

New Electrogenic Microorganism Citrobacter sp. Isolated from Microbial Fuel Cell and Bacterial Characteristics Determination
 , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
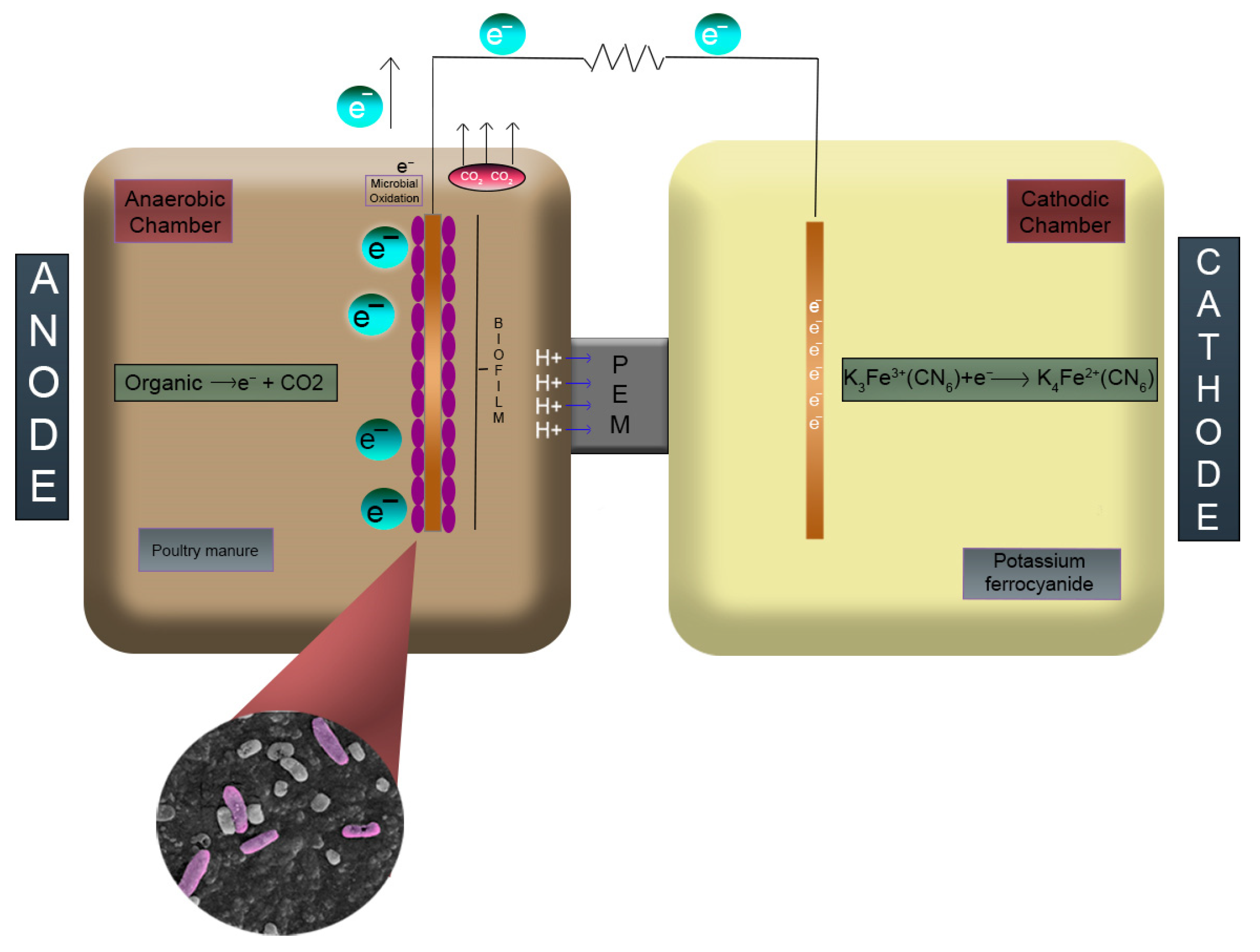
2.1. MFC Construction and Operation
2.2. Electrochemical Configuration of MFC
