1. Introduction
The use of artificial lighting in horticulture is common in greenhouses when daylight radiation is not sufficient for plant growth: for this scope, supplementary lighting is largely used during winter in Nordic regions and, to a lesser extent, also in southern countries. Plant growth demands the provision of lighting for many hours per day, so artificial light sources used have a relevant impact on energy consumption. The adoption of efficient lamps can reduce the energy demand of greenhouses and also of the production costs.
The selection of the light source is also very important for the quality and the quantity of the vegetable produced, since the light features affect the plant development: the correct growth of plants is not only influenced by the quantity of light reaching the leaf, usually calculated as the photosynthetic photon flux density (PPFD) in μmol/sm
2, but also by the spectral composition of the light source [
1,
2]. The photosynthetic process, which allows the plant to transform radiant energy into chemical energy for producing its nutriment, is operated by different types of photoreceptors, sensitive to light at specific wavelengths. Among the most relevant photoreceptors involved in the photosynthetic process of plants, there are chlorophyll a and b, which absorb light mainly in the blue and in the red region, and partially in the UV region between 300 and 400 nm, and carotenoids, which absorb in the band between 400 and 500 nm.
From the photomorphogenesis point of view, plants development is also affected by the quality of light through the activation of other three types of photoreceptors, the phytochromes, the cryptochromes and the phototropins, which regulate the physiological and morphological responses [
3]. The phytochrome Pr and Pfr are photopigment-sensitive to long wavelength light, with peaks of absorption at 660 nm and 730 nm, respectively, while the cryptochromes cry1 and cry2 absorb mainly in the blue region (with a peak of absorption at 450 nm) and ultraviolet-A (with a peak of absorption at 360 nm). Phototropins, as well as the cryptochromes, are photoreceptors of blue light; seed germination, leaf expansion, stem elongation, and root growth are the main responses related to the plant growth and development mediated by these three typologies of photoreceptors.
Plants use different portions of the luminous spectrum for performing their vital functions and for this reason the spectral composition of light is important for supporting the correct plant growth. In [
4] it was demonstrated the occurrence of physiological and morphological changes in lettuce plants grown under different multi-spectral-component lights; in [
5], the leaves of cucumber grown under red light showed dysfunctions in the photosynthetic parameters and they concluded that even a small amount of blue light is required for a correct leaf response and functioning.
Sunlight is the optimal light source for correct plant growth, as its spectral emission entirely covers the absorption bands of all the photoreceptors [
6]; for indoor plant growth, artificial lights have to be accurately selected, balancing the spectral composition with other technical features. The luminous efficiency is very important since plant growth requests several hours of light every day and this impacts on the energy consumption, as well as on production costs.
A halogen lamp is the artificial source most similar to sunlight, since it emits radiation in the whole photosynthetic active radiation (PAR) range. Nevertheless, it is not adopted for horticulture cultivation for several disadvantageous features [
7]: its low luminous efficiency, due to the fact that only a small part of the absorbed electrical energy is converted into visible light, causes high energy consumption and, consequently, high production costs; the peak in the infrared region produces elevated thermal emission, which can be dangerous for plant foliage at short distances; furthermore, the short lifetime requires frequent replacement of the lamps.
High-pressure sodium lamps (HPS) are often used in greenhouses for supporting the growth of crops, even if their spectral composition, centered in the yellow region, is not optimal for stimulating the absorption by plant photoreceptors. From a spectral point of view, fluorescent lamps are more appropriate for plant cultivation and, in fact, they are often used in growth rooms [
7].
The large development of the new LED technology opened new possibilities in the field of horticultural lighting, due to the advantages offered by their operational parameters [
8]. The very high luminous efficiency of LEDs is surely the most incentivizing feature for crop producers, which can largely employ this light source benefitting from low energy consumption and energy cost; this new solid state lighting (SSL) has a high conversion factor of energy into light and also produces high-intensity light with low wattage. Indeed, other features of LEDs seem fit for indoor plant cultivation [
9]: the very small dimensions and the various possibilities of assembly allow obtaining lamps characterized by different shapes and sizes that can be easily used even in small spaces, such as growth chambers. The low thermal output of LEDs allows placing the luminaries in proximity of plants canopies so that lamps emitting a lower luminous flux than traditional ones can be used. Moreover, the long lifetime performance reduces the maintenance costs. Additionally, the main feature of LED technology is the high capacity to manage the luminous emissions in terms of intensity, as well as in terms of spectral composition: it is possible to tailor the light emission for specific plants requirements, setting the luminous flux in accordance with the level of Photosynthetic Photon Flux (PPF) needed by the plants and the spectral power distribution (SPD) in correspondence to the absorbance bands of plant photoreceptors. Recently, several studies have been developed for optimizing the regulation of LED lighting systems in greenhouses with the aim of obtaining the optimal plant growth and high energy savings [
10,
11,
12].
