Microalgae Harvest through Fungal Pelletization—Co-Culture of Chlorella vulgaris and Aspergillus niger

Abstract
:1. Introduction
2. Results and Discussion
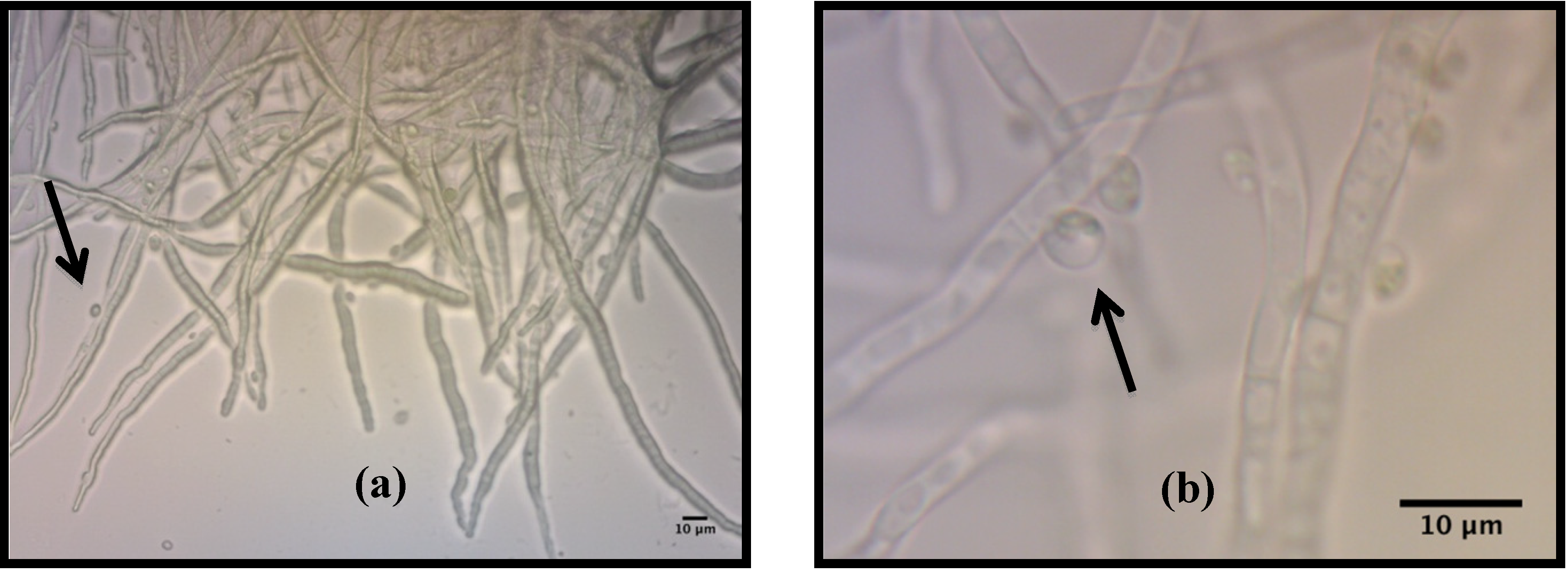
2.1. Description of Pellets Formed During the Co-Culture of C. vulgaris and A. niger

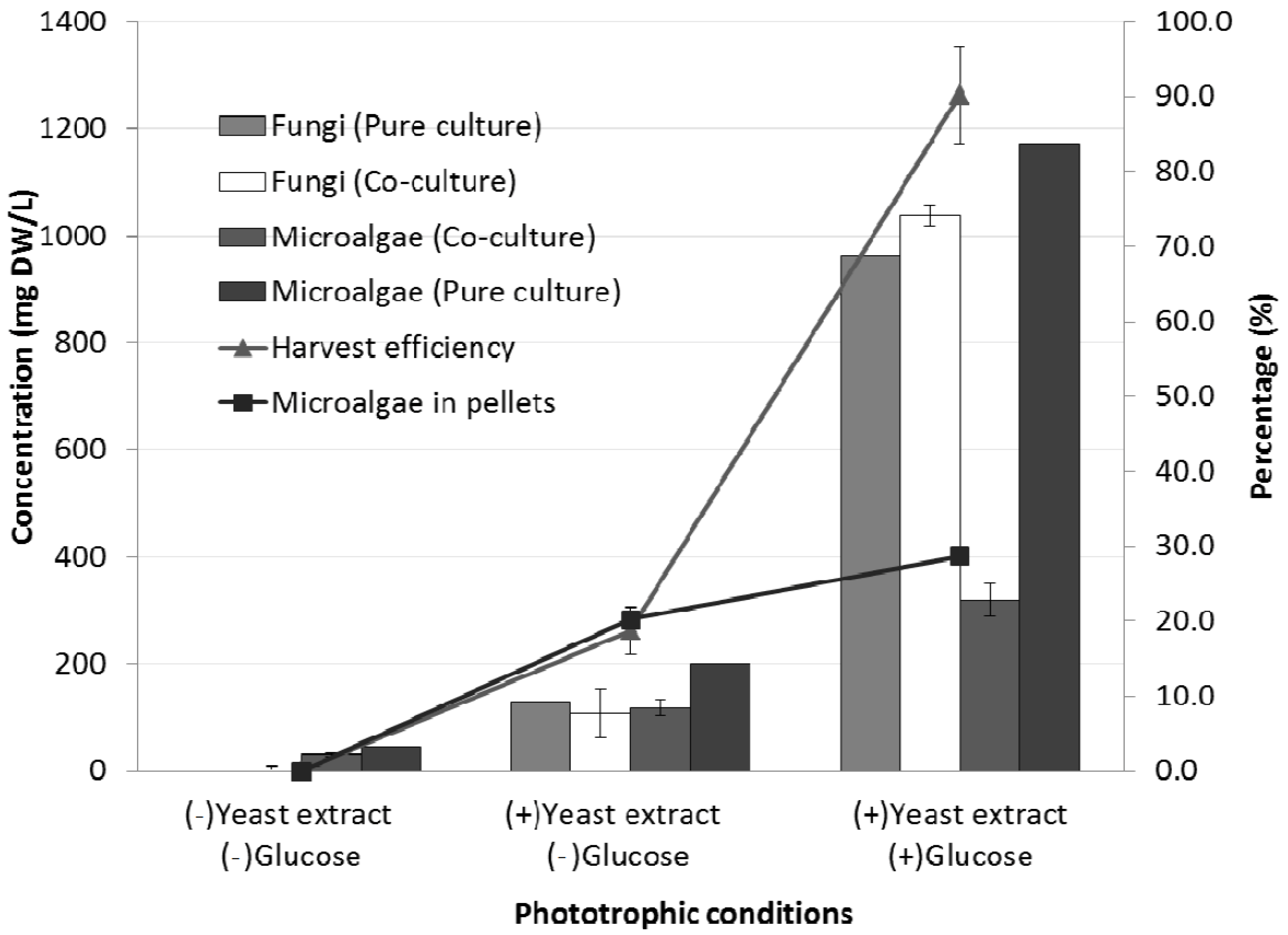
2.2. Effects of Different Carbon Sources on the Co-Culture Process
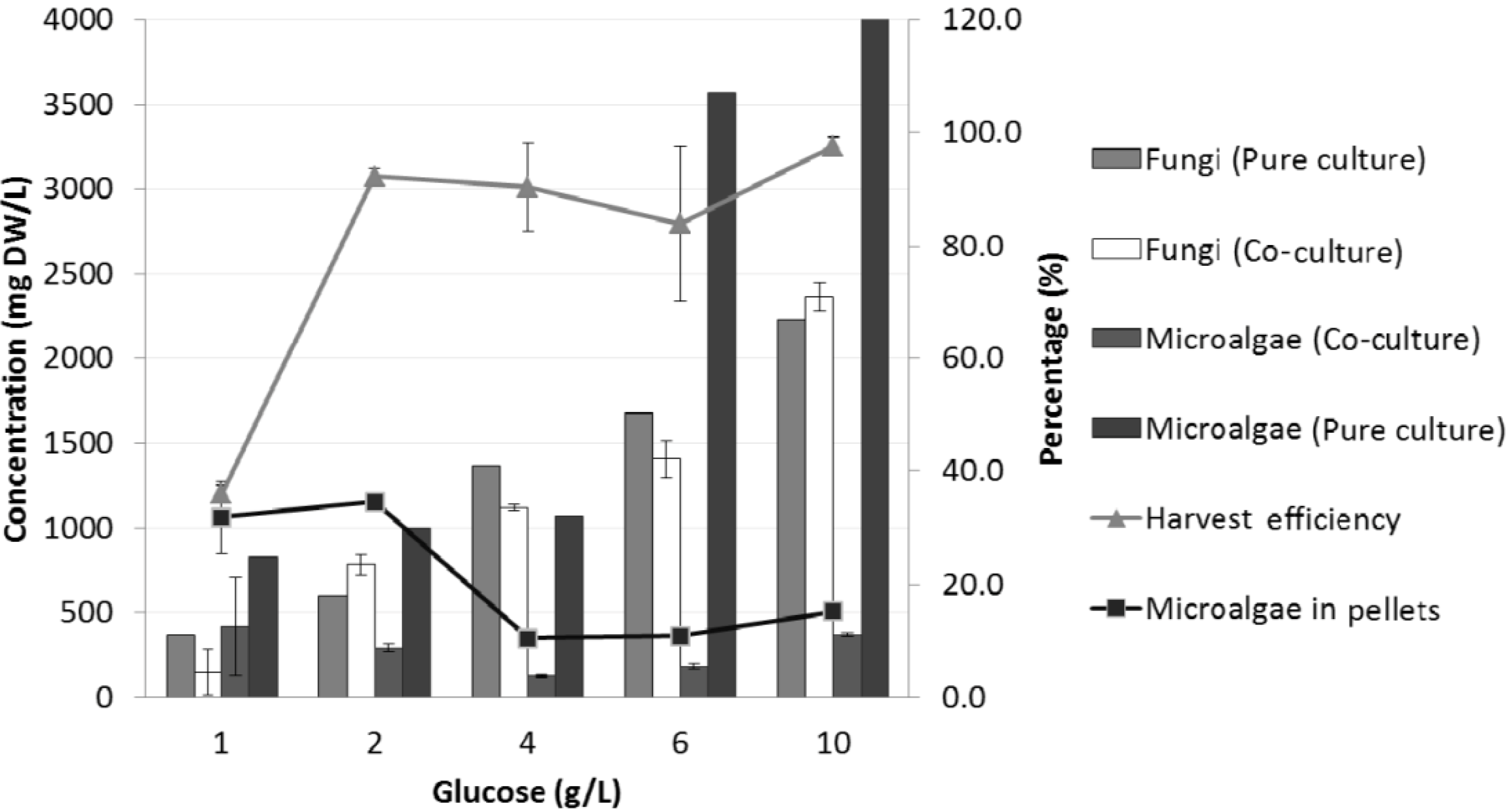
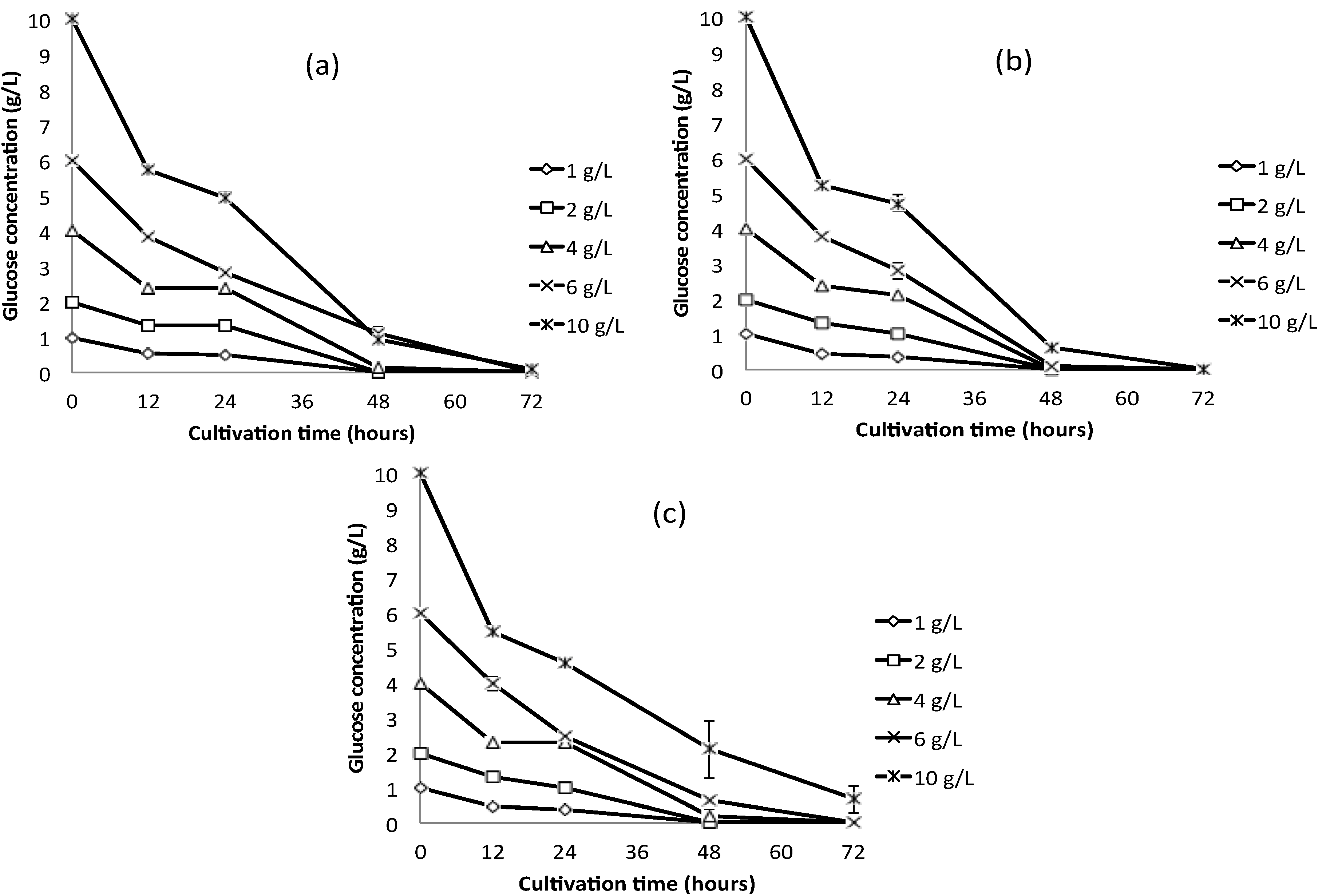
2.2.1. Glucose


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Co-culture | Glucose concentration (g/L) | ||||
