
Operation Regimes: A Comparison Based on Nannochloropsis oceanica Biomass and Lipid Productivity


 , ,
, ,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Microalgae Strain and Culture Media
2.2. Culture Scale-Up
2.3. Operation Regime Trial
2.4. Growth Assessment
2.5. Elemental Analysis
2.6. Fatty Acid Profile
2.7. Statistical Analyses
3. Results and Discussion
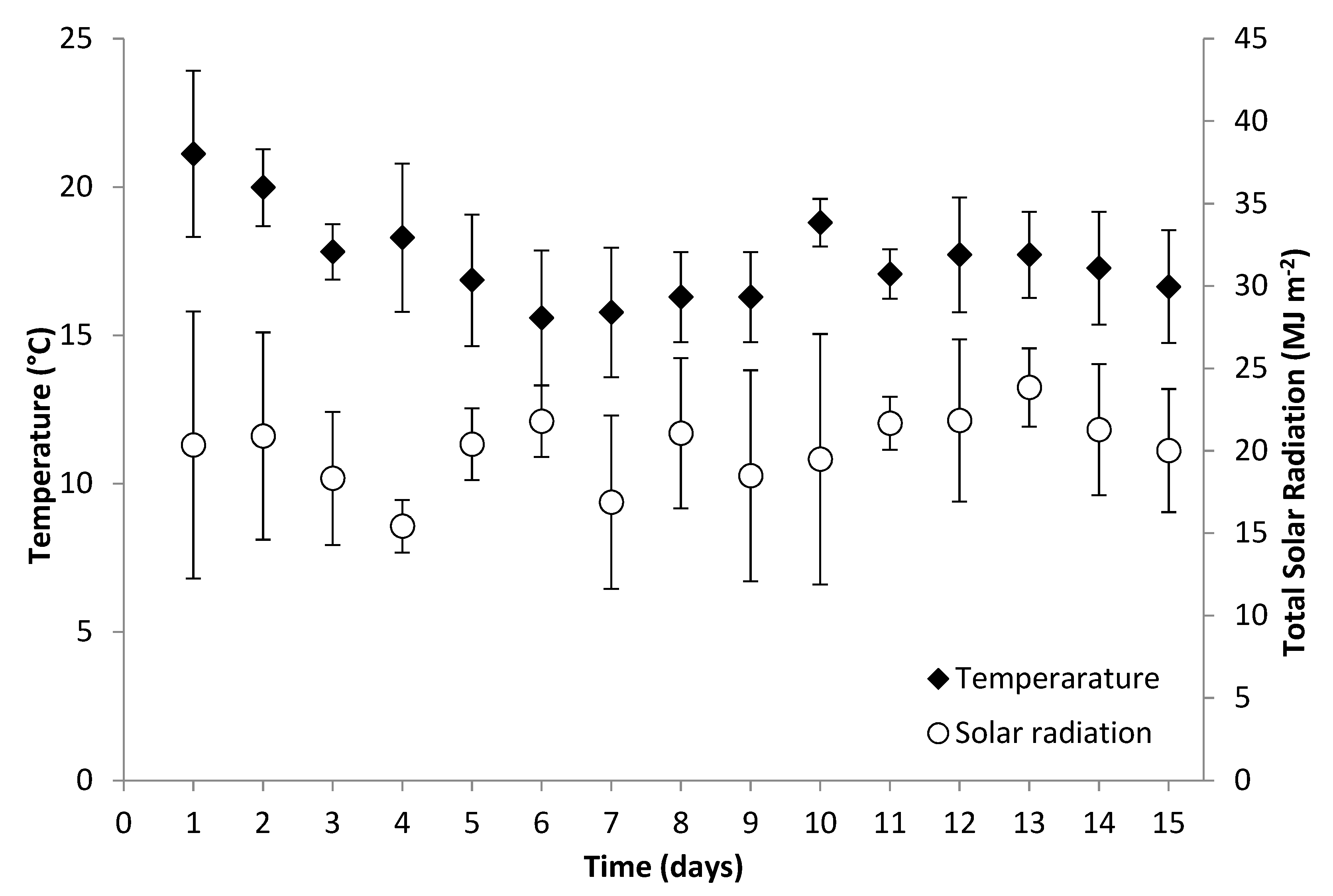
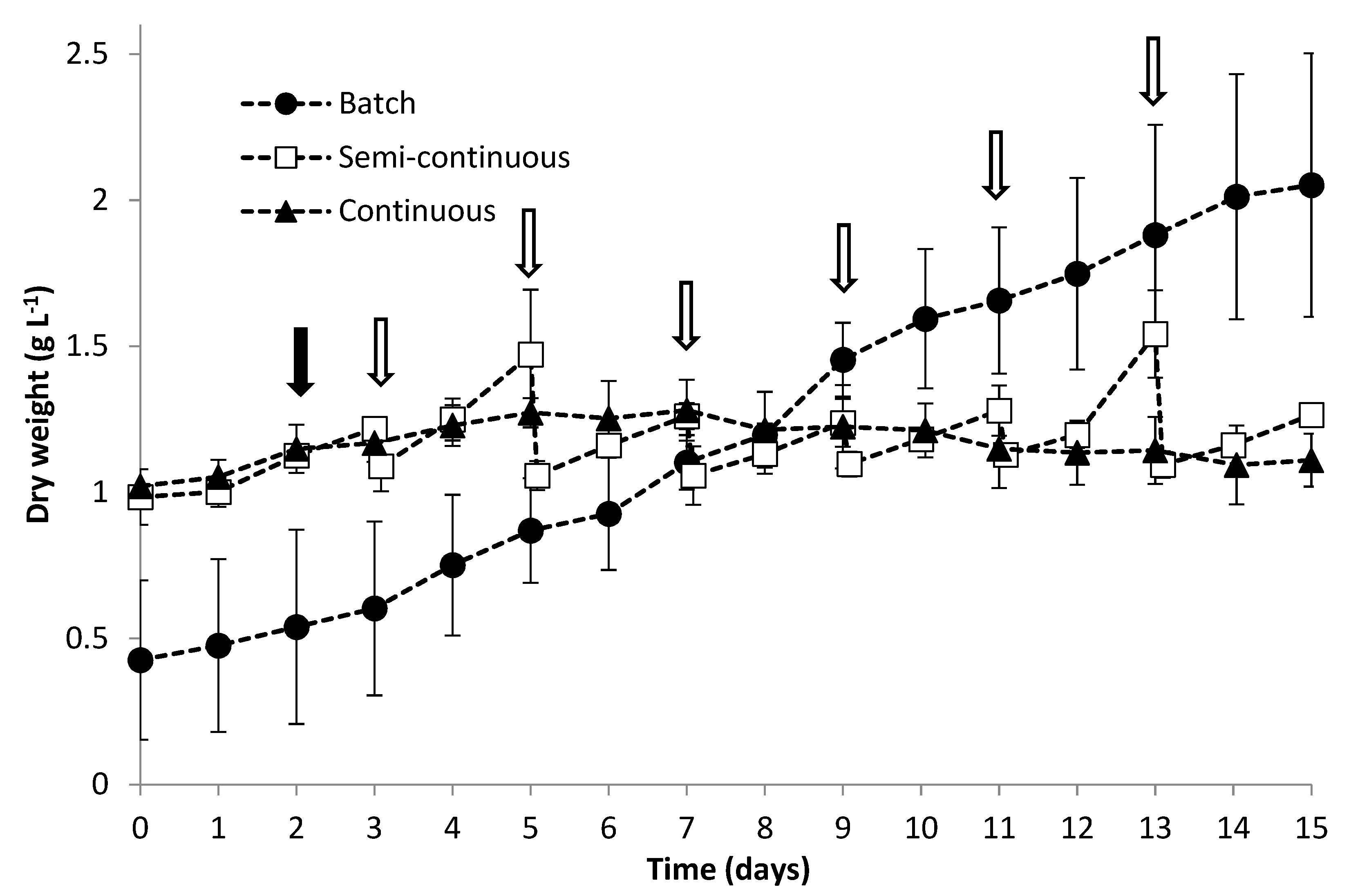
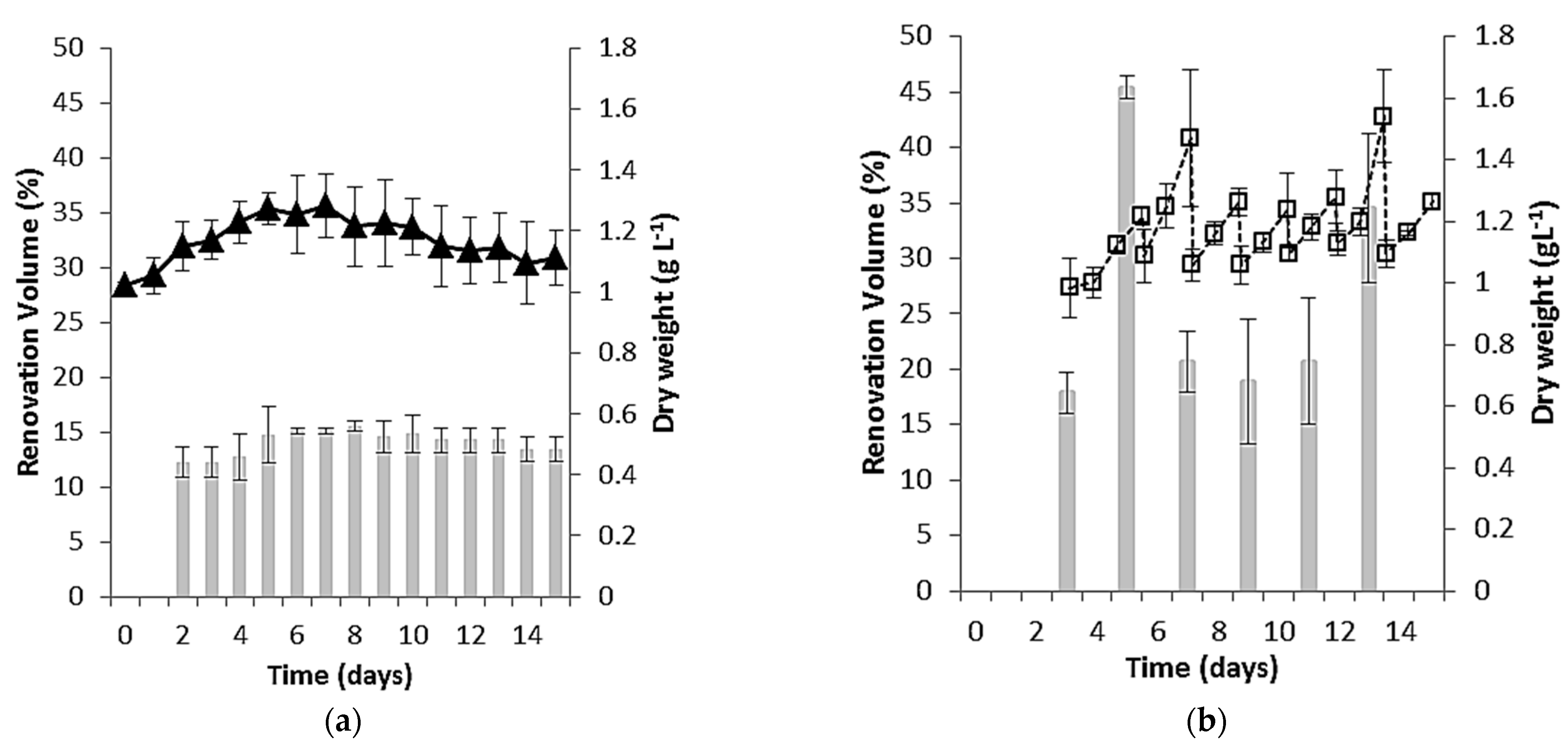
3.1. Growth Performance
3.2. Biochemical Profile
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- IPCC. Climate Change 2014: Synthesys Report. Contribution of Working Groups I, II and III to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Core Writing Team, Pachauri, R.K., Meyer, L.A., Eds.; IPCC: Geneva, Switzerland, 2015. [Google Scholar]
- Sajjadi, B.; Chen, W.Y.; Raman, A.A.A.; Ibrahim, S. Microalgae lipid and biomass for biofuel production: A comprehensive review on lipid enhancement strategies and their effects on fatty acid composition. Renew. Sustain. Energy Rev. 2018, 97, 200–232. [Google Scholar] [CrossRef]
- Rastogi, R.P.; Madamwar, D.; Pandey, A. Algal Green Chemistry: Recent Progress in Biotechnology; Elsevier: Amsterdam, The Netherlands, 2017; ISBN 9780444640413. [Google Scholar]
- Pires, J.C.M.; Alvim-Ferraz, M.C.M.; Martins, F.G.; Simões, M. Carbon dioxide capture from flue gases using microalgae: Engineering aspects and biorefinery concept. Renew. Sustain. Energy Rev. 2012, 16, 3043–3053. [Google Scholar] [CrossRef]
- Salama, E.S.; Kurade, M.B.; Abou-Shanab, R.A.I.; El-Dalatony, M.M.; Yang, I.S.; Min, B.; Jeon, B.H. Recent progress in microalgal biomass production coupled with wastewater treatment for biofuel generation. Renew. Sustain. Energy Rev. 2017, 79, 1189–1211. [Google Scholar] [CrossRef]
- Gupta, P.L.; Lee, S.M.; Choi, H.J. A mini review: Photobioreactors for large scale algal cultivation. World J. Microbiol. Biotechnol. 2015, 31, 1409–1417. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Wang, Z.; Yu, C.; Yin, Y.; Zhou, G. Evaluation of the potential of 9 Nannochloropsis strains for biodiesel production. Bioresour. Technol. 2014, 167, 503–509. [Google Scholar] [CrossRef]