In [
13] authors investigated which was the best lamp in terms of spectral composition for the stimulation of plants photoreceptors, comparing a high number of standard and innovative light sources: the calculations indicated that the optimal artificial lights for plant development are the blue LED and the warm white power LED.
A number of experiments demonstrated the attitude of LED lighting in supporting plant growth in indoor environments [
9] and some of these studies even showed that LED technology is more suitable than traditional ones. The experiments performed on basil and strawberry plants demonstrated that exposure to LED lamps, compared to fluorescent lamps, causes an increase of plant biomass, fruit yield, antioxidant compounds, and a reduction of nitrate contents [
14]. The use of red plus blue LEDs was suitable for
Lilium bulblets growth, too [
15]: this lighting condition produced bulblets with larger size, higher fresh and dry weight, higher percentage of dry matter, and highest number of roots than fluorescent lighting. In [
4] it was demonstrated that supplemental LED lighting improved the lettuce growth parameters and showed that the photosynthetic utilization efficiency was higher compared to HPS conditions.
Experiments on plant growth and development performed with different lighting spectral compositions confirmed that each band of light emission has specific effects on plant physiology and morphology [
16]. In [
17], blue light influenced leaf morphogenesis, promoting differentiation of mesophyll cells and the formation of intercellular spaces, also having positive effects on leaf thickness and area. Light in the blue wavelengths promotes chloroplast development and increases net photosynthesis, the number of stomata and the thickness of leaves in cherry tomato plants [
18]; blue light also induces a phototropic curvature of oat coleoptiles in the direction of light [
19]. Even if the net CO
2 assimilation rate is reduced, rose growth under blue light alone presents a three-day slower, but more complete, vegetative and floral development [
20].
Red light, through the phytochrome stimulation, affects leaf biomass quantity, increasing the dry weight/fresh weight ratio [
17]; supplemental far red light increases fresh and dry weight, stem length, and leaf area in lettuce, but it reduces chlorophyll, carotenoids, and the concentration of anthocyanins [
21].
As the green color of foliage indicates, green light is mainly reflected by plants and only a small percentage is absorbed by chlorophylls and carotenoids. Specific studies demonstrated that even green light produces effects in plants: in fact, it can contribute to plant development and growth in combination with red and blue light [
22,
23]. The results of [
24] showed that green light alone also supports
Lactuca sativa growth, producing leaf and petiole elongation and higher root and shoot growth at high irradiance values. In the same study the net photosynthesis with monochromatic green light (510 nm) at a high PPF level resulted in being the highest among all the light treatments, including the control condition in which white fluorescent light was used. In [
17] the green light treatment produced the highest leaf weight ratio, specific leaf mass, and leaf density.
Some of the previous studies concluded that a controlled and tailored light spectral composition can be useful for inducing specific plant responses, and also for enhancing nutritional value in vegetables [
19,
20,
21]. In [
25] authors studied the effect of red plus three short-wavelength lights (near-UV, blue, and cyan) on lettuce and concluded that the optimal growth was obtained with red and blue treatment which caused a higher carbohydrate content; in fact about the 90% of light absorption by plant leaves is in the blue and red bands [
26].