|---|---|---|---|---|---|
| 1 | 2 | 4 | 6 | 10 | |
| # of pellets/mL | 2 ± 0.2 | 5 ± 1.5 | 6 ± 0.7 | 7 ± 0.7 | 7 ± 0.8 |
| Average diameter (mm) | 3 ± 0.1 | 3.3 ± 0.3 | 3.4 ± 0.2 | 3.6 ± 0.8 | 3.8 ± 0.4 |
| Average final pH | 7.1 ± 0.0 | 7.1 ± 0.2 | 5.6 ± 0.9 | 4.1 ± 0.1 | 4.0 ± 0.2 |

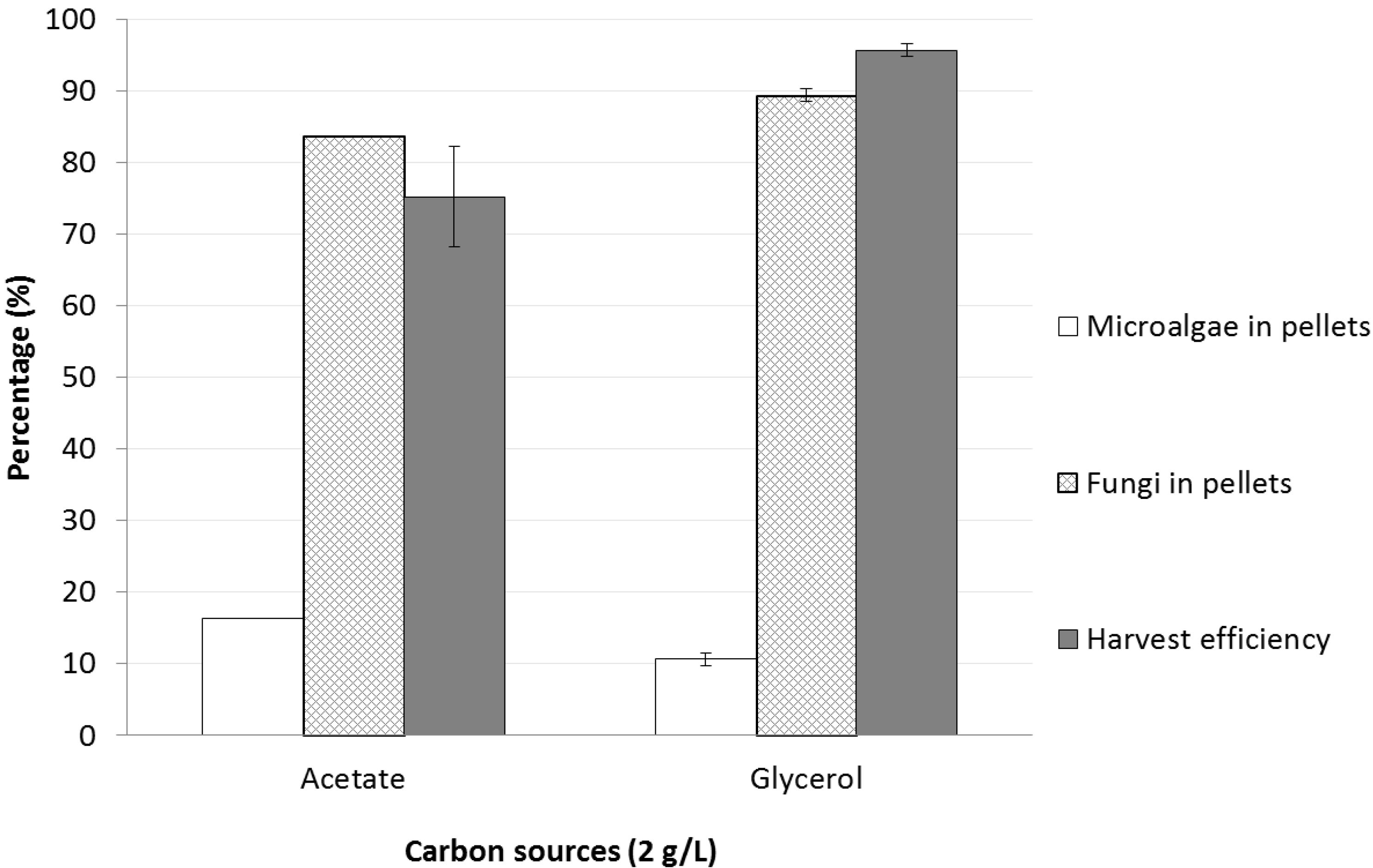
2.2.2. Sodium Acetate or Glycerol

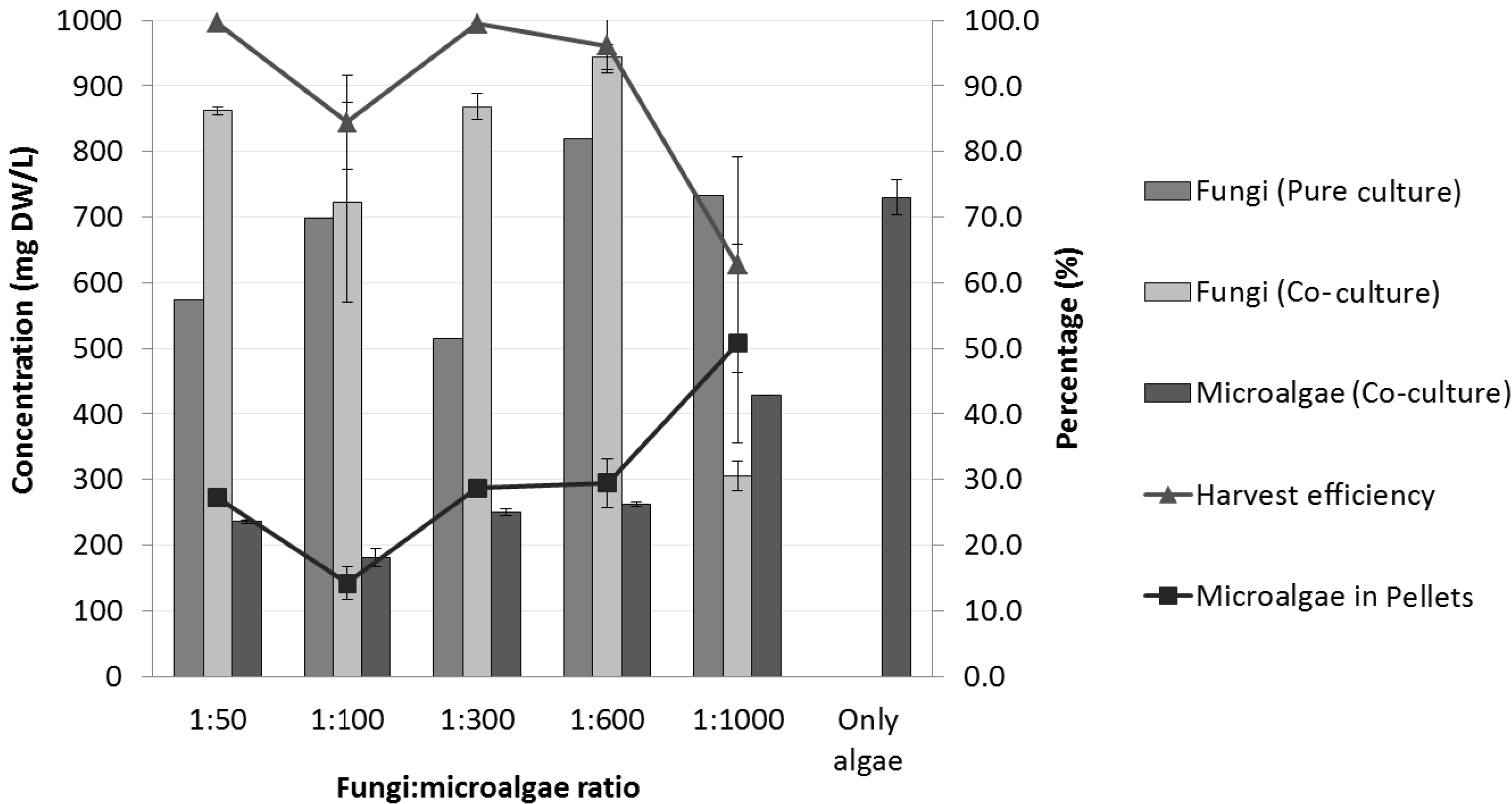
2.3. Effects of Initial Fungal Spore Concentration on the Co-Culture
| Co-culture | Fungi:microalgae ratio | ||||