- Ashour, M.; Elshobary, M.E.; El-Shenody, R.; Kamil, A.W.; Abomohra, A.E.F. Evaluation of a native oleaginous marine microalga Nannochloropsis oceanica for dual use in biodiesel production and aquaculture feed. Biomass Bioenergy 2019, 120, 439–447. [Google Scholar] [CrossRef]
- Liu, J.; Song, Y.; Qiu, W. Oleaginous microalgae Nannochloropsis as a new model for biofuel production: Review & analysis. Renew. Sustain. Energy Rev. 2017, 72, 154–162. [Google Scholar]
- Richmond, A.; Hu, Q. Handbook of Microalgal Culture: Applied Phycology and Biotechnology: Second Edition; Blackwell Publishing: Hoboken, NJ, USA, 2004; ISBN 9781118567166. [Google Scholar]
- Pereira, H.; Gangadhar, K.N.; Schulze, P.S.C.; Santos, T.; De Sousa, C.B.; Schueler, L.M.; Custódio, L.; Malcata, F.X.; Gouveia, L.; Varela, J.C.S.; et al. Isolation of a euryhaline microalgal strain, Tetraselmis sp. CTP4, as a robust feedstock for biodiesel production. Sci. Rep. 2016, 6, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Kokkinos, N.; Lazaridou, A.; Stamatis, N.; Orfanidis, S.; Mitropoulos, A.C.; Christoforidis, A.; Nikolaou, N. Biodiesel production from selected microalgae strains and determination of its properties and combustion specific characteristics. J. Eng. Sci. Technol. Rev. 2015, 8, 1–6. [Google Scholar] [CrossRef]
- Borowitzka, M.A.; Moheimani, N.R. Sustainable biofuels from algae. Mitig. Adapt. Strateg. Glob. Chang. 2013, 18, 13–25. [Google Scholar] [CrossRef]
- Cai, T.; Park, S.Y.; Racharaks, R.; Li, Y. Cultivation of Nannochloropsis salina using anaerobic digestion effluent as a nutrient source for biofuel production. Appl. Energy 2013, 108, 486–492. [Google Scholar] [CrossRef]
- San Pedro, A.; Gonzalez-Lopez, C.V.; Acien, F.G.; Molina-Grima, E. Outdoor pilot-scale production of Nannochloropsis gaditana: Influence of culture parameters and lipid production rates in tubular photobioreactors. Bioresour. Technol. 2014, 169, 667–676. [Google Scholar] [CrossRef] [PubMed]