2.3. Substrate Obtention and Preparation
2.4. Physico-Chemical Parameters
2.5. Inoculum Development and Strain Isolation of Av_G1
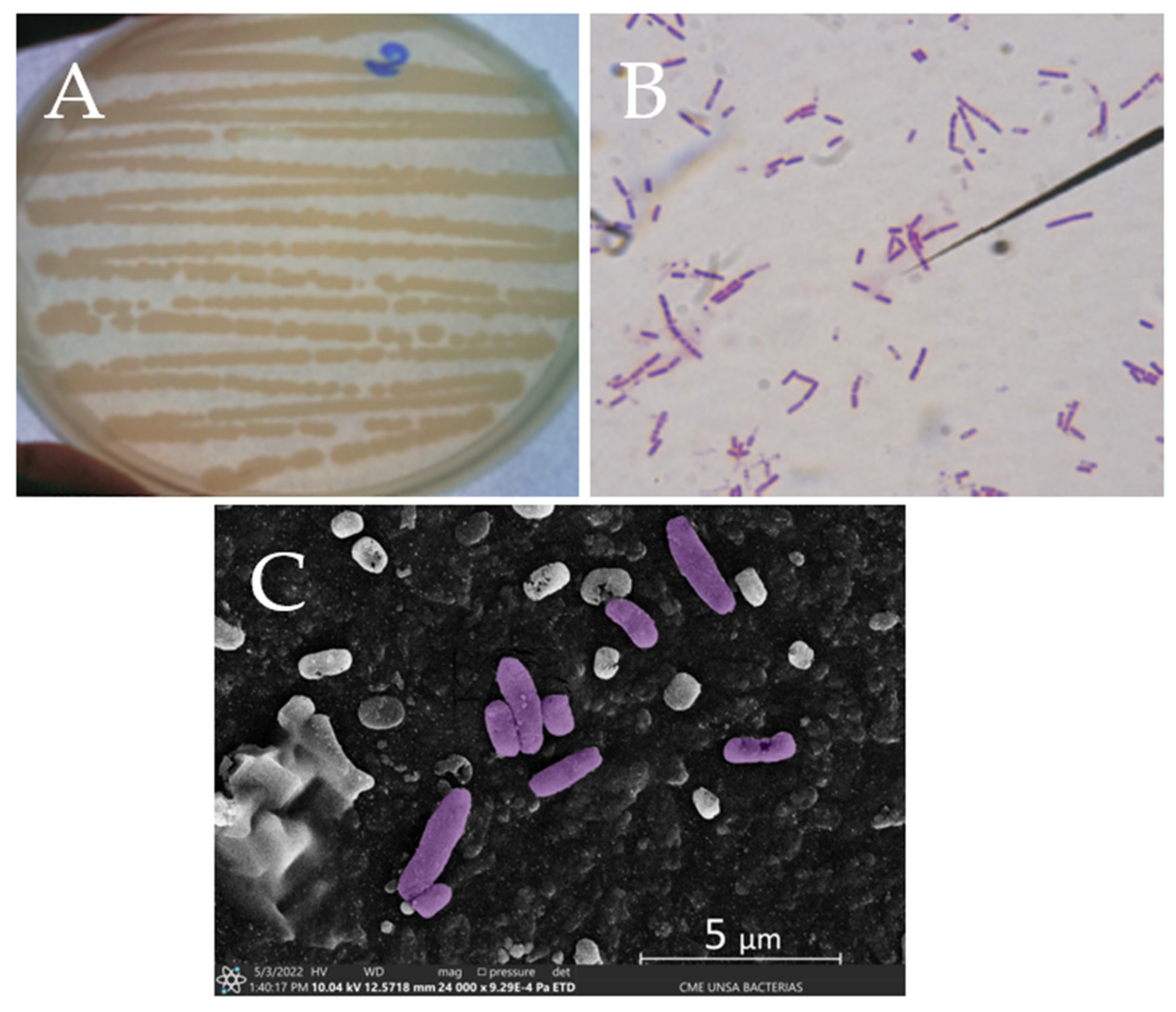
2.6. Morphological Characterization and Microscopic Analysis
2.7. Gene Sequencing for 16rDNA