Several studies were carried out with the aim of determining which blue/red light composition is the most suitable for stimulating the correct response in plants [
27]. In [
28] the optimal growth of lettuce was obtained with the spectral composition containing 90% red light and 10% blue light: with respect to different percentages and HPS control conditions, this spectral composition caused a higher weight and dry matter, soluble sugar content, and phenol production. In another work basil plants showed the best performance when the blue component in the lighting is higher than the red one (ratio 0.7); in this condition the leaf yield was at a maximum and the fresh weight decreased proportionally with the decreasing of the blue percentage in the spectrum [
14]. In [
29] authors compared the growth of pea shoots under three types of LEDs with different combinations of red and blue light components and a fluorescent lamp; two configurations of PPFD distribution were also tested. It was found that relatively low PPFD distribution (62–87 μmol/sm
2) is better for the growth of pea shoots; furthermore, all three LED lamps produced higher values of fresh weight and chlorophyll content than fluorescent lamps, but there are no significant differences in the results among the three LEDs. Results of [
5] revealed the insurgence of physiological disorders, such as low CO
2 fixation, low photosynthetic rate, and low dry weight accumulation, in plants exposed to red light alone. These dysfunctions did not occur when plants were exposed to blue light alone and they were eliminated adding even a small percentage (7%) of blue light to red light. The comparison of lettuce growth under LED lighting with different spectral compositions demonstrated that under white LEDs both plant biomass and nutritional values were higher than under red-blue LED conditions [
30]. This study indicates that LEDs having a complete spectrum are more suitable for vegetable cultivation than two monochromatic LEDs, as it produces better responses on growth parameters.
The aim of this paper is to test the performance and the role of correlate color temperature (CCT) of white LEDs on the growth of plants. In fact, as previously stated, LEDs emitting in the entire PAR region resulted in being better for plant development than monochromatic blue and red LEDs, but the optimal balancing between red and blue components has not been yet univocally defined.
In this framework, this study aims to analyze the effect of the spectral composition of light on growth regulation, morphological parameters, and chlorophyll contents in indoor cultivation of spinach plants. Spinaches are common food vegetables that grow in the winter period: for this reason they are often cultivated in greenhouses using artificial lighting as the principal or supplementary light. The selection of the correct light source allows the maximization of the vegetable production, optimizing, at the same time, the electrical energy consumption of greenhouses. The experiments of Poulet et al. [
