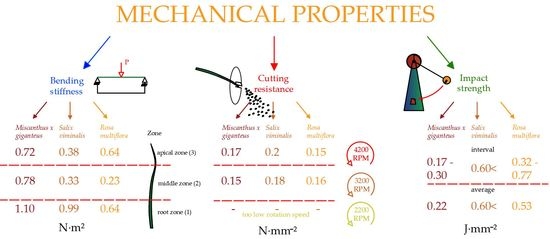
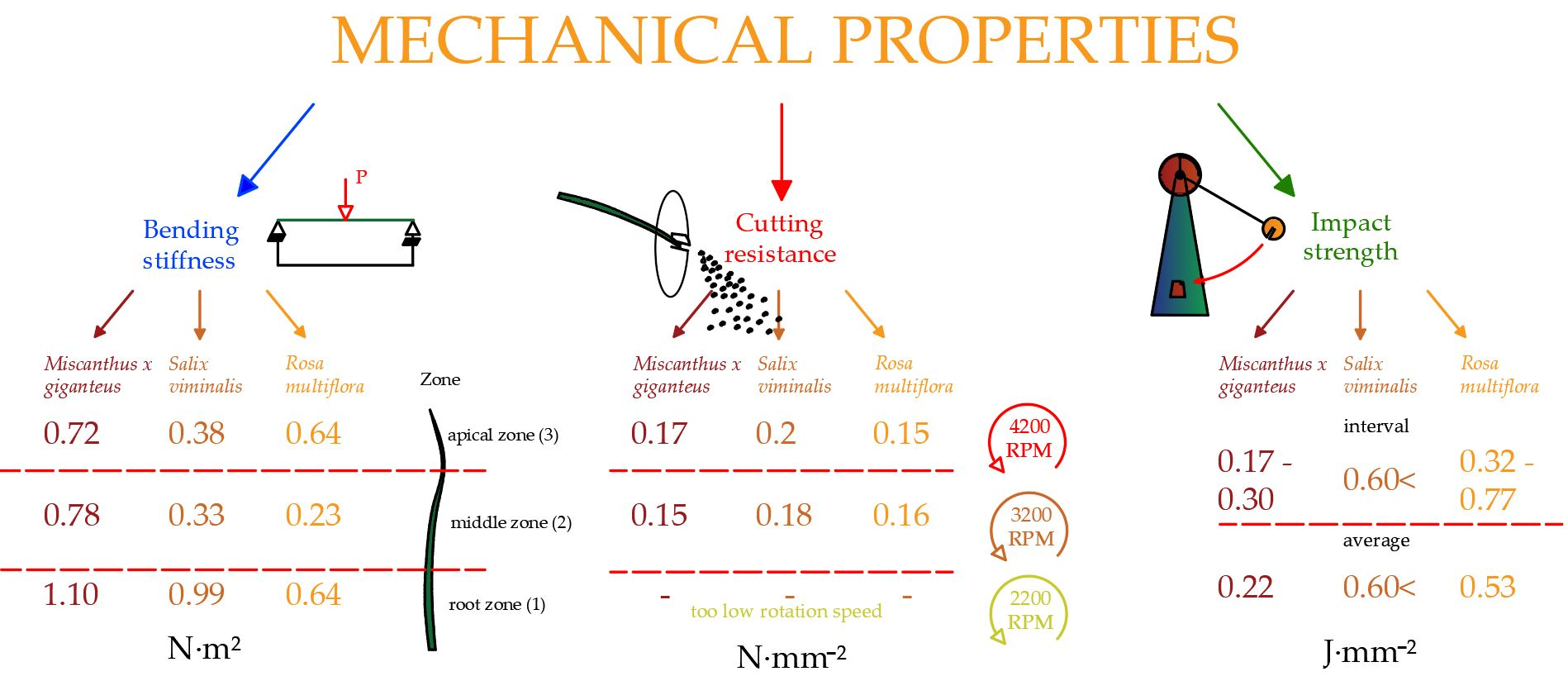
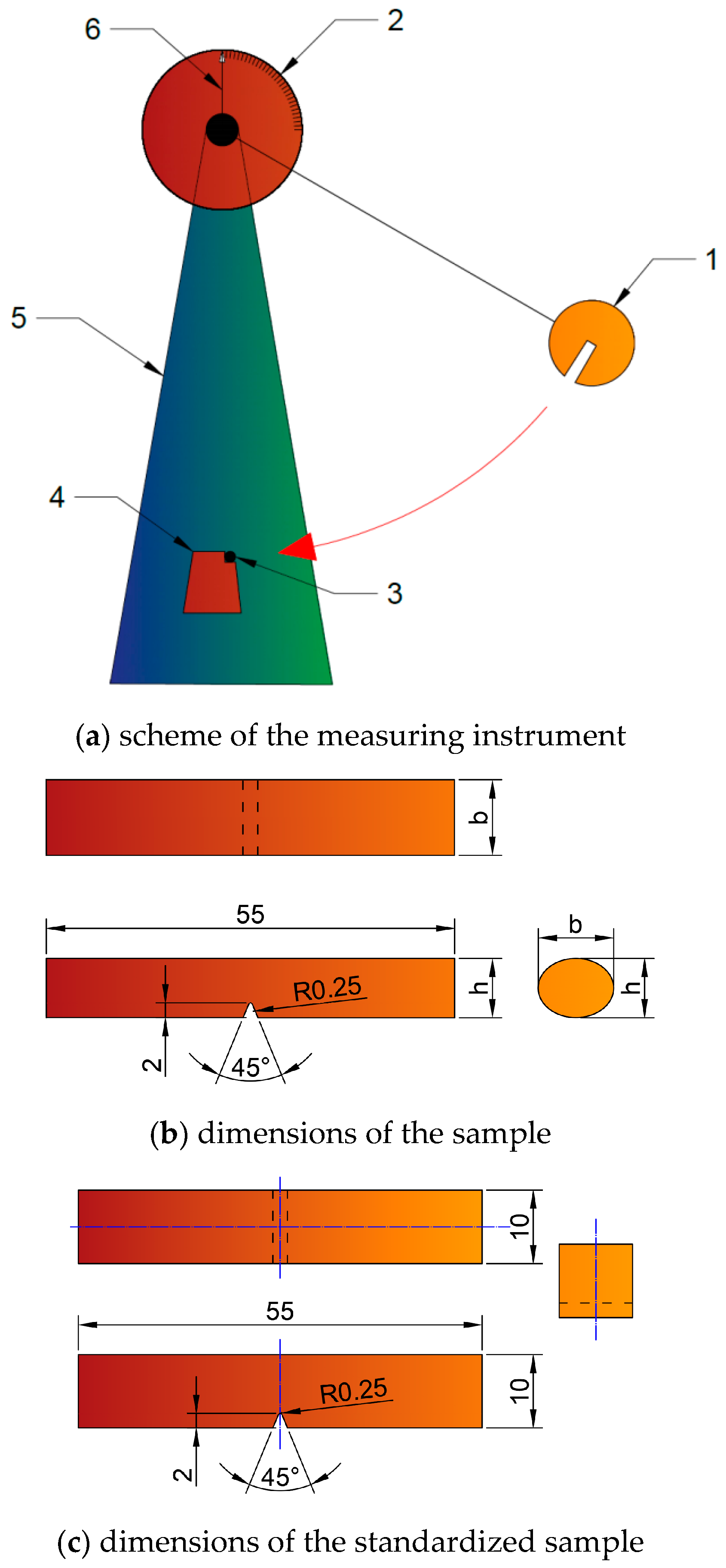
3.2. Cutting Resistance of the Biomass
Table 4 and
Table 5 show the cutting resistances, strength, and power required for the shredding process at initial rotation speeds of 4200, 3200, 2200 RPM. Unfortunately, the tests revealed that 2200 RPM for the 2.5 kW power of the engine was too low to crush the material; therefore, the results are shown for an initial speed of 3200 and 4200 RPM only.
