. Ther. Health Med. 1999, 5, 57–62. [Google Scholar] [PubMed]
- Johnson, J.R.; Burnell-Nugent, M.; Lossignol, D.; Ganae-Motan, E.D.; Potts, R.; Fallon, M.T. Multicenter, Double-Blind, Randomized, Placebo-Controlled, Parallel-Group Study of the Efficacy, Safety, and Tolerability of THC:CBD Extract and THC Extract in Patients with Intractable Cancer-Related Pain. J. Pain Symptom Manag. 2010, 39, 167–179. [Google Scholar] [CrossRef]
- Gallily, R.; Yekhtin, Z.; Hanuš, L.O. Overcoming the Bell-Shaped Dose-Response of Cannabidiol by Using Cannabis Extract Enriched in Cannabidiol. Pharmacol. Pharm. 2015, 6, 75. [Google Scholar] [CrossRef] [Green Version]
- Berman, P.; Futoran, K.; Lewitus, G.M.; Mukha, D.; Benami, M.; Shlomi, T.; Meiri, D. A New ESI-LC/MS Approach for Comprehensive Metabolic Profiling of Phytocannabinoids in Cannabis. Sci. Rep. 2018, 8, 14280. [Google Scholar] [CrossRef] [Green Version]
- Blasco-Benito, S.; Seijo-Vila, M.; Caro-Villalobos, M.; Tundidor, I.; Andradas, C.; García-Taboada, E.; Wade, J.; Smith, S.; Guzmán, M.; Pérez-Gómez, E.; et al. Appraising the “Entourage Effect”: Antitumor Action of a Pure Cannabinoid versus a Botanical Drug Preparation in Preclinical Models of Breast Cancer. Biochem. Pharmacol. 2018, 157, 285–293. [Google Scholar] [CrossRef]
- Nallathambi, R.; Mazuz, M.; Namdar, D.; Shik, M.; Namintzer, D.; Vinayaka, A.C.; Ion, A.; Faigenboim, A.; Nasser, A.; Laish, I.; et al. Identification of Synergistic Interaction Between Cannabis-Derived Compounds for Cytotoxic Activity in Colorectal Cancer Cell Lines and Colon Polyps That Induces Apoptosis-Related Cell Death and Distinct Gene Expression. Cannabis Cannabinoid Res. 2018, 3, 120–135. [Google Scholar] [CrossRef] [Green Version]
- Reddy, D.S.; Golub, V.M. The Pharmacological Basis of Cannabis Therapy for Epilepsy. J. Pharmacol. Exp. Ther. 2016, 357, 45–55. [Google Scholar] [CrossRef]
- Russo, E.B. Cannabis and Epilepsy: An Ancient Treatment Returns to the Fore. Epilepsy Behav. 2017, 70, 292–297. [Google Scholar] [CrossRef]
- Sulak, D.; Saneto, R.; Goldstein, B. The Current Status of Artisanal Cannabis for the Treatment of Epilepsy in the United States. Epilepsy Behav. 2017, 70, 328–333. [Google Scholar] [CrossRef]
- Pamplona, F.A.; da Silva, L.R.; Coan, A.C. Potential Clinical Benefits of CBD-Rich Cannabis Extracts Over Purified CBD in Treatment-Resistant Epilepsy: Observational Data Meta-Analysis. Front. Neurol. 2018, 9, 759. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.; Li, L.; Su, J.; Li, B.; Chen, T.; Wong, Y.-S. Synergistic Apoptosis-Inducing Effects on A375 Human Melanoma Cells of Natural Borneol and Curcumin. PLoS ONE 2014, 9, e101277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Zhao, Y.; Guo, D.; Liu, W.; Liu, Y. Synergistic Antimicrobial Activity of Berberine Hydrochloride, Baicalein and Borneol against Candida Albicans. Chin. Herb. Med. 2017, 9, 353–357. [Google Scholar] [CrossRef]