31] demonstrated that even the absolute energy request was lower in the lag period than in the successive growth period, as the energy spent per unit edible biomass (kWh/g
edible), as well as the energy for the unit of biomass produced (kWh/g
total) in
Lactuca sativa, were higher in the grow phase.
Two white LED lighting conditions, having similar spectral power distribution (SPD), but with different percentages in the blue and red bands, were experimented in this work. The use of white LEDs also allowed taking advantage of a complete spectrum on plant growth. This study can improve the knowledge on the usability of LED lighting in indoor plant cultivation, and can help in the choice of the most suitable artificial light source for plant growth.
3. Results
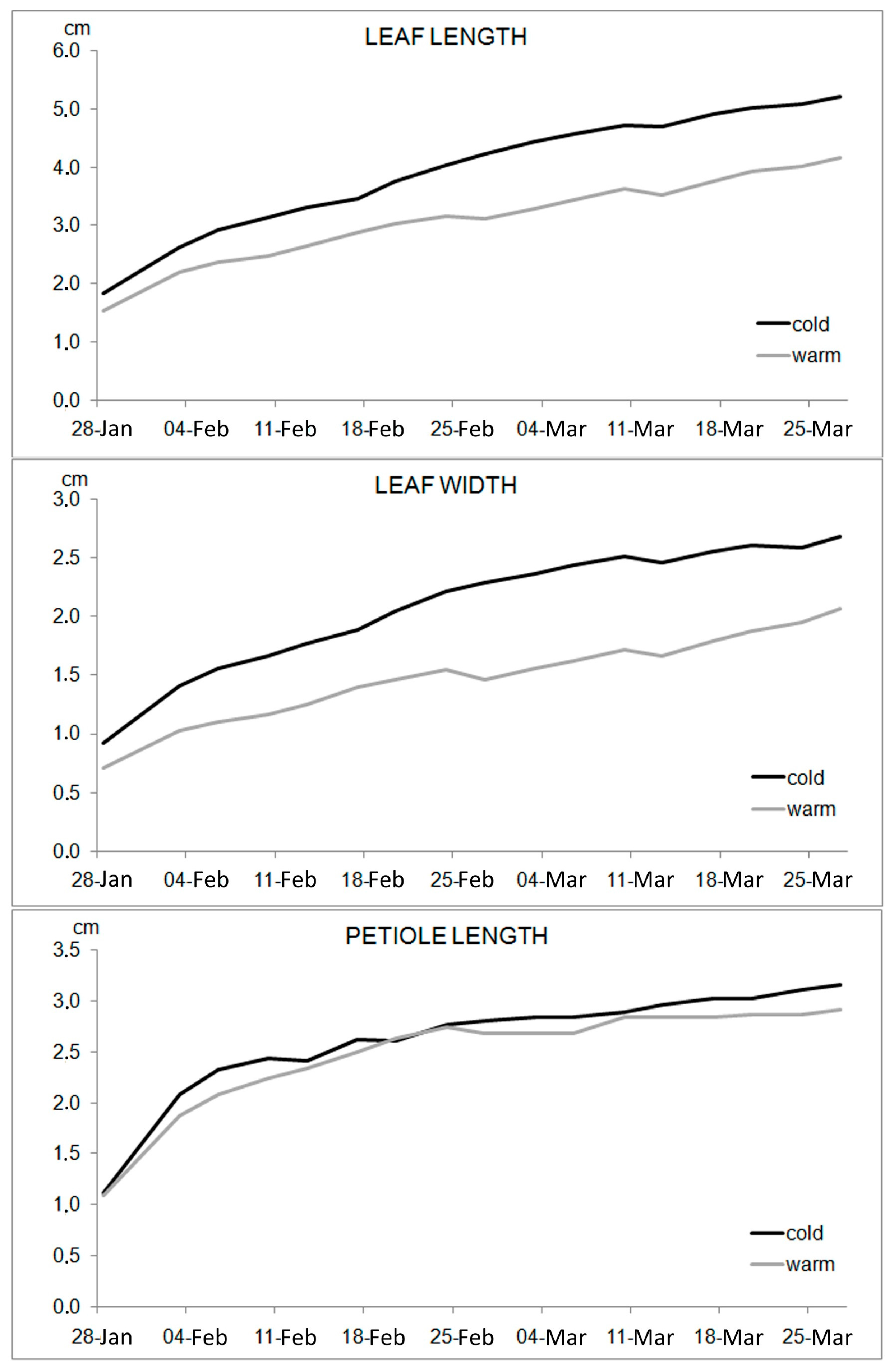
The effect of light spectral composition on plant morphology has been evaluated comparing the increasing dimensions (cm) of leaf parameters during the growth period; the mean growth of 40 leaves (10 leaves per plant) under each of the two lighting conditions is shown in the graphs of
Figure 4. This shows that the spinach plants grow in both of the LED lighting conditions, but under CL the leaves grow longer, larger, and most rapidly: in fact the inclination of the curves representing the CL leaf length and width is higher than that of the curves relative to the WL setting. Instead, no differences were registered in the petiole elongation between the two lighting conditions.
The early plant growth under CL condition seems to also be confirmed by the presence of hastate leaves in these plants since the fourth week, which emerged in the fifth week under WL.
The statistical analysis was developed on the parameters LL, LW, and PL comparing these values in three dates: before the beginning of the experiment (28 January), in the middle of the experimental period after one month of growth (27 February) and at the end of the experiment (25 March).
Before the beginning of the experiment, the LL, LW, and PL of the two samples, consisting of 40 leaves per each of the two lighting condition, did not differ: no statistical difference was found between the CL and WL leaves in terms of LL (Stat t = −0.62; p(T ≤ t) 0.53), LW (Stat t = 1.58; p(T ≤ t) 0.11), and PL (Stat t = 0.98; p(T ≤ t) 0.32).
During and after the experiment, statistical results confirmed the effect of the two different light treatments on LL and LW, but not on PL. Leaf length resulted in significant differences (a = 0.01) as after one month of growth (Stat t = 6.64; p(T ≤ t) << 0.01), as well as at the end of the study (Stat t = 3.51; p(T ≤ t) << 0.01); leaves width resulted significantly different (a = 0.01) as in the middle of the experiment (Stat t = 6.53; p(T ≤ t) << 0.01) as well as at the end of the growth period (Stat t = 5.31; p(T ≤ t) << 0.01); no difference was found on petiole length on 27 February (Stat t = 0.98; p(T ≤ t) 0.32) and on 25 March (Stat t = 2.36; p(T ≤ t) 0.02).