Based on the analysis of
Table 4 and
Table 5, it can be concluded that the highest cutting resistance was achieved by
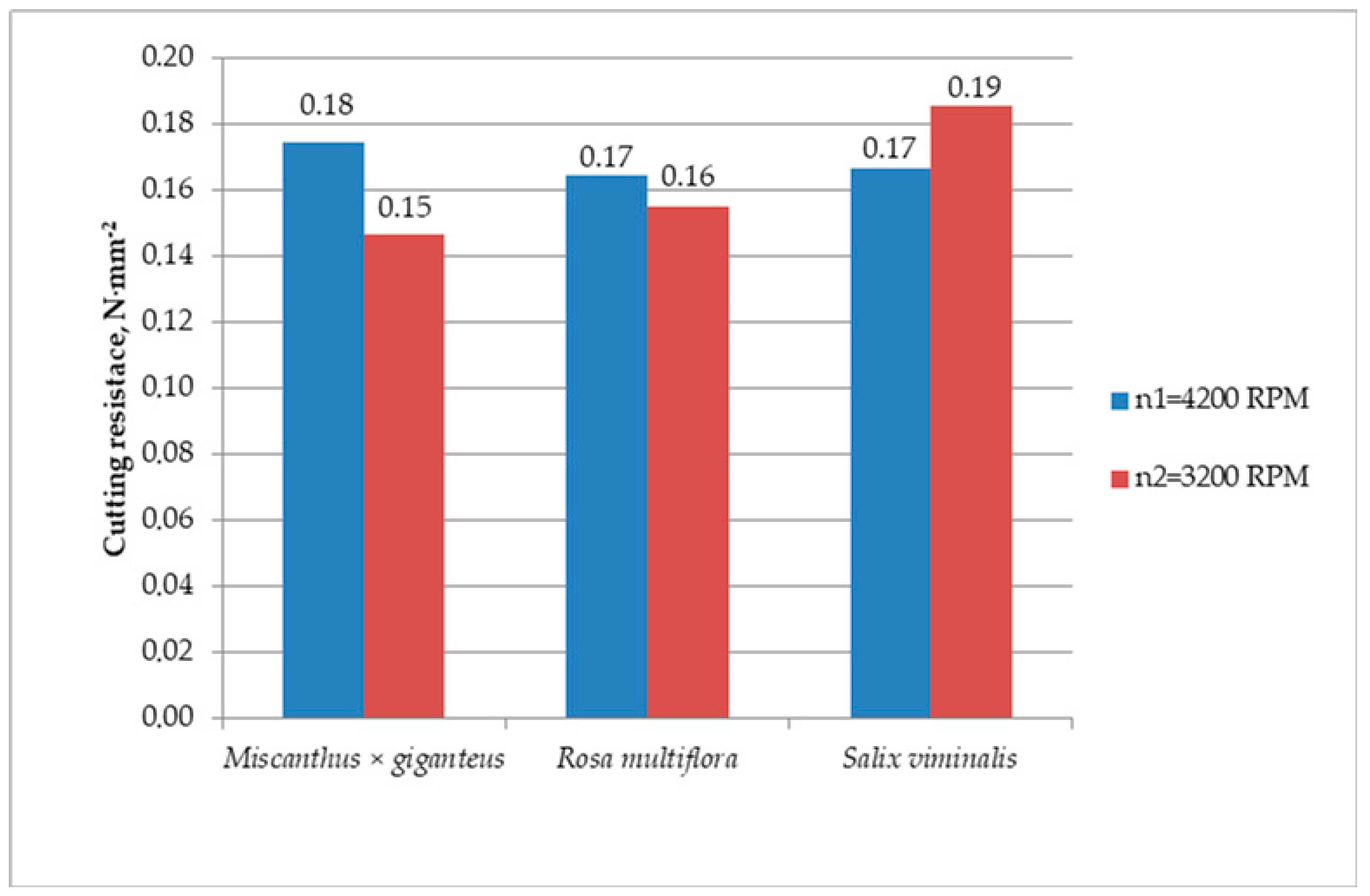
Salix viminalis, reaching values between 0.15 and 0.23 N·mm
−2, for which the cutting resistance increased at the moment of lowering the rotation speed (
Figure 6). In contrast to
Salix viminalis,
Miscanthus × giganteus and
Rosa multiflora showed a reverse strength property, whose cutting resistances were 0.11–0.23 N·mm
−2 and 0.12–0.17 N·mm
−2, respectively. In those cases, the cutting resistance to their shredding increased with the rotation speed. It can be explained by the low modulus of
Salix viminalis with a relatively large diameter of stalks.
Chen et al. [
30] studied the shear force in quasi-static conditions required for Hemp stalks, which amounted to 243 N. However, this value does not give complete information on the strength of the sample since the cross-section of the sample was not taken into account.
Chattopadhyay et al. [
31] examined the Sorghum stalks for the absorption of a part of the potential energy, thus determining the resistance in the form of energy falling on the shear surface. The results were 0.101–0.142 MJ·mm
−2, while Frączek et al. [
29] investigated Topinambur, obtaining results between 0.014 and 0.023 MJ·mm
−2. However, both mentioned measurements concerned the energy consumption of the process (taking into account the cross-section of the sample absorbing the energy), and not the cutting resistance, as examined in this work. Therefore, the comparison of the results was not possible.
Lisowski et al. [
28] checked the resistance and cutting energy of maize stalks in internodes and nodes, obtaining values of 16.9–60.4 N·mm
−2 and 10.93–26.82 J·mm
−2, depending on the measurement zone. These results showed a significant difference between the cutting resistance in the dynamic and static conditions. The cutting in the quasi-static conditions requires much more energy to cut the stalks compared to the dynamic conditions. Lisowski et al. indicated that the cutting resistance of corn stalks was one hundred times higher than the cutting resistance of
Rosa multiflora or
Miscanthus × giganteus at a rotation speed of 3200 RPM obtained in this study (0.15–0.19 N·mm
−2). The cutting energy for rapeseed stalks and pods under dynamic conditions was investigated by Skubisz et al. [
25]. The values of the cutting energy were 9.88·10
4 J·mm
−2 for pods and 11.87·10
4 J·mm
−2 for stalks. However, these values cannot be compared with the results of this study, because they take into account the energy only and not the force acting on the shear surface. Similar research was performed for buckwheat stalks by Woliński et al. [
26], for which the cutting energy amounted to 0.33–0.39 J. Unfortunately, the authors did not include the shear surface of the investigated material, only the energy needed to cut the stem was considered, so the results of the test cannot be compared, as well.