|---|---|---|---|---|---|
| 1:50 | 1:100 | 1:300 | 1:600 | 1:1,000 | |
| # of pellets/mL | 6 ± 0.6 | 5 ± 2.8 | 5 ± 0.2 | 4 ± 0.4 | 4 ± 0.8 |
| Average diameter (mm) | 2.6 ± 0.4 | 2.9 ± 0.4 | 3.4 ± 0.2 | 3.6 ± 0.9 | 3.6 ± 0.6 |

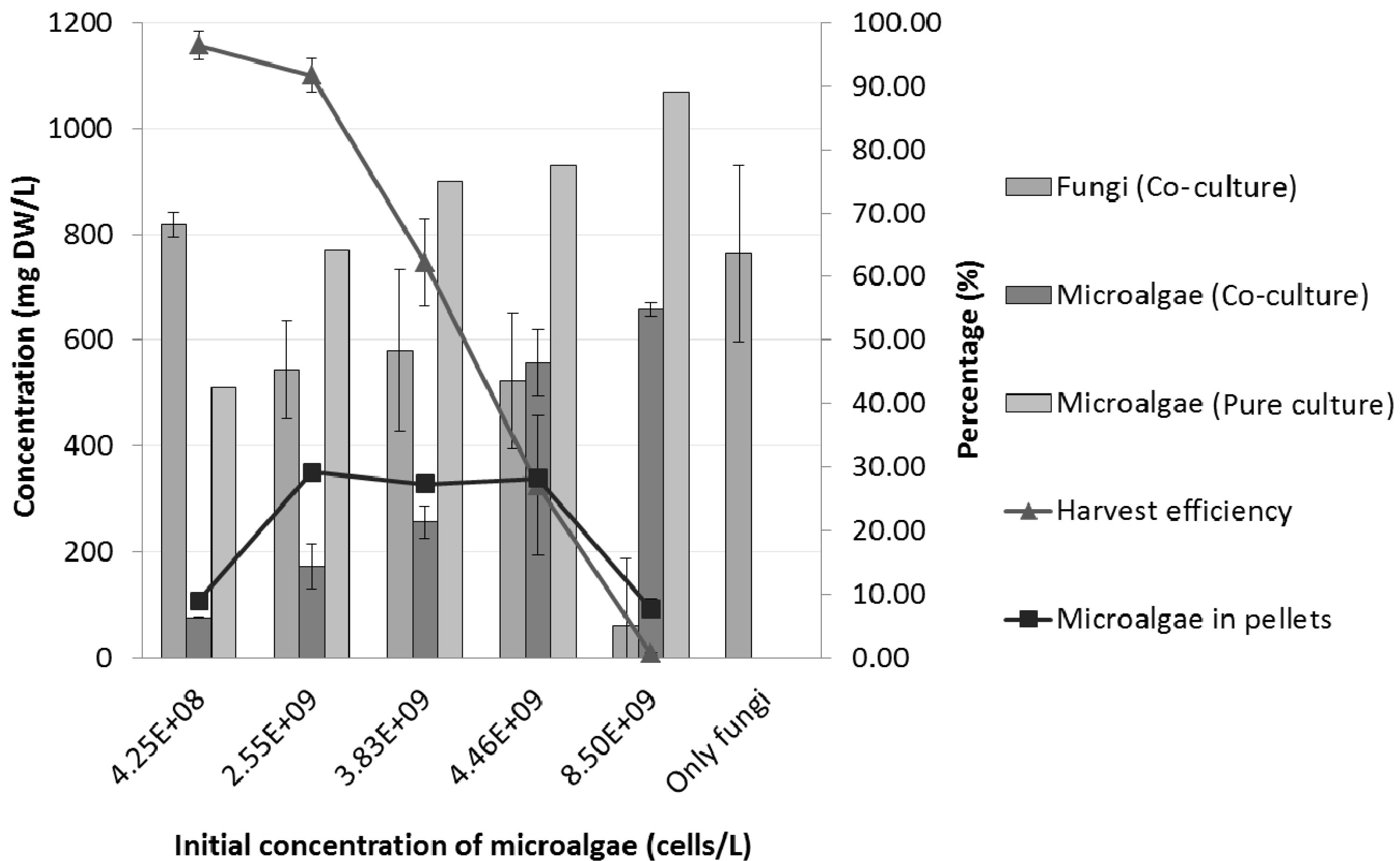
2.4. Effects of Initial Microalgal Cell Concentration on the Co-Culture Process
| Co-culture | Initial concentration of microalgal cells (cells/L) | |||||
|---|---|---|---|---|---|---|
| 0 | 4.25 × 108 | 2.55 × 109 | 3.83 × 109 | 4.46 × 109 | 8.50 × 109 | |