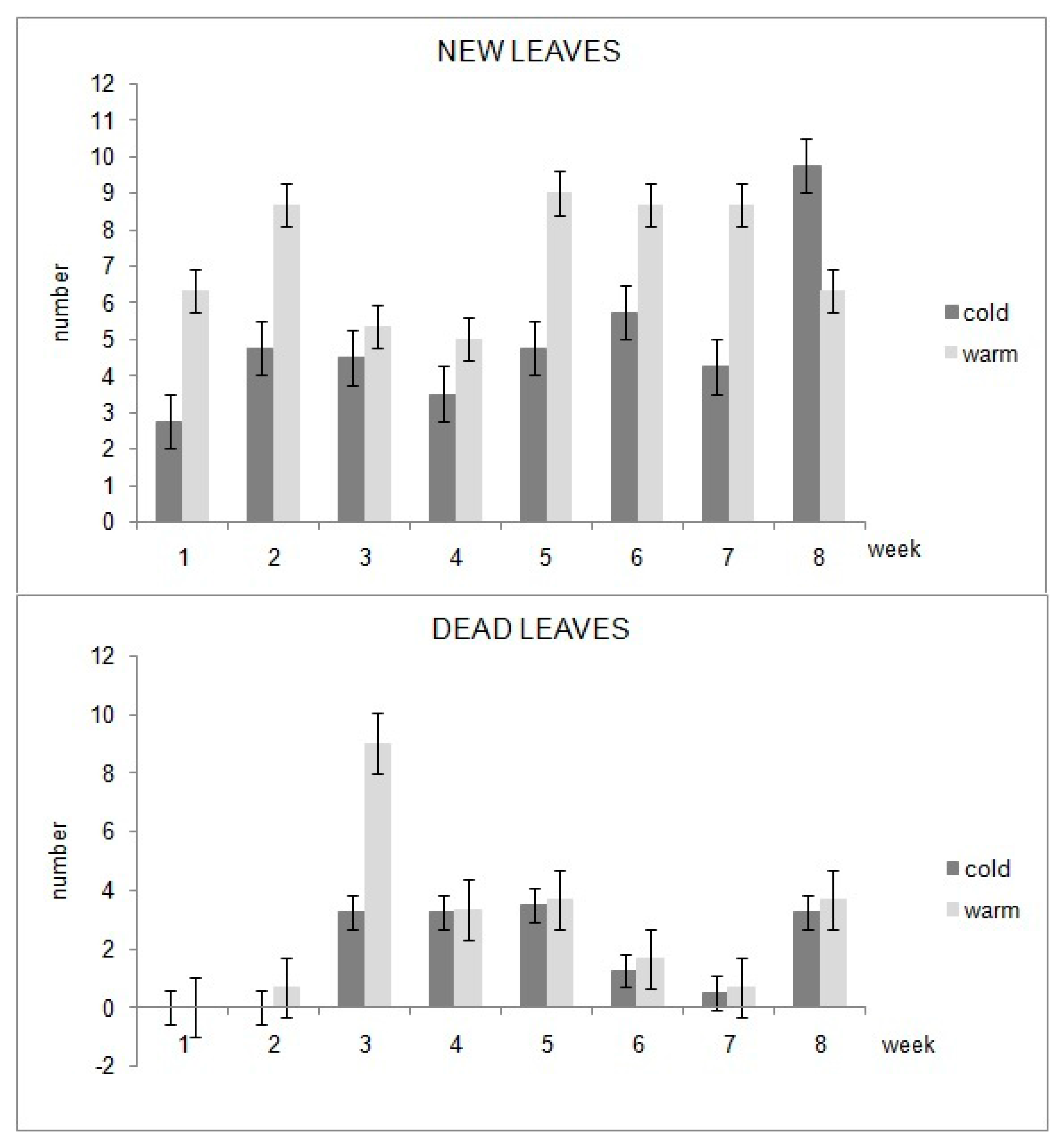
For each week, the number of new and dead leaves per plant has been counted and the mean of the values of the four plants has been calculated under the two light treatments.
Table 2 indicates that the total number of new and death leaves per plant during the entire growth period under WL is higher compared to the CL condition. Additionally, the mean value of new and death leaves per week of the four plants with standard error is shown in
Figure 5; it can be noticed that plants under WL generated a higher weekly number of new leaves in nearly the whole growth period, and also a higher weekly number of dead leaves.