Yore et al. [
32] investigated the cutting energy for rice straw at the cutting speed of 500 strokes per minute. The results showed that as the number of stems (cutting surface) increases, the amount of energy needed to cut one stem decreases. The highest value of shearing energy for one rice straw stem was 0.22 J, while average shearing energy per stem was 0.15 J (for a five-stem test). The highest peak force was 8.3 N for one stem and 5 N per stem for normalized results. Unfortunately, as the energy and the shear force did not correspond to the cutting surface, the values cannot be compared with the results obtained in this study.
In turn, Igathinathane et al. [
33] investigated the influence of the position of the corn stalks relative to the cutting blade. The results showed that the highest cutting force required the corn stalks to be oriented perpendicular to the blade (242 N), while for stalks set at an angle of 45 degrees, the force was 127 N, and for the parallel set only 46 N. In the case of these tests, the cutting cross-section was also not taken into account; moreover, the tests were carried out under quasi-static conditions, which prevented comparison of the results.
Unfortunately, none of the mentioned publications ([
25,
26,
28,
29,
30,
31,
32,
33]) contained studies related to the shear force under the dynamic conditions and referred to the cross-section of the sample.
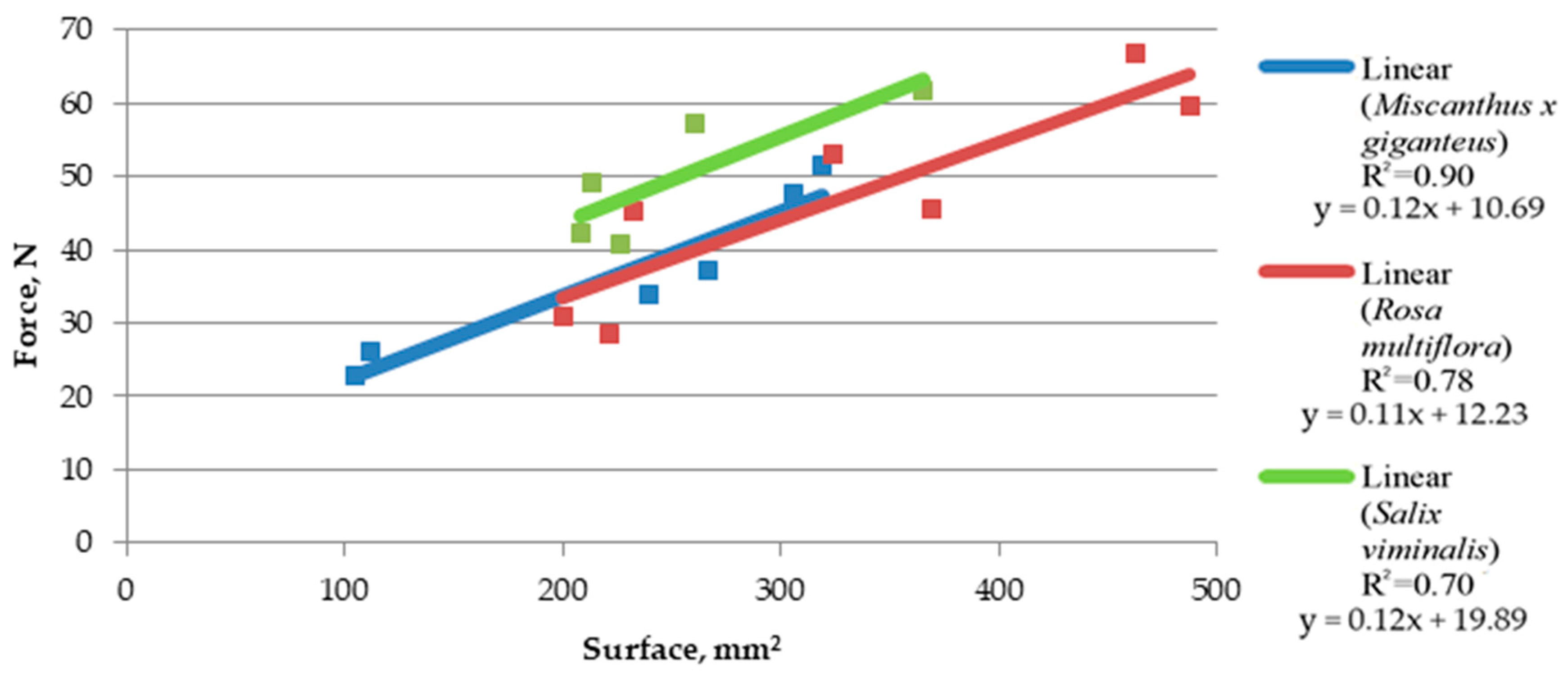
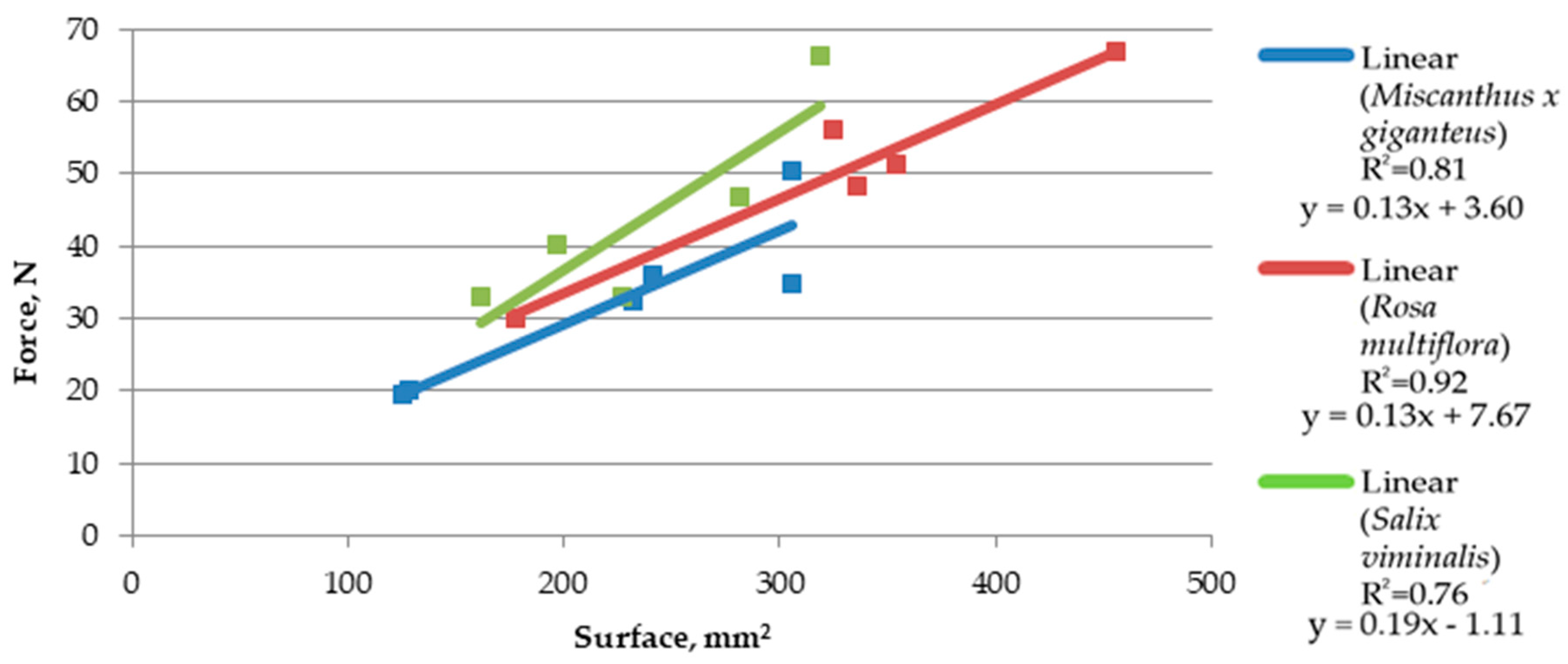
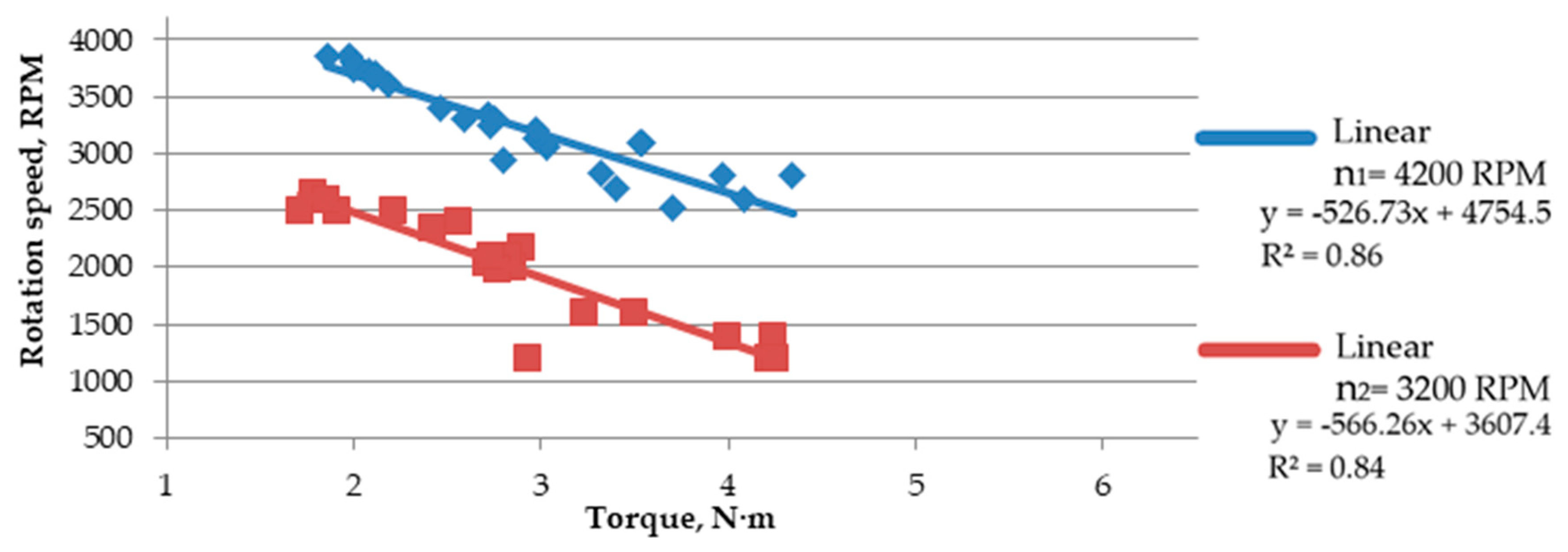
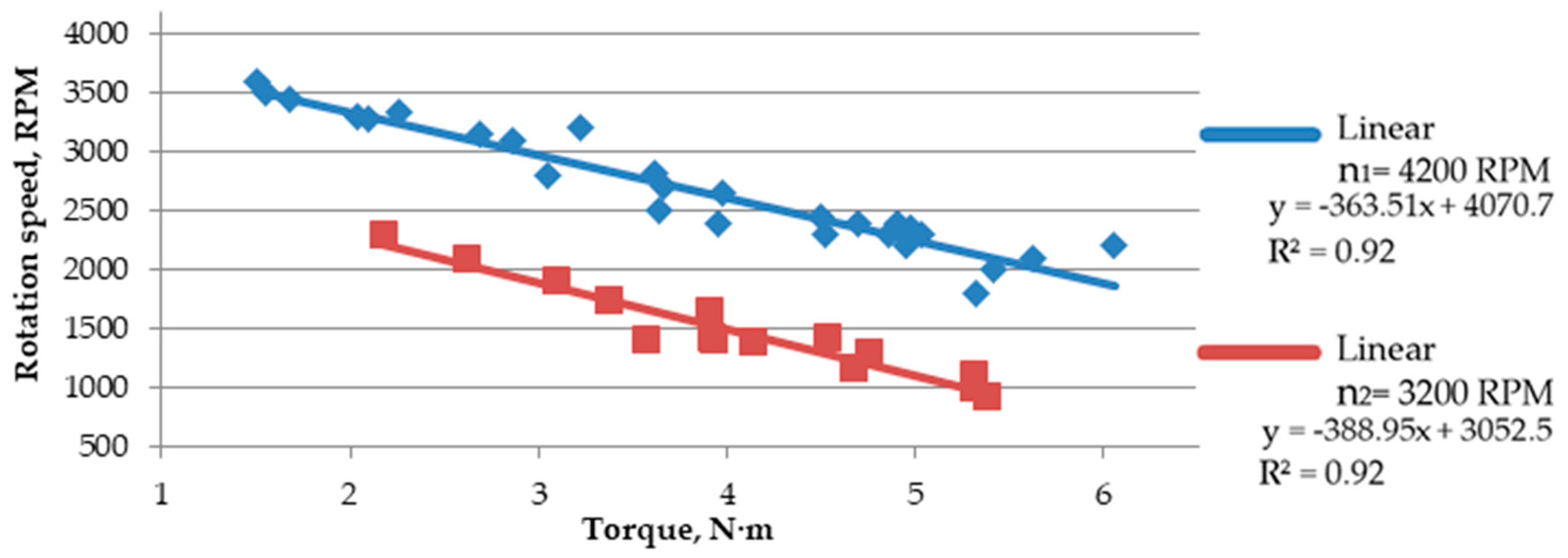
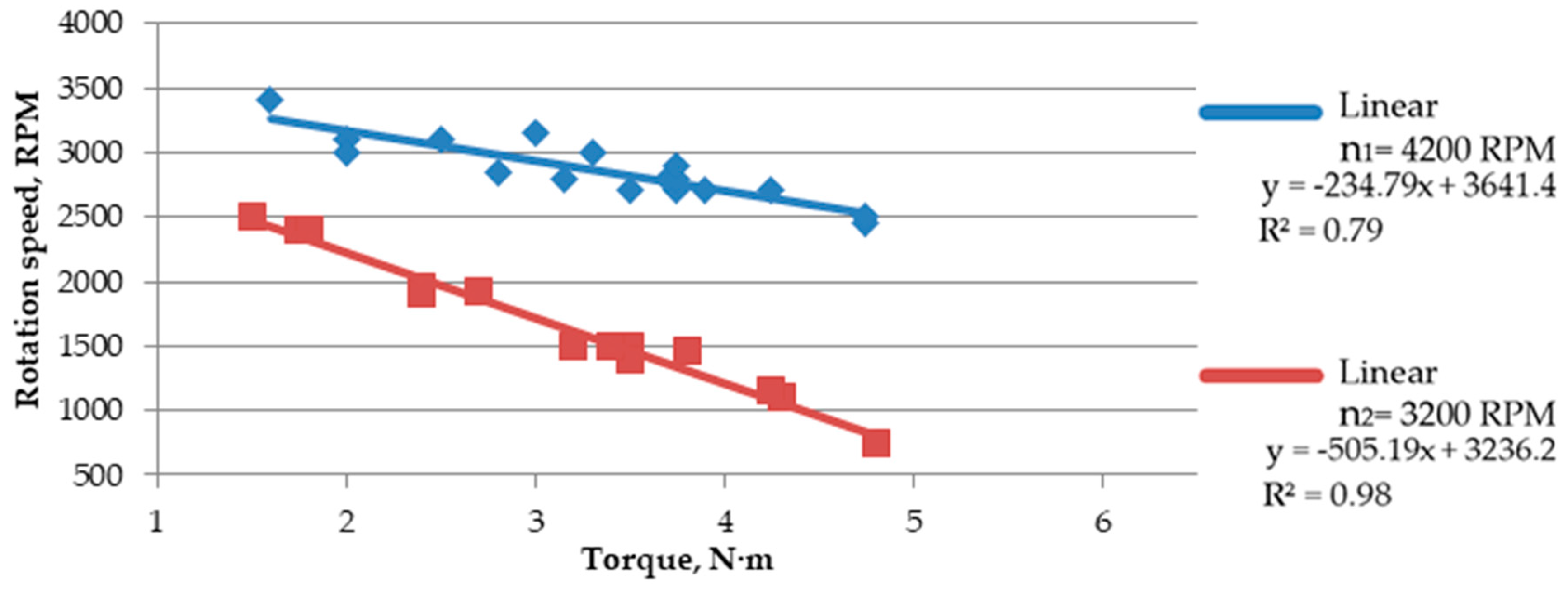
Based on the trend line (linear equation, in this case), it was observed that the increase in rotation speed causes a slight reduction in shearing forces (
Figure 7 and
Figure 8). Such a slight difference means that the force needed for comminution is similar in both cases; however, a minimal increase in force when lowering the speed indicates that a further increase in force can be expected along with lowering the rotation speed, as it was detected using a speed of 2200, at which cutting resistance was not high enough to make it impossible to crumble the material. At a rotation speed of 4200 RPM, the force concerning the shear area increased more linearly. Additionally, the