| # of pellets/mL | 5 ± 1.2 | 5 ± 1.1 | 4 ± 0.5 | 4 ± 0.4 | 3 ± 0.4 | 1 ± 0.4 |
| Average diameter (mm) | 3.5 ± 2.1 | 3.4 ± 0.1 | 3.3 ± 0.1 | 3.3 ± 0.3 | 3.0 ± 0.8 | 2.5 ± 0.1 |

2.5. Effects of Light on the Co-Culture Process

3. Experimental Section
3.1. Cell Cultivation of Microalgae and Fungal Strains
3.2. Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Yang, C.; Hua, Q.; Shimizu, K. Energetics and carbon metabolism during growth of microalgal cells under photoautotrophic, mixotrophic and cyclic light-autotrophic/dark-heterotrophic conditions. Biochem. Eng. J. 2000, 6, 87–102. [Google Scholar] [CrossRef]
- Miao, X.; Wu, Q. Biodiesel production from heterotrophic microalgal oil. Bioresour. Technol. 2006, 97, 841–846. [Google Scholar] [CrossRef]
- Shelef, G.; Sukenik, A.; Green, M. Microalgae Harvesting and Processing: A Literature Review; Technical Report for Technion Research and Development Foundation Ltd.: Haifa, Israel, August 1984. [Google Scholar]
- Zhang, J.G.; Hu, B. A novel method to harvest microalgae via co-culture of filamentous fungi to form cell pellets. Bioresour. Technol. 2012, 114, 529–535. [Google Scholar] [CrossRef]