The flowering period was the same for plants in both luminous conditions: in the 11th week spinach plants started producing flowers under CL, as well as WL. Only one stem of one plant flowered with CL, while one stem of two plants flowered under WL. In this last condition another stem flowered in one of the two plants during the 12th week under WL.
At the end of the two months the stem diameter (SD), the root diameter (RD), and the root length (RL) have been measured in each plant and the mean value has been calculated for each luminous condition. Additionally, the chlorophyll content in leaves was measured for each plant at the end of the growth period and the mean values have been calculated for each luminous condition. The results are shown in
Table 3.
The results showed that cold LED lighting fostered the development of stems and roots of spinach plants: plants grown under CL showed higher mean values of SD and RD, while plants subject to WL resulted in having the longest roots.
The statistical analysis showed that the different light treatments produced different effects on SD and RD, but not on RL. Stem diameter (Stat t = 2.67; p(T ≤ t) 0.02) and root diameter (Stat t = 2.39; p(T ≤ t) 0.02) resulted in being significantly larger (a = 0.05) in the plants grown under CL with respect to those grown under WL; while no significant difference resulted in RL (Stat t = 1.56; p(T ≤ t) 0.13) due to the light treatment.
Results show a different concentration of chlorophylls in the basal respect to the apical leaves in plants grown with both WL and CL: in fact, all the plants presented higher contents of chlorophylls in the apical leaves and lower in the basal leaves. The highest concentration of chlorophylls was found in the apical leaves of plants subjected to WL, followed by the apical ones grown under CL, while the lower chlorophylls concentration was found in the basal leaves under CL treatment. In general, the warm LEDs stimulated the chlorophyll content in spinach plants much more compared to the cold LEDs.
The statistical analysis showed that plants exposed to WL produced a significantly higher (a = 0.05) chlorophyll concentration than the plants under CL in the apical (Stat t = −3.97; p(T ≤ t) < 0.02), as well as in the basal (Stat t = −2.42; p(T ≤ t) 0.02), leaves. In this analysis the dry matter weight and the leaves thickness were not measured so the photosynthetic activity cannot be properly estimated. This shortcoming could have partially affected the considerations concerning the experiment.
4. Discussion
In this study several differences in the growth and development of spinach plants depending on the light treatment have been found. The results of two morphological parameters, leaf length (LL) and leaf width (LW), showed that cold LEDs stimulate leaf growth much more than warm LEDs; in fact, plants grown under CL treatments showed longer and larger leaves, compared to the other light treatment. Indeed, no difference was found on petiole length. Moreover, at the end of the growth period plants under CL had a larger stem diameter and root diameter than the plants subjected to WL.
Other studies found similar effects on plant morphology using light treatments of blue wavelengths: supplemental short-wavelength lighting increased leaf area, fresh weight, and dry weight of leaves [
35,
36], and blue LEDs promoted the petiole elongation of leaves [
37], dry weight [
38], fresh weight, and dry matter of the root [
15]. Blue LED light also determines higher leaf area [
38] and an increase of leaf mass per area [
5].
In [
17] it was demonstrated that the increase in leaf area with blue light is due to the development of the palisade parenchyma that facilitates light absorption by chloroplasts, and in [
5] it was found that the increase of blue light percentage determines a higher number of stomata on the leaf. It has been demonstrated that the amount of leaf dimension is connected with the increase in biomass, due to the fact that a larger leaf area allows the capture of a higher number of light rays, enhancing, as a consequence, the photosynthetic process [
30].
Cold light seems to have influenced the growth period, facilitating an advance in development: in this study morphological parameters indicate that plants subjected to CL treatment anticipated their growth of about one week with respect to both WL condition and this hypothesis, supported by the appearance of the hastate leaves in the same plants one week before than in the other plants. Similar findings occurred in the experiment of [
14], where blooming of strawberry plants was anticipated one week earlier when lighting had a predominant blue ratio, and the same happened for the blooming of azalea plants with a higher blue percentage in the lighting [
39].
The results obtained in this study indicate that warm light has a positive effect on the development of new leaves; this could be related to the higher stimulation of phytochrome generated under WL. These photoreceptors are sensitive to long-wavelength light and regulate the morphogenesis in plants. The phenomenon could also be due to the presence of an higher amount of yellow wavelengths in WL compared to the CL (26% vs. 16%), as it was found in a previous study on lettuce which showed an increase of the leaves number due to the treatment with yellow light in the spectral composition [
4].