- Rocha, P.M.d.M.; Rodilla, J.M.; Díez, D.; Elder, H.; Guala, M.S.; Silva, L.A.; Pombo, E.B. Synergistic Antibacterial Activity of the Essential Oil of Aguaribay (Schinus molle L.). Molecules 2012, 17, 12023–12036. [Google Scholar] [CrossRef] [Green Version]
- Farhanghi, A.; Aliakbarlu, J.; Tajik, H.; Mortazavi, N.; Manafi, L.; Jalilzadeh-Amin, G. Antibacterial Interactions of Pulegone and 1,8-Cineole with Monolaurin Ornisin against Staphylococcus Aureus. Food Sci. Nutr. 2022, 10, 2659–2666. [Google Scholar] [CrossRef]
- Pavithra, P.S.; Mehta, A.; Verma, R.S. Synergistic Interaction of β-Caryophyllene with Aromadendrene Oxide 2 and Phytol Induces Apoptosis on Skin Epidermoid Cancer Cells. Phytomedicine 2018, 47, 121–134. [Google Scholar] [CrossRef]
- Ferber, S.G.; Namdar, D.; Hen-Shoval, D.; Eger, G.; Koltai, H.; Shoval, G.; Shbiro, L.; Weller, A. The “Entourage Effect”: Terpenes Coupled with Cannabinoids for the Treatment of Mood Disorders and Anxiety Disorders. Curr. Neuropharmacol. 2020, 18, 87–96. [Google Scholar] [CrossRef]
- LaVigne, J.E.; Hecksel, R.; Keresztes, A.; Streicher, J.M. Cannabis Sativa Terpenes Are Cannabimimetic and Selectively Enhance Cannabinoid Activity. Sci. Rep. 2021, 11, 8232. [Google Scholar] [CrossRef]
- Di Giacomo, S.; Mariano, A.; Gullì, M.; Fraschetti, C.; Vitalone, A.; Filippi, A.; Mannina, L.; Scotto d’Abusco, A.; Di Sotto, A. Role of Caryophyllane Sesquiterpenes in the Entourage Effect of Felina 32 Hemp Inflorescence Phytocomplex in Triple Negative MDA-MB-468 Breast Cancer Cells. Molecules 2021, 26, 6688. [Google Scholar] [CrossRef]
- Santiago, M.; Sachdev, S.; Arnold, J.C.; McGregor, I.S.; Connor, M. Absence of Entourage: Terpenoids Commonly Found in Cannabis Sativa Do Not Modulate the Functional Activity of Δ 9 -THC at Human CB 1 and CB 2 Receptors. Cannabis Cannabinoid Res. 2019, 4, 165–176. [Google Scholar] [CrossRef] [Green Version]
- Heblinski, M.; Santiago, M.; Fletcher, C.; Stuart, J.; Connor, M.; McGregor, I.S.; Arnold, J.C. Terpenoids Commonly Found in Cannabis Sativa Do Not Modulate the Actions of Phytocannabinoids or Endocannabinoids on TRPA1 and TRPV1 Channels. Cannabis Cannabinoid Res. 2020, 5, 305–317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Finlay, D.B.; Sircombe, K.J.; Nimick, M.; Jones, C.; Glass, M. Terpenoids From Cannabis Do Not Mediate an Entourage Effect by Acting at Cannabinoid Receptors. Front. Pharmacol. 2020, 11, 359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McDougall, J.J.; McKenna, M.K. Anti-Inflammatory and Analgesic Properties of the Cannabis Terpene Myrcene in Rat Adjuvant Monoarthritis. Int. J. Mol. Sci. 2022, 23, 7891. [Google Scholar] [CrossRef]
- Raup-Konsavage, W.M.; Carkaci-Salli, N.; Greenland, K.; Gearhart, R.; Vrana, K.E. Cannabidiol (CBD) Oil Does Not Display an Entourage Effect in Reducing Cancer Cell Viability in Vitro. Med. Cannabis Cannabinoids 2020, 3, 95–102. [Google Scholar] [CrossRef] [PubMed]
- Cogan, P.S. The ‘Entourage Effect’ or ‘Hodge-Podge Hashish’: The Questionable Rebranding, Marketing, and Expectations of Cannabis Polypharmacy. Expert Rev. Clin. Pharmacol. 2020, 13, 835–845. [Google Scholar] [CrossRef] [PubMed]