- Benvenuti, G.; Bosma, R.; Ji, F.; Lamers, P.; Barbosa, M.J.; Wijffels, R.H. Batch and semi-continuous microalgal TAG production in lab-scale and outdoor photobioreactors. J. Appl. Phycol. 2016, 28, 3167–3177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ganesan, R.; Manigandan, S.; Samuel, M.S.; Shanmuganathan, R.; Brindhadevi, K.; Lan Chi, N.T.; Duc, P.A.; Pugazhendhi, A. A review on prospective production of biofuel from microalgae. Biotechnol. Rep. 2020, 27, e00509. [Google Scholar] [CrossRef] [PubMed]
- Zohri, A.-N.A.; Ragab, S.W.; Mekawi, M.I.; Mostafa, O.A.A. Comparison between batch, fed-batch, semi-continuous and continuous techniques for bio-ethanol production from a mixture of egyptian cane and beet molasses. Egypt. Sugar J. 2017, 9, 89–111. [Google Scholar]
- Lavens, P.; Sorgeloos, P.; Food and Agriculture Organization of the United Nations. Microalgae. In Manual on the Production and Use of Live Food for Aquaculture; Food and Agriculture Organization of the United Nations: Roma, Italy, 1996; p. 295. ISBN 9251039348. [Google Scholar]
- Brown, M.R.; Garland, C.D.; Jeffrey, S.W.; Jameson, I.D.; Leroi, J.M. The gross and amino acid compositions of batch and semi-continuous cultures of Isochrysis sp. (clone T.ISO), Pavlova lutheri and Nannochloropsis oculata. J. Appl. Phycol. 1993, 5, 285–296. [Google Scholar] [CrossRef]
- Croughan, M.S.; Konstantinov, K.B.; Cooney, C. The future of industrial bioprocessing: Batch or continuous? Biotechnol. Bioeng. 2015, 112, 648–651. [Google Scholar] [CrossRef] [PubMed]
- Bux, F. Biotechnological Applications of Microalgae: Biodiesel and Value-Added Products; CRC Press: Boca Raton, FL, USA, 2013; ISBN 9781466515307. [Google Scholar]
- Wang, B.; Lan, C.Q.; Horsman, M. Closed photobioreactors for production of microalgal biomasses. Biotechnol. Adv. 2012, 30, 904–912. [Google Scholar] [CrossRef]