Miscanthus × giganteus at a rotation speed of 4200 RPM showed a similar relationship of force to the surface like for
Rosa multiflora, while at a speed of 3200 RPM,
Rosa multiflora showed a higher cutting resistance.
The increasing torque causes a reduction of the rotation speed under the load, which increases the shear force. The highest decrease in rotation speed was observed for
Salix viminalis, characterized by the highest value of the shear resistance, while the lowest decrease was observed for
Miscanthus × giganteus, characterized by the lowest shear resistance (
Figure 9,
Figure 10 and
Figure 11).
Additionally, the application of the statistical method ANOVA (Analysis of Variance) revealed that the surface (cross-section area) had the greatest influence on the cutting force for
Miscanthus × giganteus and
Rosa multiflora. In the case of
Salix viminalis, a weak dependence of strength on the other parameters was observed. The details of this analysis (at the probability value
p < 0.05) are presented in the
Supplementary Materials (Table S1). In turn, for
Miscanthus × giganteus, the analysis showed a dependence of cutting resistance on all invetigated parameters (
Table S2 in Supplementary Materials).
Analysis of force variance and cutting resistance depending on the initial speed and decrease in rotation, presented in
Supplementary Materials (Tables S3 and S4) showed the relationship between force and rotation speeds for
Miscanthus × giganteus and
Salix viminalis. However, the relationship between cutting resistance and rotation speeds was observed only for
Miscanthus × giganteus. There was no correlation between the considered parameters for
Salix viminalis.The studies showed also that for a greater rotation speed (4200 RPM in this case) the power demand for the investigated energy plants chipping increased in relation to the lower rotation speed (3200 RPM in this case). The power demand can be calculated using the following formula:
where: P is a power (W), n is a rotation speed (RPM), and M is a torque (N·m).
For a rotation speed of 4200 RPM, the power needed to chop the material was 802 W for Miscanthus × giganteus, 806 W for Rosa multiflora and 896 W for Salix viminalis, while at 3200 RPM the power demand for Miscanthus × giganteus was 438 W for Rosa multiflora 482 W and 467 W for Salix viminalis. The differences between the power demanded for branch chipping at the given rotation speeds (3200 RPM and 4200 RPM) differed by 48% for Salix viminalis, 45% for Miscanthus × giganteus and 40% for Rosa multiflora, respectively. Thus, the rotation speed indirectly influences energy consumption during biomass processing (cutting, chipping, shredding). As a result, the rotation speed significantly affects also the energy efficiency of those processes, and its selection is an indispensable parameter that should not be overlooked during the selection of the equipment and settings of the operation conditions. During the test, it was observed that for Miscanthus × giganteus and Rosa multiflora, the cutting resistance was increasing along with rotation speed, in contrast to Salix viminalis. This relationship is due to Salix viminalis requiring higher blade rotation speeds because it is resistant to impact forces and its comminution is only due to the cutting forces. It is important information from a biomass processing point of view, indicating that the cutting efficiency, the energy consumption and the equipment used at a given rotation speed may not be effective for every type of energy plant.
3.3. Impact Strength Test of the Biomass
Based on the results of the tests carried out, it was found that the highest impact strength value was observed for
Salix viminalis. The energy needed to break it went beyond the range of the measuring instrument and amounted to above 0.57 J·mm
−2.
Table 6 and
Table 7 show the results of the Charpy impact test for
Miscanthus × giganteus and
Rosa multiflora. Based on the results, it can be concluded that
Rosa multiflora has a higher impact strength than
Miscanthus × giganteus. For
Rosa multiflora, the energy absorbed exceeded twice the measuring range of the stand.