- Brooks, B.D.; Brooks, A.E. Therapeutic Strategies to Combat Antibiotic Resistance. Adv. Drug Deliv. Rev. 2014, 78, 14–27. [Google Scholar] [CrossRef] [PubMed]
- Hu, C.-M.J.; Zhang, L. Nanoparticle-Based Combination Therapy toward Overcoming Drug Resistance in Cancer. Biochem. Pharmacol. 2012, 83, 1104–1111. [Google Scholar] [CrossRef]
- Ma, X.H.; Zheng, C.J.; Han, L.Y.; Xie, B.; Jia, J.; Cao, Z.W.; Li, Y.X.; Chen, Y.Z. Synergistic Therapeutic Actions of Herbal Ingredients and Their Mechanisms from Molecular Interaction and Network Perspectives. Drug Discov. Today 2009, 14, 579–588. [Google Scholar] [CrossRef]
- Spinella, M. The Importance of Pharmacological Synergy in Psychoactive Herbal Medicines. Altern. Med. Rev. J. Clin. Ther. 2002, 7, 9. [Google Scholar]
- Wang, S.; Zhu, F.; Marcone, M.F. Staghorn Sumac Reduces 5-Fluorouracil-Induced Toxicity in Normal Cells. J. Med. Food 2015, 18, 938–940. [Google Scholar] [CrossRef]
- McCune, L.M.; Johns, T. Antioxidant Activity in Medicinal Plants Associated with the Symptoms of Diabetes Mellitus Used by the Indigenous Peoples of the North American Boreal Forest. J. Ethnopharmacol. 2002, 82, 197–205. [Google Scholar] [CrossRef] [PubMed]
- Leyte-Lugo, M.; Britton, E.R.; Foil, D.H.; Brown, A.R.; Todd, D.A.; Rivera-Chávez, J.; Oberlies, N.H.; Cech, N.B. Secondary Metabolites from the Leaves of the Medicinal Plant Goldenseal (Hydrastis Canadensis). Phytochem. Lett. 2017, 20, 54–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Britton, E.R.; Kellogg, J.J.; Kvalheim, O.M.; Cech, N.B. Biochemometrics to Identify Synergists and Additives from Botanical Medicines: A Case Study with Hydrastis Canadensis (Goldenseal). J. Nat. Prod. 2018, 81, 484–493. [Google Scholar] [CrossRef] [Green Version]
- Sepulveda, D.E.; Morris, D.P.; Raup-Konsavage, W.M.; Sun, D.; Vrana, K.E.; Graziane, N.M. Cannabigerol (CBG) Attenuates Mechanical Hypersensitivity Elicited by Chemotherapy-Induced Peripheral Neuropathy. Eur. J. Pain 2022, 26, 1950–1966. [Google Scholar] [CrossRef]
- Sepulveda, D.E.; Vrana, K.E.; Graziane, N.M.; Raup-Konsavage, W.M. Combinations of Cannabidiol and Δ9-Tetrahydrocannabinol in Reducing Chemotherapeutic Induced Neuropathic Pain. Biomedicines 2022, 10, 2548. [Google Scholar] [CrossRef]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chacon, F.T.; Raup-Konsavage, W.M.; Vrana, K.E.; Kellogg, J.J. Secondary Terpenes in Cannabis sativa L.: Synthesis and Synergy. Biomedicines 2022, 10, 3142. https://doi.org/10.3390/biomedicines10123142
Chacon FT, Raup-Konsavage WM, Vrana KE, Kellogg JJ. Secondary Terpenes in Cannabis sativa L.: Synthesis and Synergy. Biomedicines. 2022; 10(12):3142. https://doi.org/10.3390/biomedicines10123142
Chicago/Turabian StyleChacon, Francisco T., Wesley M. Raup-Konsavage, Kent E. Vrana, and Joshua J. Kellogg. 2022. "Secondary Terpenes in Cannabis sativa L.: Synthesis and Synergy" Biomedicines 10, no. 12: 3142. https://doi.org/10.3390/biomedicines10123142