- Mata, T.M.; Martins, A.A.; Caetano, N.S. Microalgae for biodiesel production and other applications: A review. Renew. Sustain. Energy Rev. 2010, 14, 217–232. [Google Scholar] [CrossRef] [Green Version]
- Callejón-Ferre, A.J.; Velázquez-Martí, B.; López-Martínez, J.A.; Manzano-Agugliaro, F. Greenhouse crop residues: Energy potential and models for the prediction of their higher heating value. Renew. Sustain. Energy Rev. 2011, 15, 948–955. [Google Scholar] [CrossRef]
- Armstrong, F.A.J. Determination of Nitrate in Water by Ultraviolet Spectrophotometry. Anal. Chem. 1963, 35, 1292–1294. [Google Scholar] [CrossRef]
- Bligh, E.G.; Dyer, W.J. A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef] [Green Version]
- Guedes, A.C.; Barbosa, C.R.; Amaro, H.M.; Pereira, C.I.; Malcata, F.X. Microalgal and cyanobacterial cell extracts for use as natural antibacterial additives against food pathogens. Int. J. Food Sci. Technol. 2011, 46, 862–870. [Google Scholar] [CrossRef]
- Lepage, G.; Roy, C.C. Improved recovery of fatty acid through direct transesterification without prior extraction or purification. J. Lipid Res. 1984, 25, 1391–1396. [Google Scholar] [CrossRef]
- Pereira, H.; Barreira, L.; Mozes, A.; Florindo, C.; Polo, C.; Duarte, C.V.; Custádio, L.; Varela, J. Microplate-based high throughput screening procedure for the isolation of lipid-rich marine microalgae. Biotechnol. Biofuels 2011, 4, 61. [Google Scholar] [CrossRef] [Green Version]
- Quinn, J.C.; Yates, T.; Douglas, N.; Weyer, K.; Butler, J.; Bradley, T.H.; Lammers, P.J. Nannochloropsis production metrics in a scalable outdoor photobioreactor for commercial applications. Bioresour. Technol. 2012, 117, 164–171. [Google Scholar] [CrossRef] [PubMed]
- Ledda, C.; Romero Villegas, G.I.; Adani, F.; Acién Fernández, F.G.; Molina Grima, E. Utilization of centrate from wastewater treatment for the outdoor production of Nannochloropsis gaditana biomass at pilot-scale. Algal Res. 2015, 12, 17–25. [Google Scholar] [CrossRef]