The differences in measurements were significant, and resulted from both the surface of the sample cross-section and the sampling zone, for the
Miscanthus × giganteus standard deviation was 0.15 J·mm
−2 and for the
Rosa multiflora 0.06 J·mm
−2. The highest value of
Rosa multiflora impact strength was 0.77 J·mm
−2, with an average impact strength 0.53 J·mm
−2, while the lowest value was 0.32 J·mm
−2. For
Miscanthus × giganteus, the highest value amounted 0.30 J·mm
−2 and the lowest one was 0.13 J·mm
−2. Moreover, applying ANOVA, a high dependence of impact strength (at the
p < 0.05) on the surface was found for
Miscanthus × giganteus, as shown in the
Supplementary Materials (Table S5).
The impact tests carried out led to the conclusion that Miscanthus × giganteus belongs to the group of plants characterized by a low impact strength, which indicates its fragility. Therefore, it is more suitable for impact crushing than cutting. The difference in impact strength in the tested plants proves that the grinding methods should be properly selected, depending on their mechanical properties. Grinding a plant with a low impact strength (Miscanthus × giganteus) using knives may be unnecessary and economically unprofitable, as its apparent cutting may result from its crushing. Then, it is unnecessary to use and constantly replace expensive blades, because a better solution would be to use a hammer mill.
3.4. Bending Stiffness of the Biomass
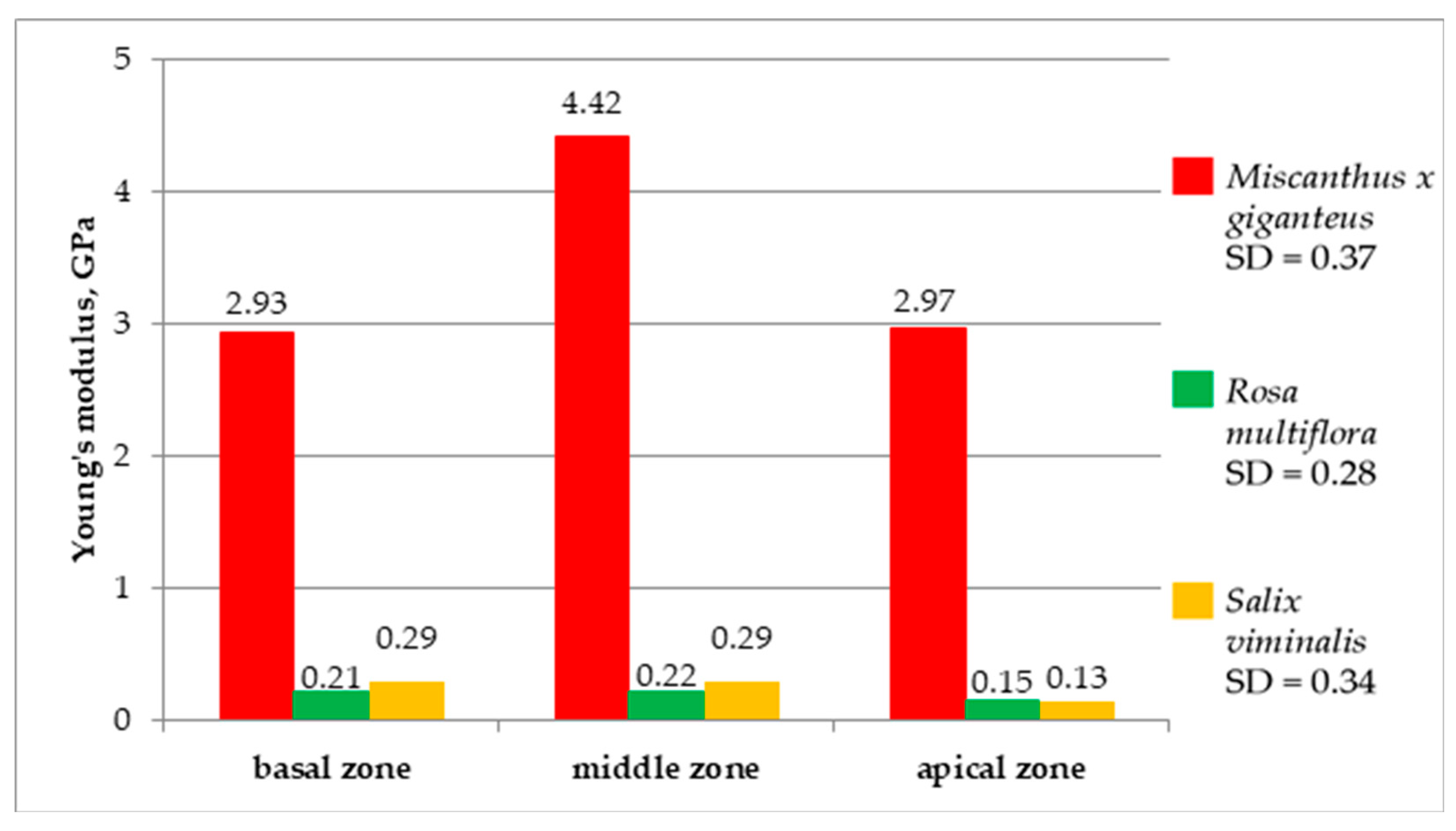
The dependence of Young’s modulus, bending stress and bending stiffness on the stalk measurement zone are shown in
Figure 12,
Figure 13 and
Figure 14. In the case of Young’s modulus (
Figure 12), significantly higher values were observed for
Miscanthus × giganteus. The greatest value was obtained in the middle zone of the stalk (4.42 GPa). In the basal and apical zones, Young’s modulus amounted to 2.93 GPa and 2.97 GPa, respectively. The maximum achieved in the middle zone can be explained by the fact that
Miscanthus × giganteus stalk is thick, but hollow in the basal zone, which weakens it in terms of the strength of the material against bending. The middle zone is filled with tissue in the whole volume and relatively thicker and stronger than the apical zone. In the case of
Salix viminalis and
Rosa multiflora, Young’s modulus was much lower. Depending on the zone, for
Salix viminalis, it was 0.13–0.29 GPa, and for
Rosa multiflora, it was 0.15–0.22 GPa. This arises from the fact, that both plants have morphological features completely different from
Miscanthus × giganteus.
Salix viminalis and
Rosa multiflora are flexible and contain much more reinforcing tissue cells responsible for their elasticity.
These results are in line with the values obtained by Nowakowski et al. [
35]. The Young’s modulus determined by the authors were as follows: for
Salix viminalis, 0.29–0.58 GPa;
Rosa multiflora, 0.85–1.67 GPa,
Miscanthus × giganteus, 1.70–1.84 GPa. The deviations in the measurement results are probably caused by the differences in the maturity of the plants, the time of harvesting, and the moisture content of the material. Kaack et al. [
48] also performed the tests of Young’s modulus for
Miscanthus × giganteus, reaching similar values in the range of 2–8 GPa.