2.7.1. Lysis and DNA Extraction
2.7.2. DNA Amplification
2.7.3. Electrophoresis and DNA Purification
2.8. Bioinformatic and Phylogeny Analysis
3. Results
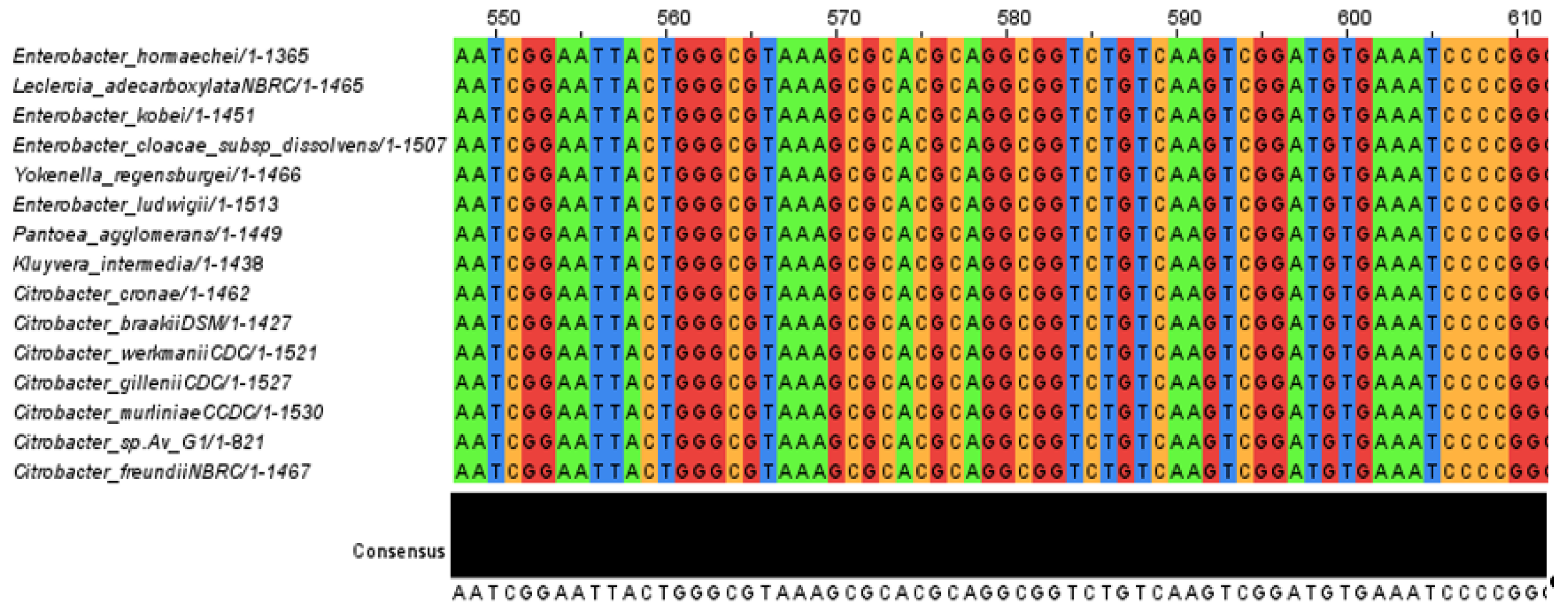
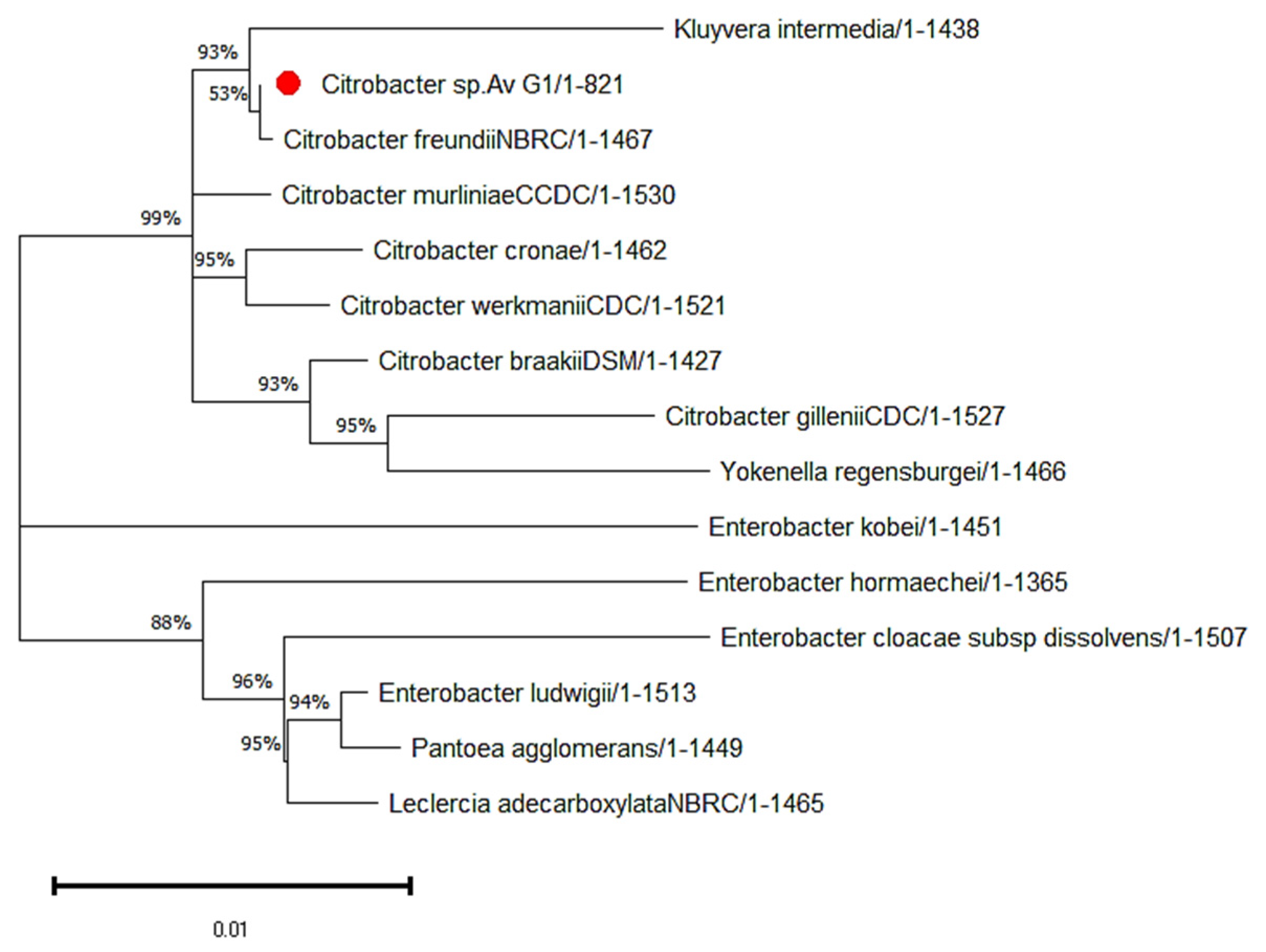
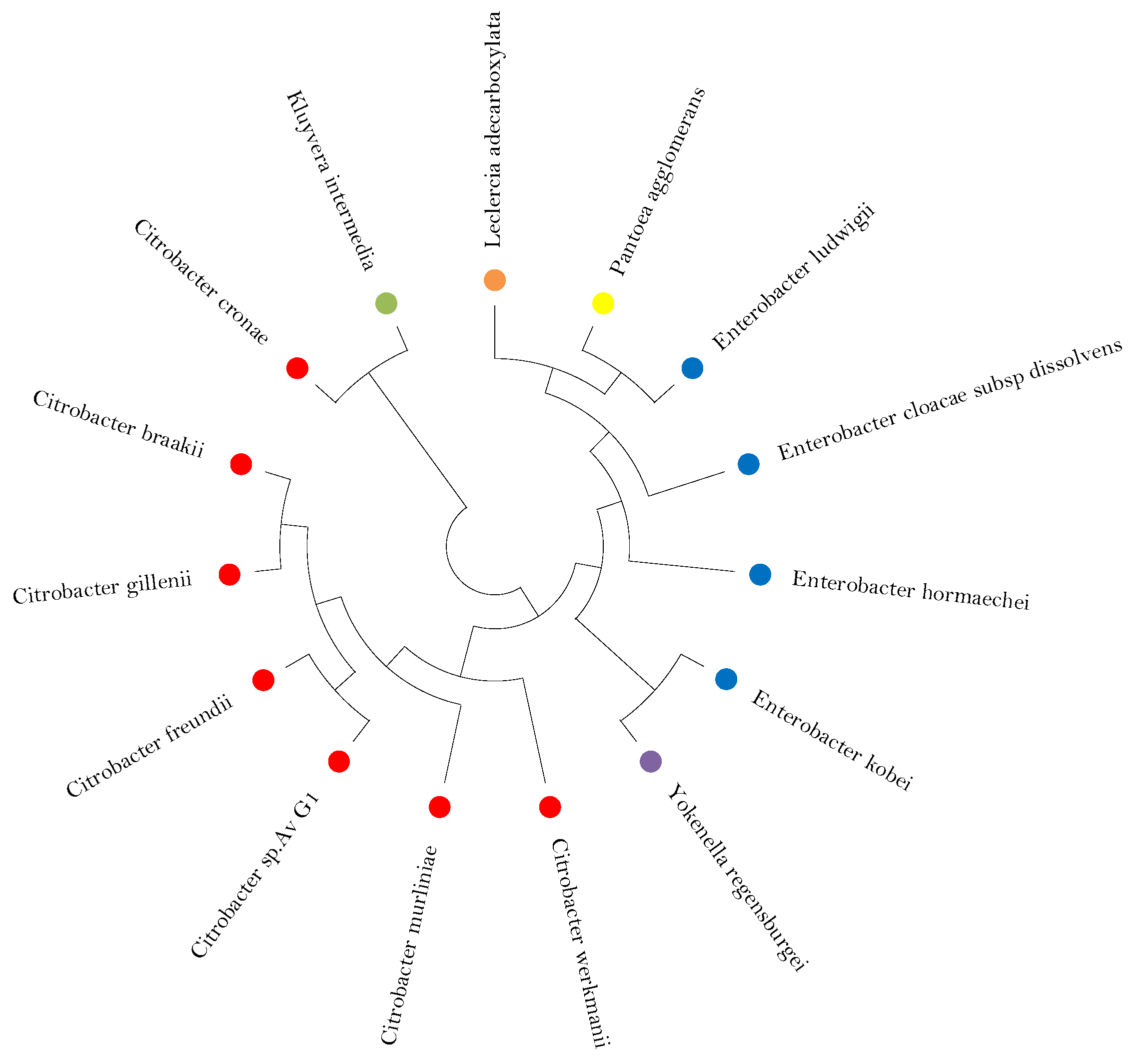