- Chini Zittelli, G.; Lavista, F.; Bastianini, A.; Rodolfi, L.; Vincenzini, M.; Tredici, M.R. Production of eicosapentaenoic acid by Nannochloropsis sp. cultures in outdoor tubular photobioreactors. Prog. Ind. Microbiol. 1999, 35, 299–312. [Google Scholar]
- Camacho-Rodríguez, J.; González-Céspedes, A.M.; Cerón-García, M.C.; Fernández-Sevilla, J.M.; Acién-Fernández, F.G.; Molina-Grima, E. A quantitative study of eicosapentaenoic acid (EPA) production by Nannochloropsis gaditana for aquaculture as a function of dilution rate, temperature and average irradiance. Appl. Microbiol. Biotechnol. 2014, 98, 2429–2440. [Google Scholar] [CrossRef]
- De Vree, J.H.; Bosma, R.; Janssen, M.; Barbosa, M.J.; Wijffels, R.H. Comparison of four outdoor pilot-scale photobioreactors. Biotechnol. Biofuels 2015, 8, 215. [Google Scholar] [CrossRef] [Green Version]
- Zhang, D.; Xue, S.; Sun, Z.; Liang, K.; Wang, L.; Zhang, Q.; Cong, W. Investigation of continuous-batch mode of two-stage culture of Nannochloropsis sp. for lipid production. Bioprocess Biosyst. Eng. 2014, 37, 2073–2082. [Google Scholar] [CrossRef] [PubMed]
- Nogueira, N.; Nascimento, F.J.A.; Cunha, C.; Cordeiro, N. Nannochloropsis gaditana grown outdoors in annular photobioreactors: Operation strategies. Algal Res. 2020, 48, 101913. [Google Scholar] [CrossRef]
- Fakhry, E.M.; El Maghraby, D.M. Lipid accumulation in response to nitrogen limitation and variation of temperature in nannochloropsis salina. Bot. Stud. 2015, 56, 6. [Google Scholar] [CrossRef] [Green Version]
- Carneiro, M.; Cicchi, B.; Maia, I.B.; Pereira, H.; Zittelli, G.C.; Varela, J.; Malcata, F.X.; Torzillo, G. Effect of temperature on growth, photosynthesis and biochemical composition of Nannochloropsis oceanica, grown outdoors in tubular photobioreactors. Algal Res. 2020, 49, 101923. [Google Scholar] [CrossRef]
- Cunha, P.; Pereira, H.; Costa, M.; Pereira, J.; Silva, J.T.; Fernandes, N.; Varela, J.; Silva, J.; Simões, M. Nannochloropsis oceanica Cultivation in Pilot-Scale Raceway Ponds—From Design to Cultivation. Appl. Sci. 2020, 10, 1725. [Google Scholar] [CrossRef] [Green Version]
