A Probabilistic Model for Crystal Growth Applied to Protein Deposition at the Microscale



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. The Model
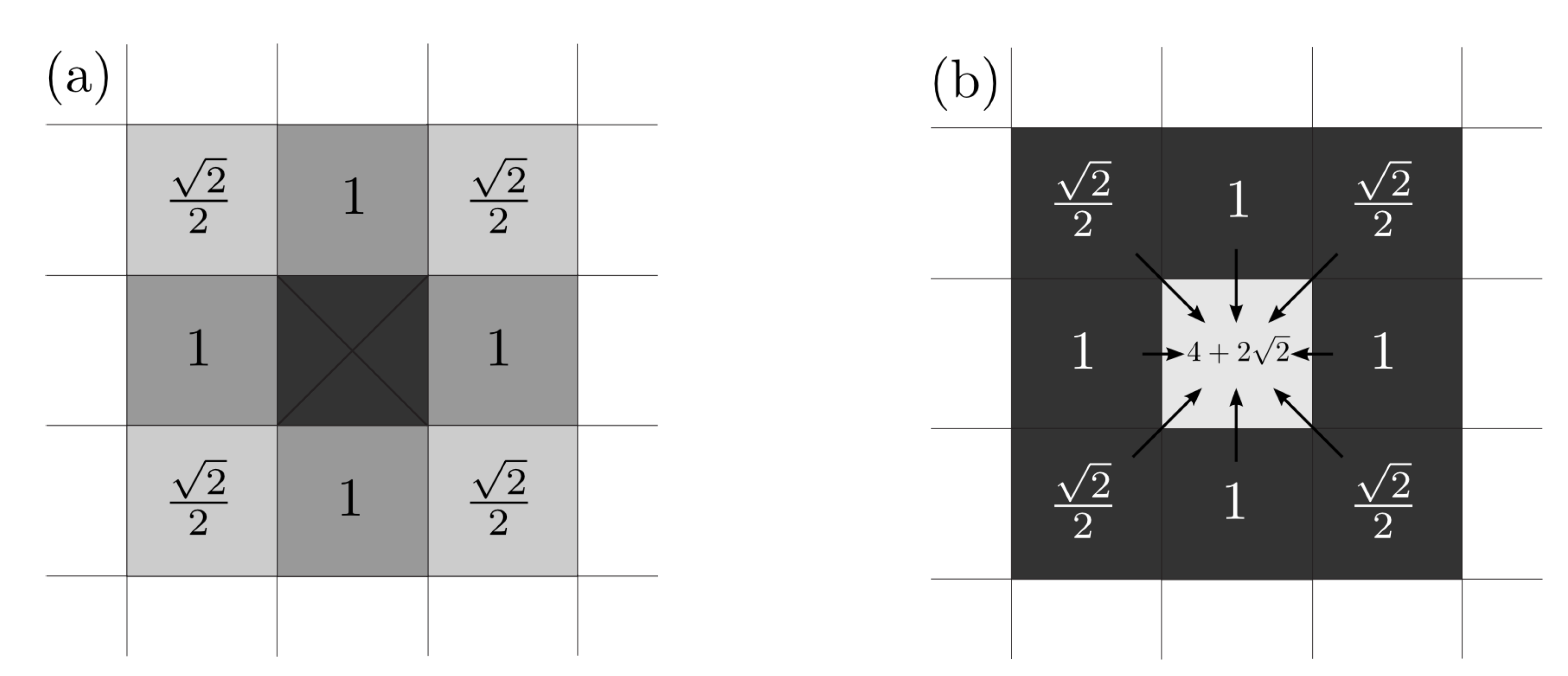
2.1. Structure
- If a cell is occupied, it cannot be occupied again, i.e., if , then .
- If a cell is occupied, then the probability of occupation of the adjacent free cells is increased, see Equation (1).