The root morphology is an index of plant wellbeing, as well-developed roots can enhance the correct plant development through the adequate provision of water and minerals; in our experiment both luminous treatments determined vigorous and long roots (
Table 2). Furthermore, CL seems to enhance root width, as spinach exposed to this lighting showed largest roots.
The experimental results indicated that the highest concentration of chlorophylls in spinach leaf was produced under WL: this effect could be linked to the spectral composition of light emitted by this lamp. In fact, as stated above, it emits a higher percentage of red wavelengths than blue wavelength and, consequently, can correctly stimulate the photosynthetic photoreceptors. The datum is consistent with the literature which has investigated the correct blue/red ratio in the SPD of lighting for obtaining the best plant response in terms of photosynthetic and physiological systems: previous studies demonstrated, indeed, that lettuce and cucumber spectral demand is only 10% of blue and 90% or less of red wavelengths [
5,
21,
28].
The use of two white LEDs, which have a continuous and broad-band spectral power distribution, has allowed the plants to grow, taking advantage of all the wavelengths included in the PAR region; in fact, the green spectral bands which are not absorbed by plant can also have a positive impact on its growth and development. As demonstrated by [
24], seedlings grown under green, red, and blue LED light are longer than those grown under red (630 nm) and blue (470 nm) alone.
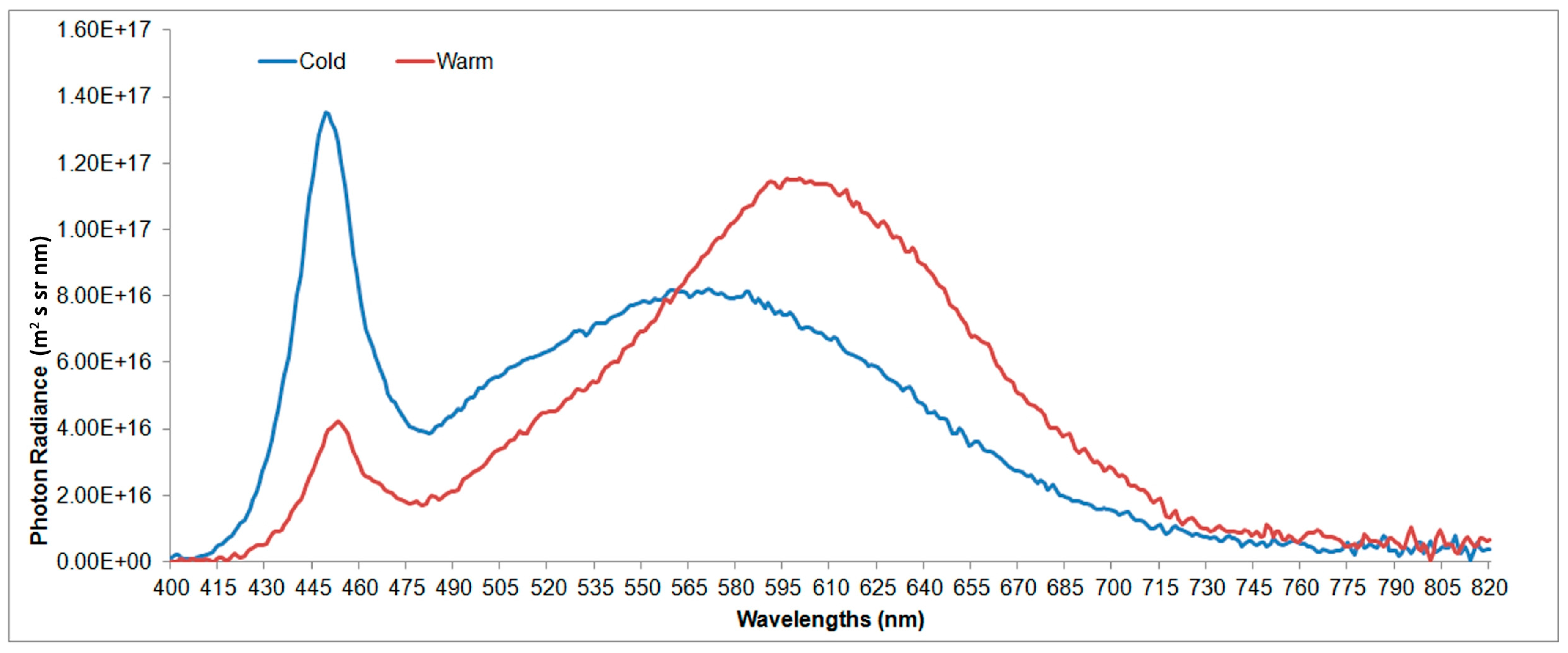
Nevertheless, in this study the percentage of the green component of the two LEDs’ spectra is comparable to that shown in
Figure 2, so it was not possible to evaluate if, and to what extent, this spectral content affected plant growth.