- Golimowski, W.; Berger, W.A.; Pasyniuk, P.; Rzeźnik, W.; Czechlowski, M.; Koniuszy, A. Biofuel parameter dependence on waste fats’ fatty acids profile. Fuel 2017, 197, 482–487. [Google Scholar] [CrossRef]
- Wu, H.; Miao, X. Biodiesel quality and biochemical changes of microalgae Chlorella pyrenoidosa and Scenedesmus obliquus in response to nitrate levels. Bioresour. Technol. 2014, 170, 421–427. [Google Scholar] [CrossRef]
- Chen, L.; Liu, T.; Zhang, W.; Chen, X.; Wang, J. Biodiesel production from algae oil high in free fatty acids by two-step catalytic conversion. Bioresour. Technol. 2012, 111, 208–214. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Production Regime | Specific Growth Rate (Day−1) | Water (m3/Cycle) | Water (m3 kg−1 Produced Biomass) | Average Dilution Rate (Day−1) |
|---|---|---|---|---|
| Batch | 0.129 ± 0.020 | 2.6 ± 0.0 a | 0.62 ± 0.07 a | - |
| Semi-continuous | - | 6.7 ± 0.4 ab | 1.04 ± 0.03 b | 0.122 ± 0.011 a |
| Continuous | - | 7.4 ± 0.1 b | 1.25 ± 0.14 b | 0.140 ± 0.010 a |
| Production Regime | Volumetric Productivity (g L−1 day−1) | Maximum Volumetric Productivity (g L−1 day−1) | Areal Productivity (g m2 day−1) | Maximum Areal Productivity (g m2 day−1) | Photosynthetic Efficiency (%) |
|---|---|---|---|---|---|
| Batch | 0.108 ± 0.011 a | 0.333 ± 0.03 a | 10.7 ± 1.1 a | 31.6 ± 3.4 a | 0.358 ± 0.016 a |
| Semi-continuous | 0.165 ± 0.013 b | 0.427 ± 0.020 b | 16.3 ± 1.3 b | 40.4 ± 1.9 b | 0.436 ± 0.043 a |
| Continuous | 0.154 ± 0.021 b | 0.266 ± 0.028 a | 15.2 ± 2.0 b | 25.2 ± 2.6 a | 0.481 ± 0.073 a |
| Production Regime | Protein (%) | Lipids (%) | Ash (%) | Carbohydrates (%) | ||||
|---|---|---|---|---|---|---|---|---|
| Average | Min | Max | Average | Min | Max | |||
| Batch | 29.6 ± 3.6 a | 22.0 ± 3.1 a | 10.7 | 9.4 | 11.9 | 37.8 | 34.7 | 40.8 |
| Semi-continuous | 28.6 ± 2.8 a | 24.0 ± 5.3 a | 14.9 | 13.0 | 16.8 | 32.5 | 26.4 | 38.6 |
| Continuous | 28.9 ± 2.5 a | 19.1 ± 2.5 a | 12.9 | 12.0 | 13.8 | 39.1 | 36.4 | 41.8 |
| Production Regime | Lipid Productivity (mg L−1 day−1) |
|---|---|