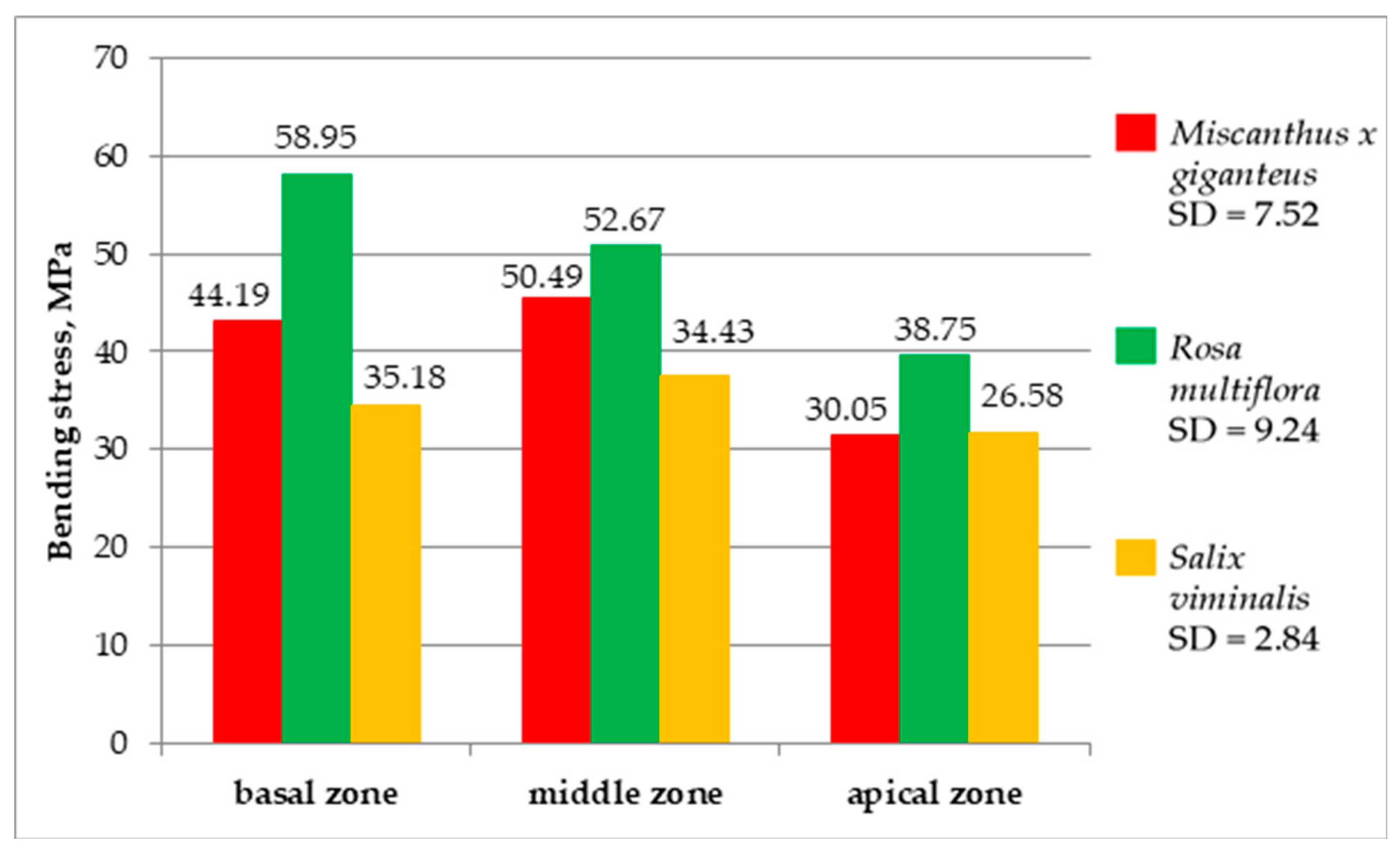
The results of bending stress for the investigated energy plants are shown in
Figure 13. The highest values of bending stresses reached
Rosa multiflora 38.75–58.95 MPa.
Miscanthus × giganteus oscillated within 30.05–50.49 MPa, while the values for
Salix viminalis were in the range 26.58–35.18 MPa.
Differences were found in bending stresses between the zones. Rosa multiflora and Salix viminalis achieved the highest values of bending stress in the basal zone and the lowest in the apical zone. Meanwhile, Miscanthus × giganteus achieved the highest value of bending stresses in the central zone. The reason for this was that the central zone was characterized by the highest strength properties, because it was lignified enough to have a high concentration of reinforcing tissue, while it was not ripe enough to develop a hole in the stalk, weakening the stalk to forces acting along its cross-section.
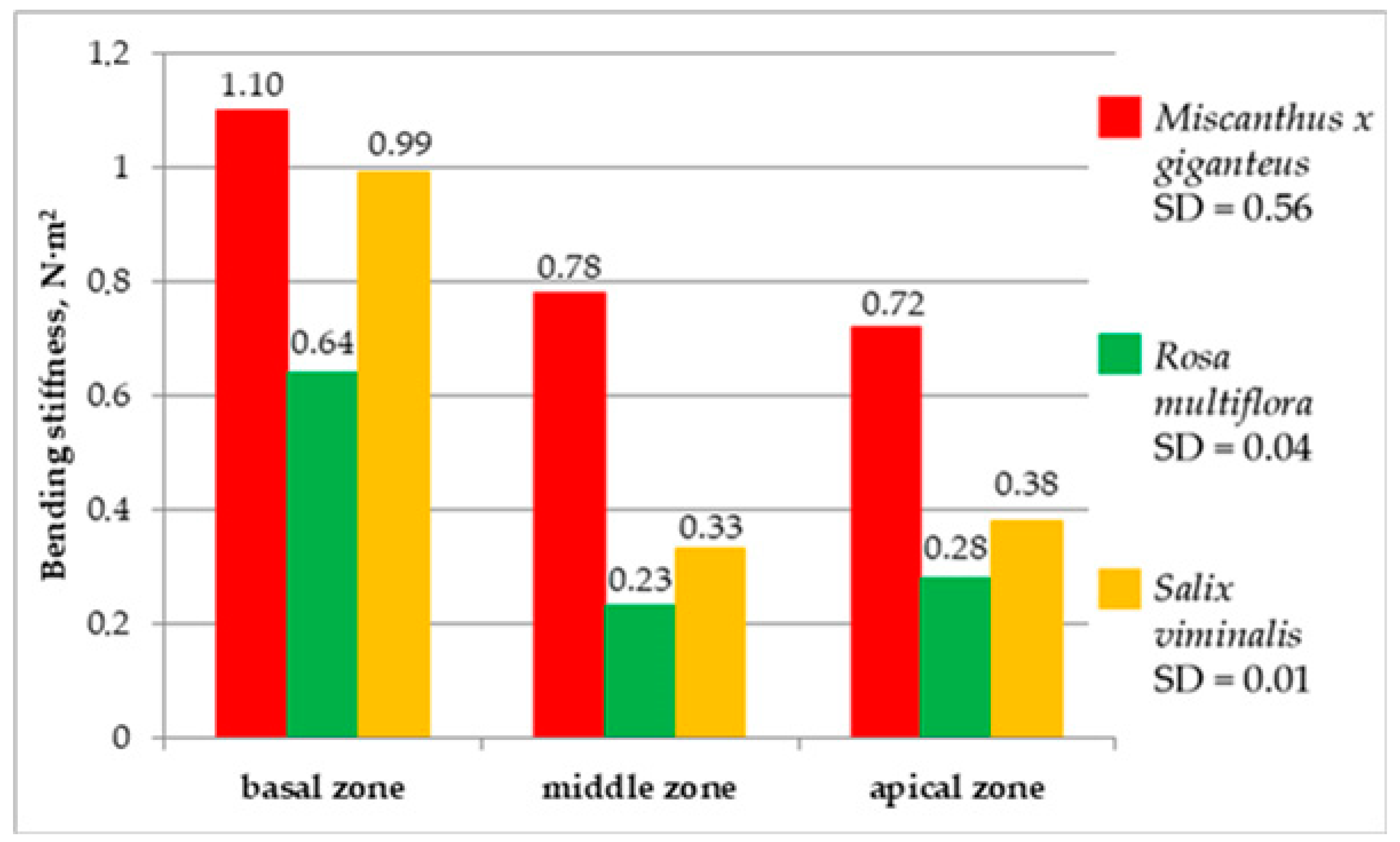
Finally, the bending stiffness in three zones of the energy plants was investigated (
Figure 14). The highest bending stiffness was achieved by
Miscanthus × giganteus. The values amounted as follow: 1.1 N·m
2 for the basal zone, 0.78 N·m
2 in the central zone and 0.72 N·m
2 for the apical zone.