3.1. Phylogeny Analysis and Sequence of the Isolated Strain Av_G1
3.2. Characterization of Citrobacter sp. Av_G1
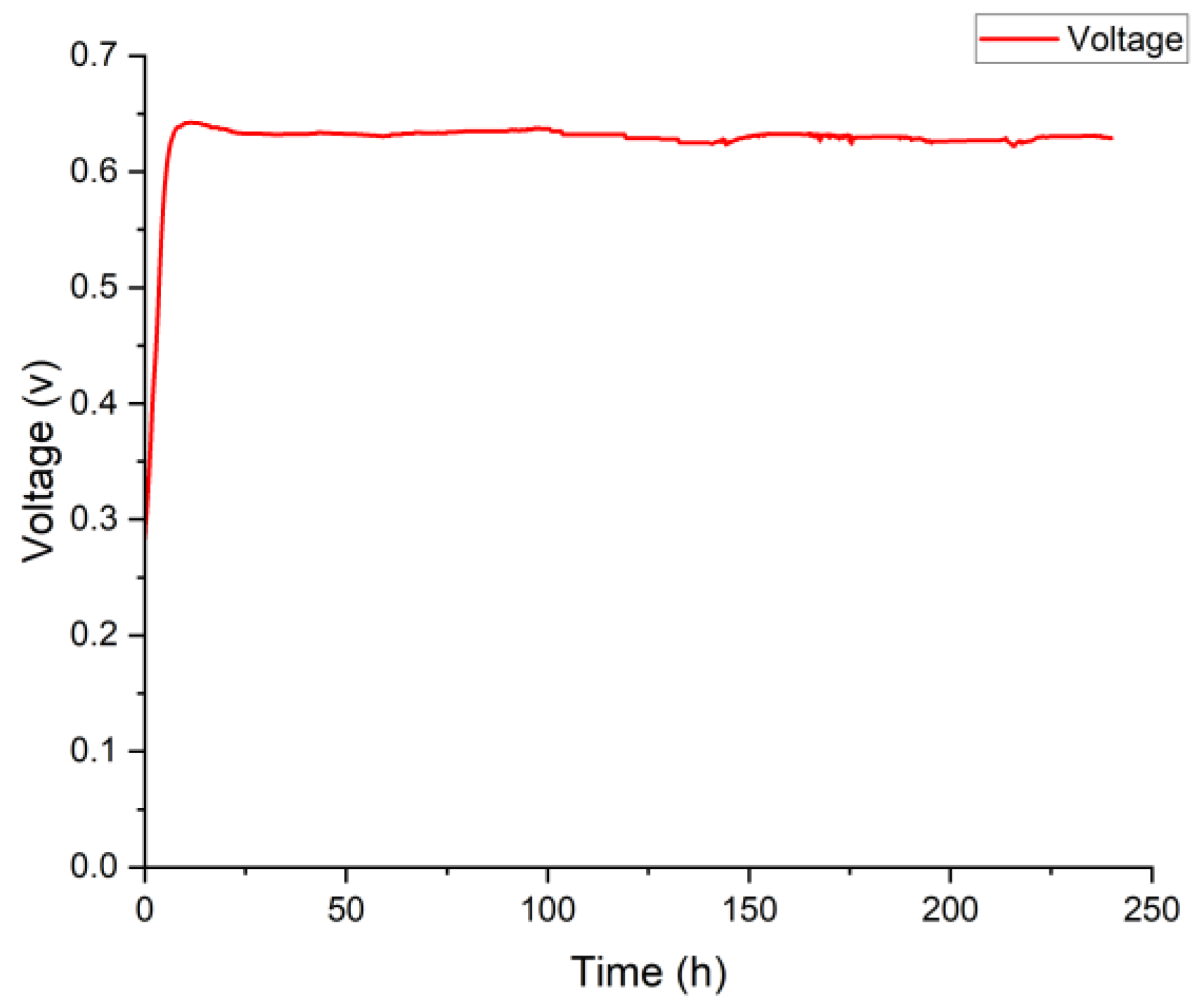
3.3. Current Generation of Citrobacter sp. Av_G1 in MFC
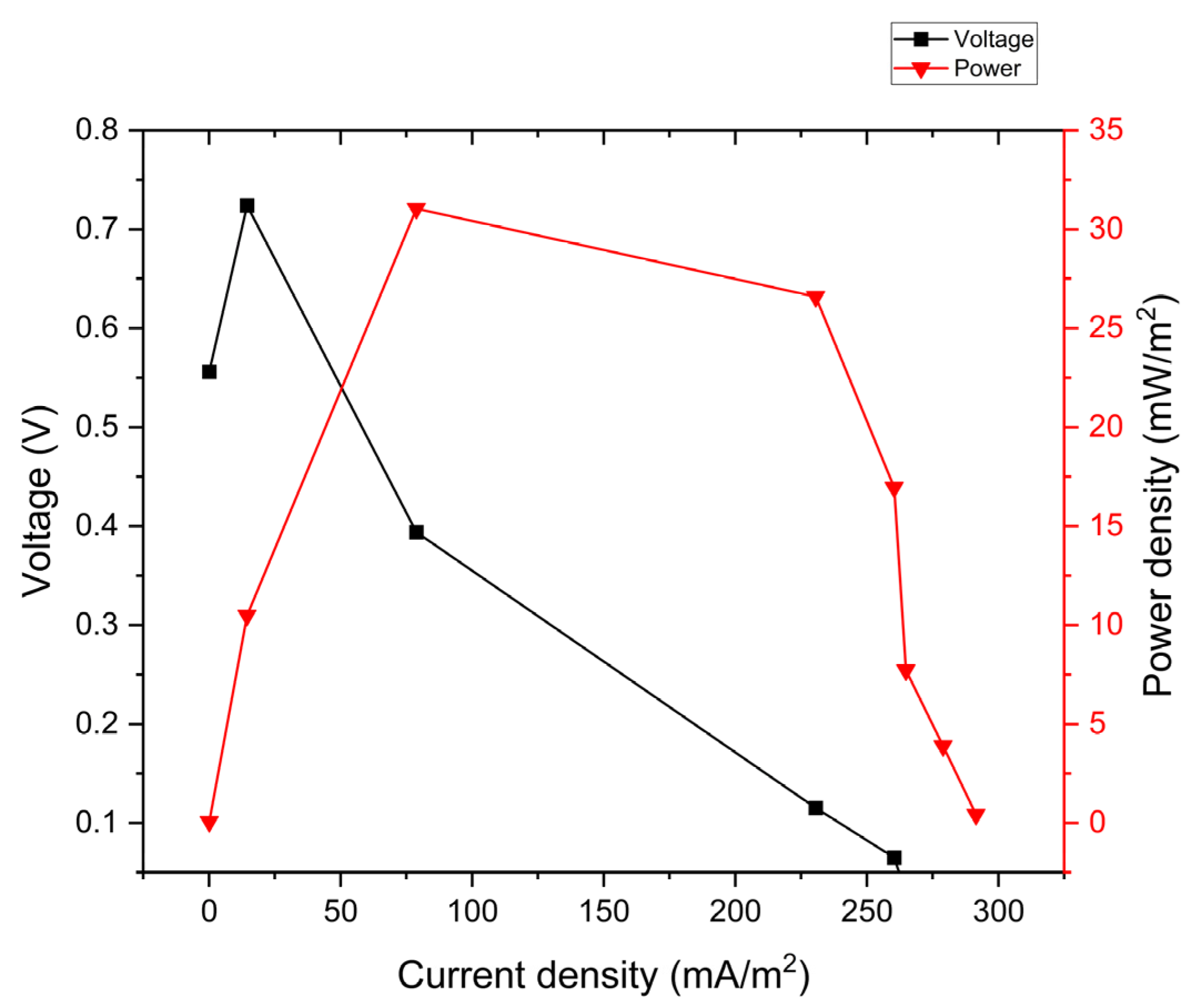
3.4. Polarization Curves and Power Density
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lorenczik, S.; Jorquera, J.; Hevia-Coch, P.; Hungerford, Z. Electricity Market Report Home Page. Available online: https://iea.blob.core.windows.net/assets/d75d928b-9448-4c9b-b13d6a92145af5a3/ElectricityMarketReport_January2022.pdf (accessed on 23 November 2022).