| Batch | 22.9 ± 3.7 a |
| Semi-continuous | 39.6 ± 3.1 b |
| Continuous | 29.5 ± 3.9 ab |
| FAME | Batch | Semi-Continuous | Continuous |
|---|---|---|---|
| C 14:0 (%) | 6.88 ± 0.44 a | 5.99 ± 0.66 a | 6.02 ± 0.39 a |
| C 16:1 (%) | 32.79 ± 0.55 a | 32.28 ± 0.65 a | 32.28 ± 1.79 a |
| C 16:0 (%) | 33.58 ± 1.76 a | 38.24 ± 2.10 b | 36.20 ± 0.94 ab |
| C 18:2 ω6 (%) | 0.75 ± 0.09 a | 0.59 ± 0.59 b | 0.76 ± 0.08 a |
| C 18:1 (%) | 11.96 ± 2.98 a | 11.07 ± 0.76 a | 12.40 ± 2.46 a |
| C 18:0 (%) | 1.05 ± 0.26 a | 1.95 ± 0.26 b | 1.59 ± 0.19 ab |
| C 20:4 ω6 (%) | 1.85 ± 0.48 a | 1.42 ± 0.30 a | 1.46 ± 0.11 a |
| C 20:5 ω3 (%) | 11.12 ± 3.4 a | 8.44 ± 2.00 a | 9.16 ± 0.08 a |
| SFA (%) | 41.52 ± 1.60 a | 46.19 ± 1.64 b | 43.80 ± 1.40 ab |
| MUFA (%) | 44.76 ± 2.43 a | 43.36 ± 0.92 a | 44.68 ± 067 a |
| PUFA (%) | 13.73 ± 1.00 a | 10.45 ± 2.31 a | 11.52 ± 0.88 a |
| PUFA/SFA | 0.33 ± 0.11 a | 0.23 ± 0.06 a | 0.26 ± 0.03 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guerra, I.; Pereira, H.; Costa, M.; Silva, J.T.; Santos, T.; Varela, J.; Mateus, M.; Silva, J. Operation Regimes: A Comparison Based on Nannochloropsis oceanica Biomass and Lipid Productivity. Energies 2021, 14, 1542. https://doi.org/10.3390/en14061542
Guerra I, Pereira H, Costa M, Silva JT, Santos T, Varela J, Mateus M, Silva J. Operation Regimes: A Comparison Based on Nannochloropsis oceanica Biomass and Lipid Productivity. Energies. 2021; 14(6):1542. https://doi.org/10.3390/en14061542
Chicago/Turabian StyleGuerra, Inês, Hugo Pereira, Margarida Costa, Joana T. Silva, Tamára Santos, João Varela, Marília Mateus, and Joana Silva. 2021. "Operation Regimes: A Comparison Based on Nannochloropsis oceanica Biomass and Lipid Productivity" Energies 14, no. 6: 1542. https://doi.org/10.3390/en14061542
APA StyleGuerra, I., Pereira, H., Costa, M., Silva, J. T., Santos, T., Varela, J., Mateus, M., & Silva, J. (2021). Operation Regimes: A Comparison Based on Nannochloropsis oceanica Biomass and Lipid Productivity. Energies, 14(6), 1542. https://doi.org/10.3390/en14061542