The lowest value of bending stiffness was recorded for Rosa multiflora, for which the highest result was achieved by the basal zone (0.64 N·m2) and the lowest by middle zone 0.23 N·m2, in the apical zone the value of the stiffness was 0.28 N·m2. Salix viminalis achieved the following results: for the basal zone, 0.99 N·m2; middle zone, 0.33 N·m2; and apical zone 0.38 N·m2. The basal zone was characterized by the greatest stiffness for all investigated energy plants. As the plant grows, the thickness of the stalk increases, reducing its flexibility, as well as its susceptibility during the baling process. On the other hand, too low a stiffness of the plant may have a negative impact on the cutting process; however, tests of cutting resistance have determined that at given stiffness the middle and apical zone of plants are susceptible to crushing by the knife method.
During the cutting resistance tests, the Miscanthus × giganteus was shredded due to the impact strength of the blade than because of the cutting force. Such a behavior of the investigated biomass is confirmed by the results of the bending stiffness, stress and Young’s modulus tests. Miscanthus × giganteus was the most fragile plant characterized by the highest bending stiffness and much higher Young’s modulus in relation to Rosa multiflora and Salix viminalis.
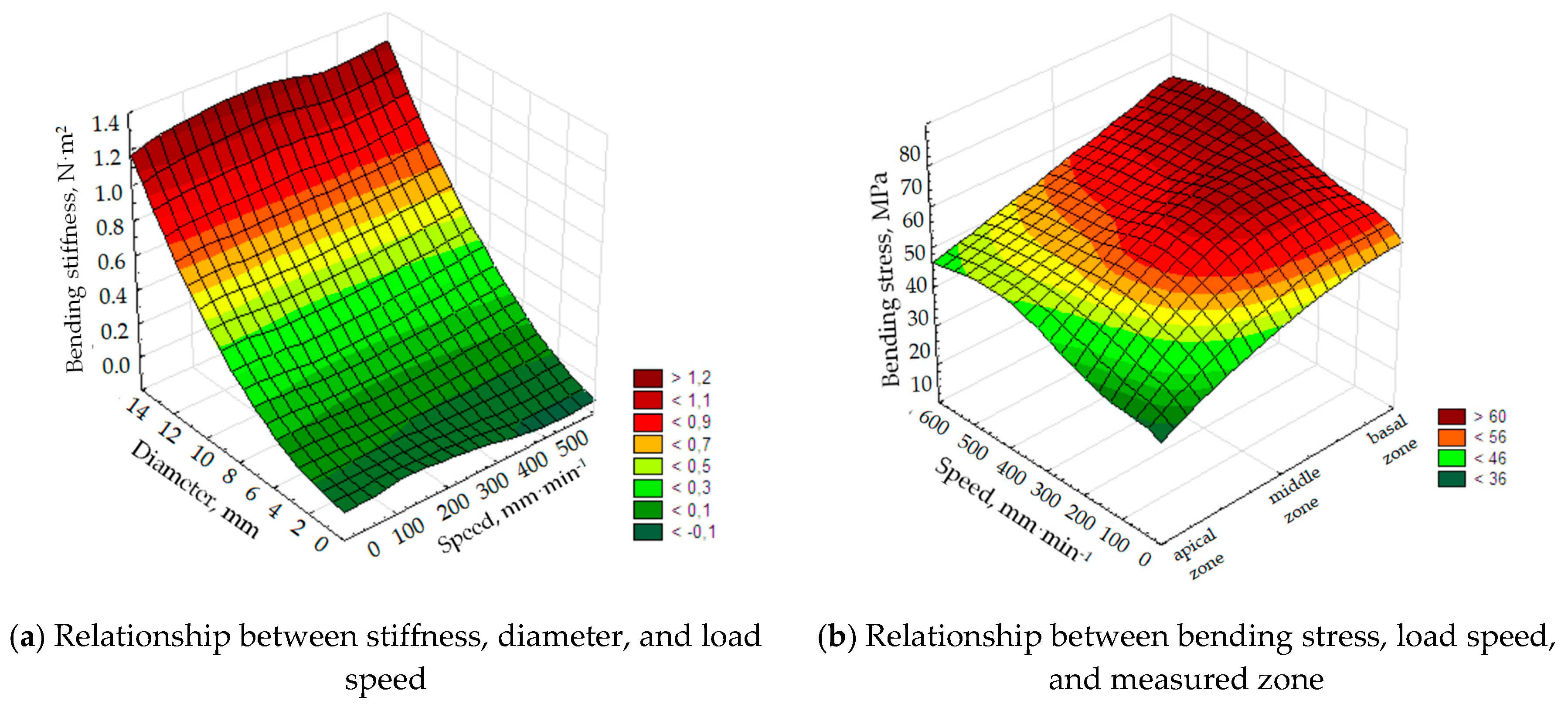
Additionally, the initial correlation between physical parameters was studied. Based on graphical maps in
Figure 15, it can be concluded that the bending stiffness increased with the sample diameter. The speed of sample loading did not significantly affect the results of bending stiffness and bending stress. The apical zone was characterized by smaller stresses during sample loading, which was related to the bending rigidity decreasing along with the diameter.
The analysis of variance was also performed in the case of the tests of the bending stiffness (
Tables S6 and S7 in the Supplementary Materials). The aim was to determine the influence of the stalks’ diameter, measuring zone and speed of sample loading on the bending stiffness of the investigated energy plants (stalks). According to the results shown in
Table S7, a significant dependence of bending stiffness on the diameter was observed for
Rosa multiflora and
Salix viminalis. For
Miscanthus × giganteus, this correlation was not as relevant. The bending stiffness is also affected by speed and measuring zone (
Table S7), but these parameters do not have an influence as great as that of the diameter of the plant (
Table S6). The results seem to be satisfactory, but they need further verification, including tests with the use of other plants.




















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}