- Ochs, A.; Killeen, P.; Majano, A.M. Advancing Bioelectricity in Latin America and the Caribbean. Proceedings and Outcomes of the First Implementation Year of a Community of Practice of Leading Countries in the Region; Resilient and Low Emission Development Strategies: San José, Costa Rica, 2017. [Google Scholar]
- Bathia, S.K.; Joo, H.S.; Yang, Y.H. Biowaste-to-bioenergy using biomedical methods—A mini-review. Energy Convers. Manag. 2018, 177, 640–660. [Google Scholar] [CrossRef]
- Moqsud, M.A.; Bushra, Q.S.; Rahman, M.H. Composting barrel for sustainable organic waste management in Bangladesh. Waste Manag. Res. 2011, 29, 1286–1293. [Google Scholar] [CrossRef] [Green Version]
- Varanasi, J.L.; Das, D. Microbial Fuel Cell; Springer: Kharagpur, India, 2017; Chapter 2; pp. 43–62. ISBN 978-3-319-66793-5. [Google Scholar]
- Flimban, S.G.; Ismail, I.M.; Kim, T.; Oh, S. Overview of Recent Advancements in the Microbial Fuel Cell from Fundamentals to Applications: Design, Major Elements, and Scalability. Energies 2019, 12, 3390. [Google Scholar] [CrossRef] [Green Version]
- Santoro, C.; Arbizzani, C.; Erable, B.; Leropoulos, L. Microbial fuel cells: From fundamentals to applications. A review. J. Power Sources 2017, 356, 225–244. [Google Scholar] [CrossRef] [PubMed]
- Fadzli, F.; Bhawani, S.; Mohammad, R.E. Microbial Fuel Cell: Recent Developments in Organic Substrate Use and Bacterial Electrode Interaction. J. Chem. 2021, 2021, 4570388. [Google Scholar] [CrossRef]
- Logan, B.E.; Rabaey, K. Conversion of wastes into bioelectricity and chemicals by using microbial electrochemical technologies. Science 2012, 337, 686–690. [Google Scholar] [CrossRef] [Green Version]
- Saha, T.; Protity, A.; Zohora, F.; Shaha, M.; Ahmed, I.; Barua, E.; Sarker, P.; Mukherjee, S.; Barua, A.; Salimullah, M.; et al. Microbial Fuel Cell (MFC) Application for Generation of Electricity from Dumping Rubbish and Identification of Potential Electrogenic Bacteria. Adv. Ind. Biotechnol. 2019, 2, 1–8. [Google Scholar] [CrossRef]
- Chabert, N.; Ali, O.; Achouak, W. All ecosystems potentially host electrogenic bacteria. Bioelectrochemistry 2015, 106, 88–96. [Google Scholar] [CrossRef] [PubMed]
- Reguera, G.; Kashefi, K. The electrifying physiology of Geobacter bacteria, 30 years on. Adv. Microb. Physiol. 2019, 74, 1–96. [Google Scholar] [CrossRef] [PubMed]
- Yang, T.; Coppi, M.; Lovley, D.; Sun, J. Metabolic response of Geobacter sulfurreducens towards electron donor/acceptor variation. Microb. Cell Factories 2010, 9, 90. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Zhou, M.; Tian, X.; Tan, C.; McDaniel, C.; Hassett, D.; Gu, T. Microbial fuel cell (MFC) power performance improvement through enhanced microbial electrogenicity. Biotechnol. Adv. 2018, 36, 1316–1327. [Google Scholar] [CrossRef]
- Sacco, N.; Bonetto, M.; Cortón, E. Isolation and Characterization of a Novel Electrogenic Bacterium, Dietzia sp. RNV-4. PLoS ONE 2017, 12, e0169955. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Logan, B.; Rossi, R.; Ragab, A.; Saikaly, P. Electroactive microorganisms in bioelectrochemical systems. Nat. Rev. Microbiol. 2019, 17, 307–319. [Google Scholar] [CrossRef] [PubMed]
- Guang, L.; Koomson, D.; Jingyu, H.; Ewusi-Mensah, D.; Miwornunyuie, N. Performance of Exoelectrogenic Bacteria Used in Microbial Desalination Cell Technology. Int. J. Environ. Res. Public Health 2020, 17, 1121. [Google Scholar] [CrossRef] [Green Version]
- Karthikeyan, R.; Singh, R.; Bose, A. Microbial electron uptake in microbial electrosynthesis: A mini-review. J. Ind. Microbiol. Biotechnol. 2019, 46, 1419–1426. [Google Scholar] [CrossRef]
- Ng, C.; Xu, J.; Cai, Z.; Yang, L.; Thompson, I.; Huang, W.; Cao, B. Elevated intracellular cyclic-di-GMP level in Shewanella oneidensis increases expression of c-type cytochromes. Microb. Biotechnol. 2020, 13, 1904–1916. [Google Scholar] [CrossRef] [PubMed]
- Mostafa, G.; Ramli, W.D.; Ahmad, F.I.; Yaghoob, J.; Manal, I.; Alireza, M.; Othman, J. Simultaneous wastewater treatment and electricity generation by microbial fuel cell: Performance comparison and cost investigation of using Nafion 117 and SPEEK as separators. Desalination 2013, 325, 1–6. [Google Scholar] [CrossRef]
- Wei, L.; Han, H.; Shen, J. Effects of cathodic electron acceptors and potassium ferricyanide concentrations on the performance of microbial fuel cell. Int. J. Hydrog. Energy 2012, 37, 12980–12986. [Google Scholar] [CrossRef]
- Fraiwan, A.; Choi, S. A stackable, two-chambered, paper-based microbial fuel cell. Biosens. Bioelectron. 2016, 83, 27–32. [Google Scholar] [CrossRef] [Green Version]
- Yu, D.; Bai, L.; Zhai, J.; Wang, Y.; Dong, S. Toxicity detection in water containing heavy metal ions with a self-powered microbial fuel cell-based biosensor. Talanta 2017, 168, 210–216. [Google Scholar] [CrossRef] [PubMed]
- Naik, S.; Jujjavarappu, S.E. Simultaneous bioelectricity generation from cost-effective MFC and water treatment using various wastewater samples. Environ. Sci. Pollut. Res. 2020, 27, 27383–27393. [Google Scholar] [CrossRef]
- ASTM-D 4547: 2009; Standard Guide for Sampling Waste and Soils for Volatile Organic Compounds. American Society for Testing and Materials: Philadelphia, PA, USA, 2009. Available online: https://infostore.saiglobal.com/en-us/standards/astm-d-4547-2009-149061_saig_astm_astm_344953/ (accessed on 12 October 2022).