The increase of chlorophylls in plants has a positive impact on the photosynthesis rate, as the CO
2 assimilation in leaves increases [
20], and on the amount of produced chemical elements, in particular, carbohydrates, as they are the main product of the photosynthetic process. Additionally, the existence of a relationship between the chlorophyll concentration and the nitrate status in leaves has been demonstrated [
40]. In several studies the effect of the light spectral composition on the concentration of chemical elements has been investigated; LED lighting containing a small percentage of blue, and a high percentage of red, wavelengths produces elevated sugar and phenol contents in lettuce, but also reduces the presence of nitrate; no effect has been found on antioxidant elements in basil [
14] and on ascorbic acid [
28] and protein in lettuce [
30]. The concentration of chemical elements in edible leaves is very important for the nutritional quality of vegetables.
From an energy point of view, it was observed that white cold LEDs have a slightly lower luminous efficiency compared to the warm LED (103 lm/W and 113 lm/W respectively), but the photon efficiency, as shown in
Table 1, is much higher. This consideration confirms that CL seems to be most suitable for spinach plant growth compared to WL, from both the development and energy efficiency points of view. Results of Poulet et al. [
31] indicated that the energy consumption per biomass produced using white LEDs was similar to that of red+blue LED treatment. Nevertheless, other studies in the literature demonstrated that monochromatic blue and red LEDs are more efficient than white LEDs from the luminous efficiency considered as the ratio between light energy output and electrical energy input [
41].
5. Conclusions
In this work the effect of different spectral compositions of light on the growth and development of spinach plants has been investigated. Spinach has been selected as object of investigation as it is a common food plant, often cultivated in greenhouses as it grows easily in indoor environments. It is also a wintry plant, so its cultivation, occurring under unfavorable natural light conditions, can demand supplementary artificial lighting.
Two white LEDs having different CCT and different percentages of blue and red wavelengths in the spectra, have been employed as experimental treatments. Differently from other studies developed in the literature, where monochromatic LED lamps have been used, in this work the plant development has been investigated under LEDs emitting white light, having a continuous spectrum in the whole PAR region.
The results demonstrated that the spectral composition of light impacts on plant physiology and morphology; in fact, the investigated parameters showed that plants under different luminous conditions had different growth and development.
Warm LED light treatment affected the new leaves’ production and the content of chlorophylls in leaves; the plants subjected to WL resulted in having the highest number of new leaves per week during almost the whole growth period (
Figure 5). Their apical leaves also showed a higher concentration of chlorophylls after two months of treatment compared to the plants under CL (
Table 3). If the chlorophyll amount in leaves is linked to the content of chemical elements like sugars, as other studies indicated, it is possible to hypothesize that plants grown under WL contain elevated nutrients, but this datum has not been investigated in this work.
Cold LED light treatment promoted an increase of plant size, as spinach grown under the CL condition presented the longest and largest leaf area (
Figure 3) and larger stem and root diameter when compared to warm lighting (WL) (
Table 2). Moreover, these plants have shown a more rapid growth compared to the one of plants under WL.
Results of this study indicated that the adoption of cold LEDs would help plant growth more than warm LEDs, in accordance with the improvements in the production of spinach. Moreover, the reduction of the production period would allow the minimization of the energy consumption for a single plant growth and, in general, the production costs. On the other hand, it is also important to highlight that, even if under CL the spinach growth has been higher and more rapid, the WL produced a better development of the photosynthetic photoreceptors, which could be relevant for the production quality.









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}