- Zhou, W.; Cheng, Y.; Li, Y.; Wan, Y.; Liu, Y.; Lin, X.; Ruan, R. Novel fungal pelletization-assisted technology for algae harvesting and wastewater treatment. Appl. Biochem. Biotechnol. 2012, 167, 214–228. [Google Scholar] [CrossRef]
- Xia, C.; Zhang, J.; Zhang, W.; Hu, B. A new cultivation method for microbial oil production: Cell pelletization and lipid accumulation by Mucor circinelloides. Biotechnol. Biofuels 2011, 4, 15. [Google Scholar] [CrossRef]
- Zhou, W.G.; Min, M.; Hu, B.; Ma, X.C.; Liu, Y.H.; Wang, Q.; Shi, J.; Chen, P.; Ruan, R. Filamentous fungi assisted bio-flocculation: A novel alternative technique for harvesting heterotrophic and autotrophic microalgal cells. Sep. Purif. Technol. 2013, 107, 158–165. [Google Scholar] [CrossRef]
- Mackay, S.; Pereira Gomes, E.; Rossi, P.; Schwitzguebel, J.P.; Holliger, C. Pelletization of micro-algae by induced lichen formation through co-culture with filamentous fungi. In Proceedings of 5th Swiss Microbial Ecology Meeting, Murten, Switzerland, 4–6 February 2013.