- Kim, J.R.; Min, B.; Logan, B.E. Evaluation of procedures to acclimate a microbial fuel cell for electricity production. Appl. Microbiol. Biotechnol. 2005, 68, 23–30. [Google Scholar] [CrossRef] [PubMed]
- Yuvraj, C.; Aranganathan, V. MFC—An Approach in Enhancing Electricity Generation Using Electroactive Biofilm of Dissimilatory Iron-Reducing (DIR) Bacteria. Arab. J. Sci. Eng. 2017, 42, 2341–23477. [Google Scholar] [CrossRef]
- Xiao, K.; Wang, K.; Liu, C.; Wang, M.; Dong, W.; Pan, T. Influence of Triton X-100 and β-cyclodextrin on the bioavailability and biodegradation of crystalline phenanthrene covered with biofilms. Process Biochem. 2021, in press. [CrossRef]
- Speers, A.M.; Colloggi, D.L.; Reguera, G. Anaerobic Cell Culture. Curr. Protoc. Microbiol. 2009, 12, A.4F.1–A.4F.16. [Google Scholar] [CrossRef]
- Smith, A.; Hussey, M. Gram Stain Protocols Home Page. Available online: https://asm.org/getattachment/5c95a063-326b-4b2f-98ce-001de9a5ece3/gram-stain-protocol-2886.pdf (accessed on 1 November 2022).
- Kammoun, R.; Zmantar, T.; Ghoul, S. Scanning electron microscopy approach to observe bacterial adhesion to dental surfaces. MethodsX 2020, 7, 101107. [Google Scholar] [CrossRef] [PubMed]
- Bazzicalupo, M.; Fancelli, S. DNA Extraction from Bacterial Cultures. In Fingerprinting Methods Based on Arbitrarily Primed PCR; Springer Lab Manuals; Micheli, M.R., Bova, R., Eds.; Springer: Berlin/Heidelberg, Germany, 1997. [Google Scholar] [CrossRef]
- Qiagen. QIAquick® Spin Handbook QIAGEN; Qiagen: Hilden, Germany, 2020; pp. 19–20. [Google Scholar]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naïve Bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Appl. Environ. Microbiol. 2007, 73, 5261–5267. [Google Scholar] [CrossRef] [Green Version]
- NCBI Resource Coordinators. Database resources of the National Center for Biotechnology Information. Nucleic Acids Res. 2016, 44, D7–D19. [Google Scholar] [CrossRef] [Green Version]
- McWilliam, H.; Li, W.; Uludag, M.; Squizzato, S.; Park, Y.M.; Buso, N.; Cowley, A.P.; Lopez, R. Analysis Tool Web Services from the EMBL-EBI. Nucleic Acids Res. 2013, 41, 597–600. [Google Scholar] [CrossRef] [Green Version]
- Clamp, M.; Cuff, J.; Searle, S.M.; Barton, G.J. The Jalview Java alignment editor. Bioinformatics 2004, 20, 426–427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Jenkins, C.; Ling, C.; Ciesielczuk, H.; Lockwood, J.; Hopkins, S.; McHugh, T.; Gillespie, S.; Kibbler, C. Detection and identification of bacteria in clinical samples by 16S rRNA gene sequencing: Comparison of two different approaches in clinical practice. J. Med. Microbiol. 2012, 61 Pt 4, 483–488. [Google Scholar] [CrossRef]
- Fazal, T.; Saif ur Rehman, M.; Mushtaq, A.; Hafeez, A.; Javed, F.; Aslam, M.; Fatima, M.; Faisal, A.; Iqbal, J.; Rehman, F.; et al. Simultaneous production of bioelectricity and biogas from chicken droppings and dairy industry wastewater employing bioelectrochemical system. Fuel 2019, 256, 115902. [Google Scholar] [CrossRef]
- Gazali, T.A.; Moqsud, M.A. The Effectiveness of Animal Dungs and Leaf Mold for Bioelectricity Generation Using Microbial Fuel Cell with Soils. J. Sustain. Bioenergy Syst. 2017, 4, 165–181. [Google Scholar] [CrossRef]
- Watanabe, K. Recent Developments in Microbial Fuel Cell Technologies for Sustainable Bioenergy. J. Biosci. Bioeng. 2008, 106, 528–536. [Google Scholar] [CrossRef]
- Pant, D.; van Bogaert, G.; Diels, L.; Vanbroekhoven, K. A review of the substrates used in microbial fuel cells (MFCs) for sustainable energy production. Bioresour. Technol. 2010, 101, 1533–1543. [Google Scholar] [CrossRef] [PubMed]
- El-Nahhal, Y.Z.; Al-Agha, M.R.; El-Nahhal, I.Y.; el Aila, N.A.; El-Nahal, F.I.; Alhalabi, R.A. Electricity generation from animal manure. Biomass Bioenergy 2020, 136, 105531. [Google Scholar] [CrossRef]
- Mohd Azmi, N.; Mohd Sabri, M.N.I.; Tajarudin, H.A.; Shoparwe, N.F.; Makhtar, M.M.Z.; Shukor, H.; Alam, M.; Siddiqui, M.R.; Rafatullah, M. The Effect of Different Pretreatment of Chicken Manure for Electricity Generation in Membrane-Less Microbial Fuel Cell. Catalysts 2022, 12, 810. [Google Scholar] [CrossRef]
- Oyiwona, G.E.; Ogbonna, J.C.; Anyanwu, C.U.; Okabe, S. Electricity generation potential of poultry droppings wastewater in microbial fuel cell using rice husk charcoal electrodes. Bioresour. Bioprocess. 2018, 5, 13. [Google Scholar] [CrossRef] [Green Version]
- Rojas-Flores, S.; de La Cruz-Noriega, M.; Nazario-Naveda, R.; Benites, S.M.; Delfín-Narciso, D.; Rojas-Villacorta, W.; Romero, C.V. Bioelectricity through microbial fuel cells using avocado waste. Energy Rep. 2022, 8, 376–382. [Google Scholar] [CrossRef]
- Singh, A.; Kaushik, A. Sustained energy production from wastewater in microbial fuel cell: Effect of inoculum sources, electrode spacing and working volume. 3 Biotech 2021, 11, 344. [Google Scholar] [CrossRef]
- Cheng, S.; Liu, H.; Logan, B.E. Increased Power Generation in a Continuous Flow MFC with Advective Flow through the Porous Anode and Reduced Electrode Spacing. Environ. Sci. Technol 2006, 40, 2426–2432. [Google Scholar] [CrossRef] [PubMed]
- Clauwaert, P.; Aelterman, P.; Pham, T.; De Schamphelaire, L.; Carballa, M.; Rabaey, K.; Verstraete, W. Minimizing losses in bio-electrochemical systems: The road to applications. Appl. Microbiol. Biotechnol. 2008, 79, 901–913. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Hour | mV pH | pH | ORP (mV) | OD (ppm) | Conductivity (µS/cm) | TDS (ppm) |
|---|---|---|---|---|---|---|
| 24 | 10 | 6.84 | −229.2 | 1.34 | 9044 | 4523 |
| 240 | 24.1 | 6.59 | −227.9 | 5.28 | 6480 | 3243 |
| CEPA | Simmons Citrate Agar | L.I.A. | SH2 | Motility | Ornithine | Methyl Red Indicator | Oxidase | Catalase | Gram Stain Test |
|---|---|---|---|---|---|---|---|---|---|
| Av_G1 | + | − | + | + | − | − | + | + | − |
| Hour | mV–pH | pH | ORP mV | OD ppm | uS/cm | TDS ppm |
|---|---|---|---|---|---|---|
| 24 | 10 | 6.84 | −229.2 | 1.34 | 9044 | 4523 |
| 42 | 6.7 | 6.89 | −211.8 | 2.03 | 6309 | 3156 |
| 74 | −3.2 | 7.06 | −182.2 | 1.94 | 7615 | 3806 |
| 95 | −1.7 | 7.04 | −176.5 | 3.57 | 8173 | 4088 |
| 119 | −10.1 | 7.18 | −163.5 | 2.76 | 17,230 | 8646 |
| 143 | 21.1 | 6.64 | −142.6 | 3.01 | 15,350 | 7680 |
| 164 | 22.4 | 6.63 | −179.6 | 2.44 | 12,470 | 6236 |
| 196 | 23.9 | 6.61 | −186.2 | 6.18 | 7149 | 6386 |
| 208 | 24.2 | 6.57 | −213.4 | 5.89 | 6917 | 4283 |
| 240 | 24.1 | 6.59 | −227.9 | 5.28 | 6480 | 3243 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jimenez Pacheco, H.G.; Portocarrero Banda, A.A.; Vilca Cayllahua, E.I.; Miranda Ramos, L.M.; Alvarez Tohalino, V.L.; Del Carpio Beltran, H.J.; Delgado Sarmiento, P.K. New Electrogenic Microorganism Citrobacter sp. Isolated from Microbial Fuel Cell and Bacterial Characteristics Determination. Energies 2023, 16, 3047. https://doi.org/10.3390/en16073047
Jimenez Pacheco HG, Portocarrero Banda AA, Vilca Cayllahua EI, Miranda Ramos LM, Alvarez Tohalino VL, Del Carpio Beltran HJ, Delgado Sarmiento PK. New Electrogenic Microorganism Citrobacter sp. Isolated from Microbial Fuel Cell and Bacterial Characteristics Determination. Energies. 2023; 16(7):3047. https://doi.org/10.3390/en16073047
Chicago/Turabian StyleJimenez Pacheco, Hugo Guillermo, Abdel Alejandro Portocarrero Banda, Eric Ivan Vilca Cayllahua, Lilia Mary Miranda Ramos, Victor Ludgardo Alvarez Tohalino, Herbert Jesús Del Carpio Beltran, and Pavel Kewin Delgado Sarmiento. 2023. "New Electrogenic Microorganism Citrobacter sp. Isolated from Microbial Fuel Cell and Bacterial Characteristics Determination" Energies 16, no. 7: 3047. https://doi.org/10.3390/en16073047
APA StyleJimenez Pacheco, H. G., Portocarrero Banda, A. A., Vilca Cayllahua, E. I., Miranda Ramos, L. M., Alvarez Tohalino, V. L., Del Carpio Beltran, H. J., & Delgado Sarmiento, P. K. (2023). New Electrogenic Microorganism Citrobacter sp. Isolated from Microbial Fuel Cell and Bacterial Characteristics Determination. Energies, 16(7), 3047. https://doi.org/10.3390/en16073047