- Lustigman, B.; Lee, L.H.; Khalil, A. Effects of nickel and pH on the growth of Chlorella vulgaris. Bull. Environ. Contam. Toxicol. 1995, 55, 73–80. [Google Scholar] [CrossRef]
- Nielsen, J. Modelling the growth of filamentous fungi. In Modern Biochemical Engineering; Springer: Berlin/Heidelberg, Germany, 1992; Volume 46; pp. 187–223. [Google Scholar]
- Liang, Y.N.; Sarkany, N.; Cui, Y. Biomass and lipid productivities of Chlorella vulgaris under autotrophic, heterotrophic and mixotrophic growth conditions. Biotechnol. Lett. 2009, 31, 1043–1049. [Google Scholar] [CrossRef]
- Metz, B.; Kossen, N.W.F. The growth of molds in the form of pellets—A literature review. Biotechnol. Bioeng. 1977, 19, 781–799. [Google Scholar] [CrossRef]
- Zmak, P.M.; Podgornik, A.; Podgornik, H.; Koloini, T. Impact of pellet size on growth and lignin peroxidase activity of Phanerochaete chrysosporium. World J. Microbiol. Biotechnol. 2006, 22, 1243–1249. [Google Scholar]
- Heredia-Arroyo, T.; Wei, W.; Hu, B. Oil Accumulation via Heterotrophic/Mixotrophic Chlorella protothecoides. Appl. Biochem. Biotechnol. 2010, 162, 1978–1995. [Google Scholar] [CrossRef]
- Heredia-Arroyo, T.; Wei, W.; Ruan, R.; Hu, B. Mixotrophic cultivation of Chlorella vulgaris and its potential application for the oil accumulation from non-sugar materials. Biomass Bioenergy 2011, 35, 2245–2253. [Google Scholar] [CrossRef]
- Becker, E.W. Microalgae: Biotechnology and Microbiology; Canbridge University Press: Canbridge, UK, 1995. [Google Scholar]
- Xiong, W.; Li, X.F.; Xiang, J.Y.; Wu, Q.Y. High-density fermentation of microalga Chlorella protothecoides in bioreactor for microbio-diesel production. Appli. Microbiol. Biotechnol. 2008, 78, 29–36. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Gultom, S.O.; Zamalloa, C.; Hu, B. Microalgae Harvest through Fungal Pelletization—Co-Culture of Chlorella vulgaris and Aspergillus niger. Energies 2014, 7, 4417-4429. https://doi.org/10.3390/en7074417
Gultom SO, Zamalloa C, Hu B. Microalgae Harvest through Fungal Pelletization—Co-Culture of Chlorella vulgaris and Aspergillus niger. Energies. 2014; 7(7):4417-4429. https://doi.org/10.3390/en7074417
Chicago/Turabian StyleGultom, Sarman Oktovianus, Carlos Zamalloa, and Bo Hu. 2014. "Microalgae Harvest through Fungal Pelletization—Co-Culture of Chlorella vulgaris and Aspergillus niger" Energies 7, no. 7: 4417-4429. https://doi.org/10.3390/en7074417
APA StyleGultom, S. O., Zamalloa, C., & Hu, B. (2014). Microalgae Harvest through Fungal Pelletization—Co-Culture of Chlorella vulgaris and Aspergillus niger. Energies, 7(7), 4417-4429. https://doi.org/10.3390/en7074417