2.2. Kinetics
3. The Parameters
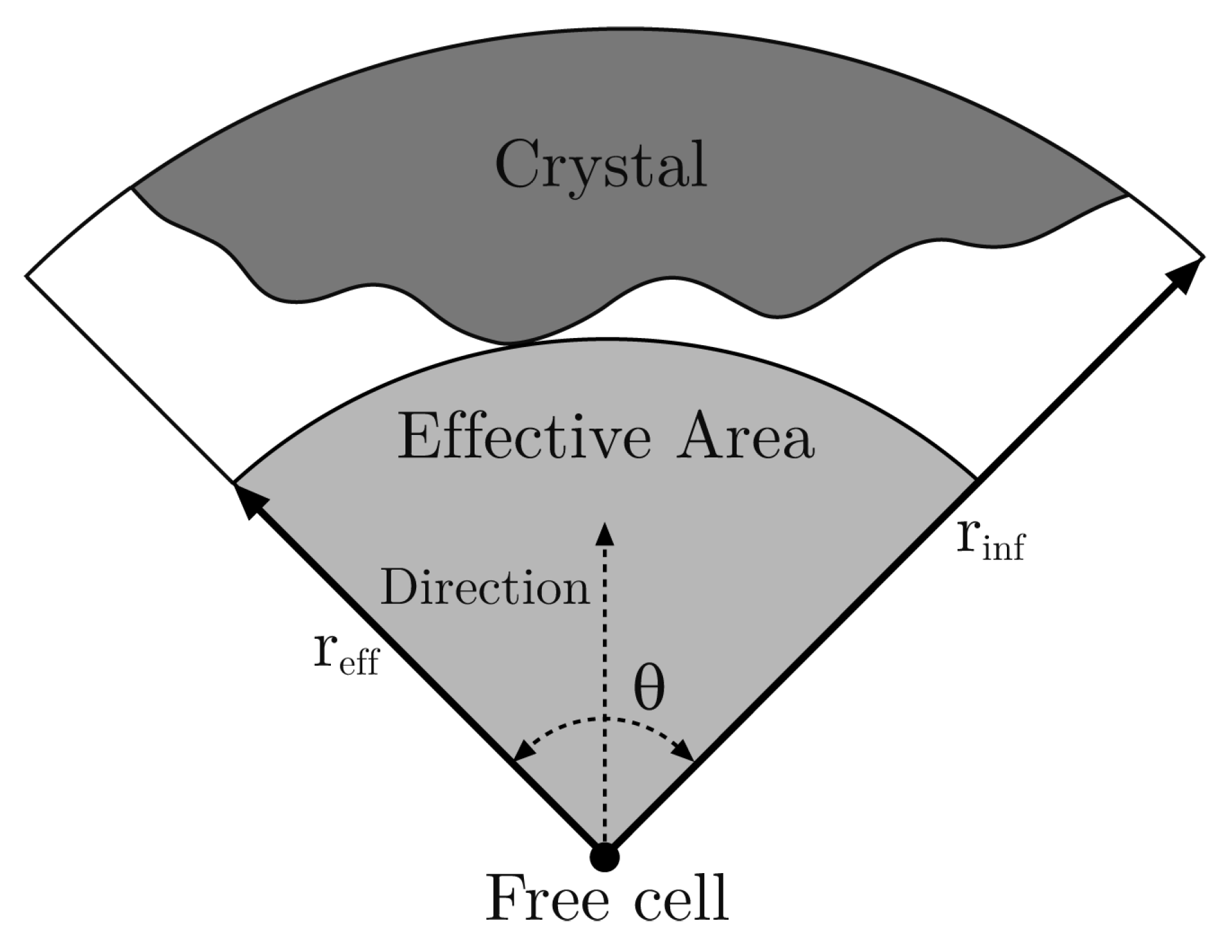
3.1. Structural Parameters
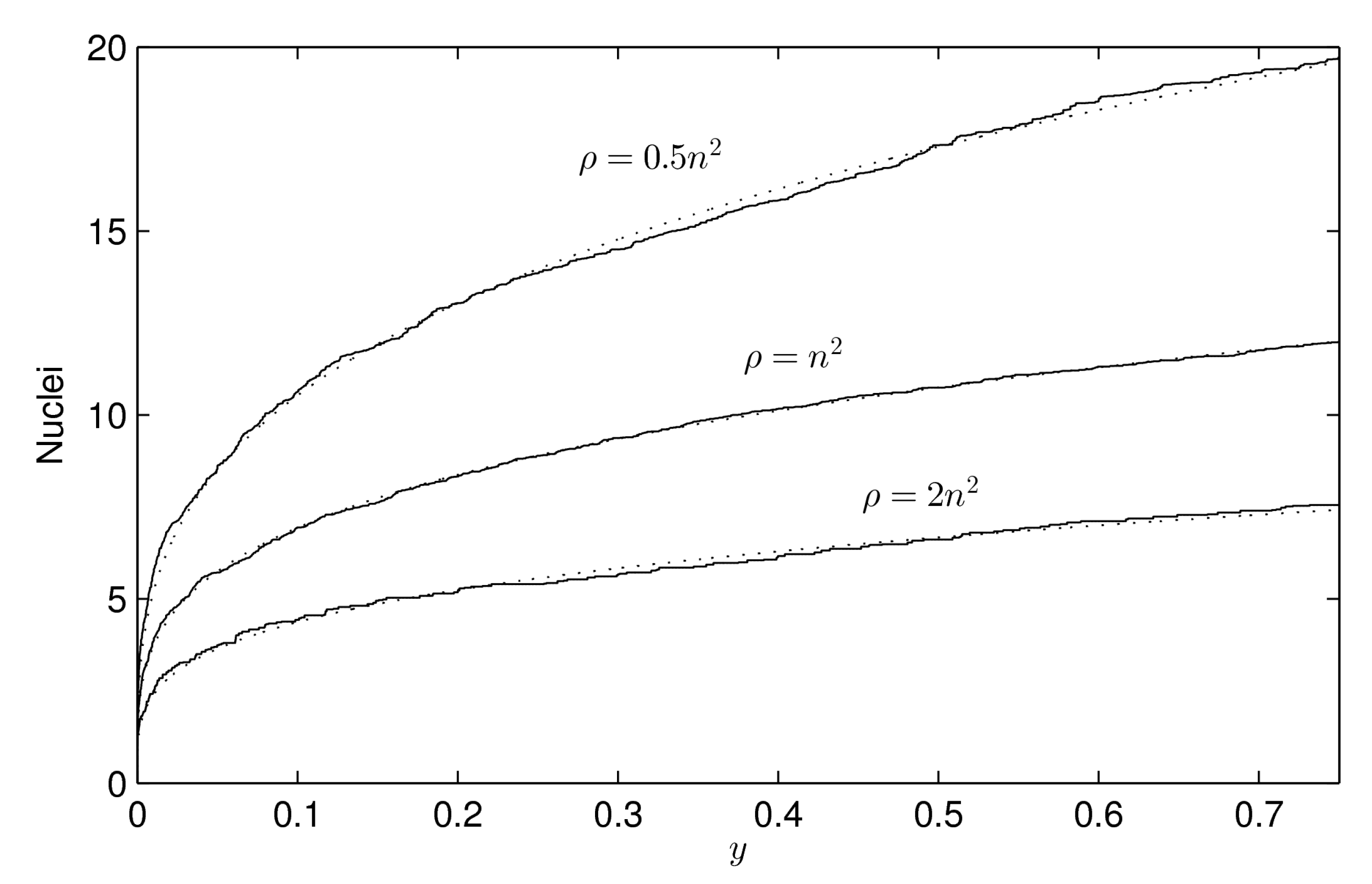
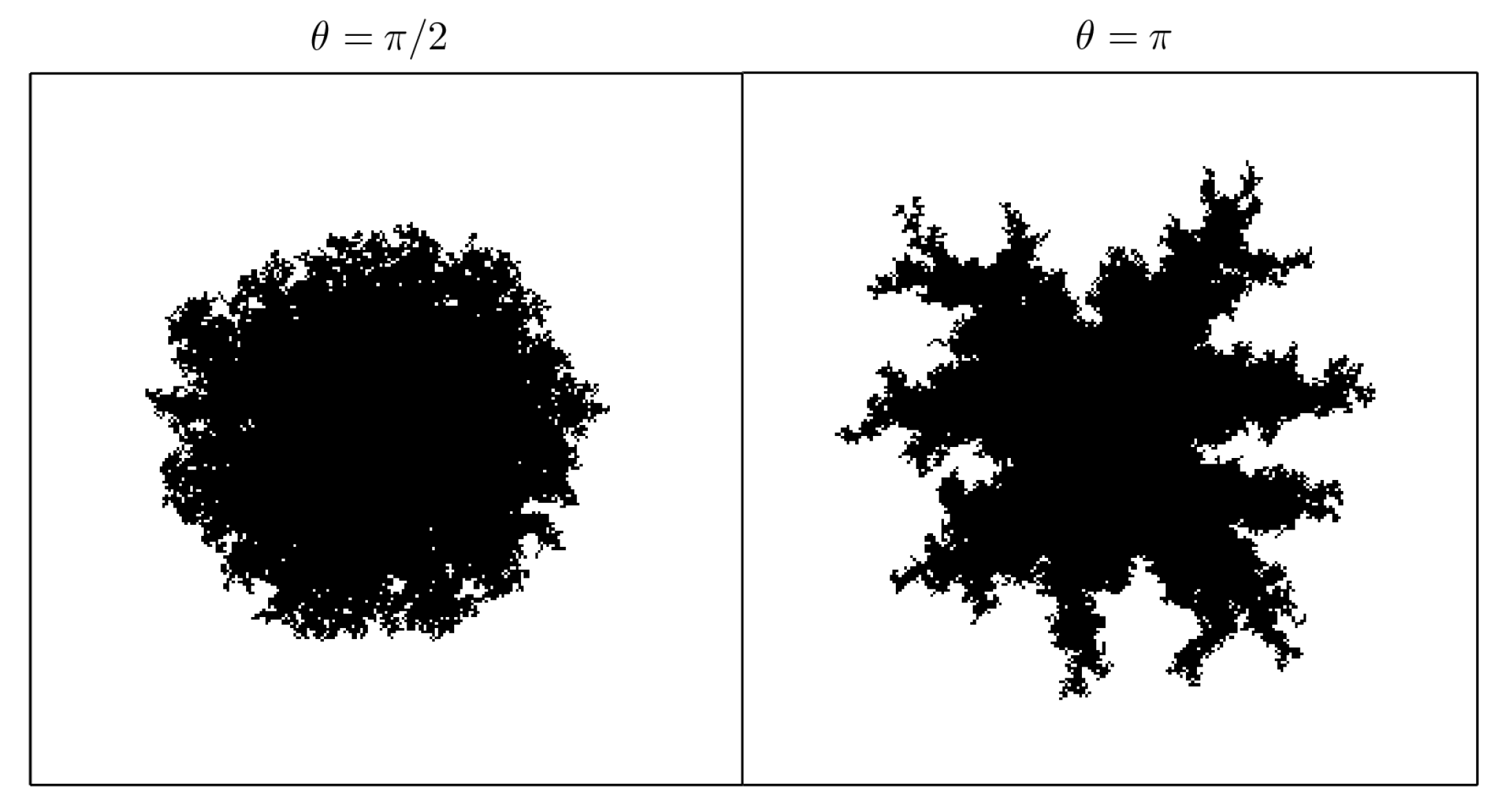
- : the “difficulty of filling” parameter, see Equation (2). It ranges from 0 to 1 and determines the importance of the maximum effective radius in the model. It is also related to the rate at which void regions are filled. Setting means that the probability of occupation of a cell does not depend on the maximum effective radius. In Figure 6 how this parameter (along with ) affects the shape of the crystal is shown.
- : the “effective dimension” parameter, see Equation (2). It must be positive and, as explained before, in our model, is set to 2. It also determines how the maximum effective radius affects the crystal growth.
3.2. Kinetic Parameters
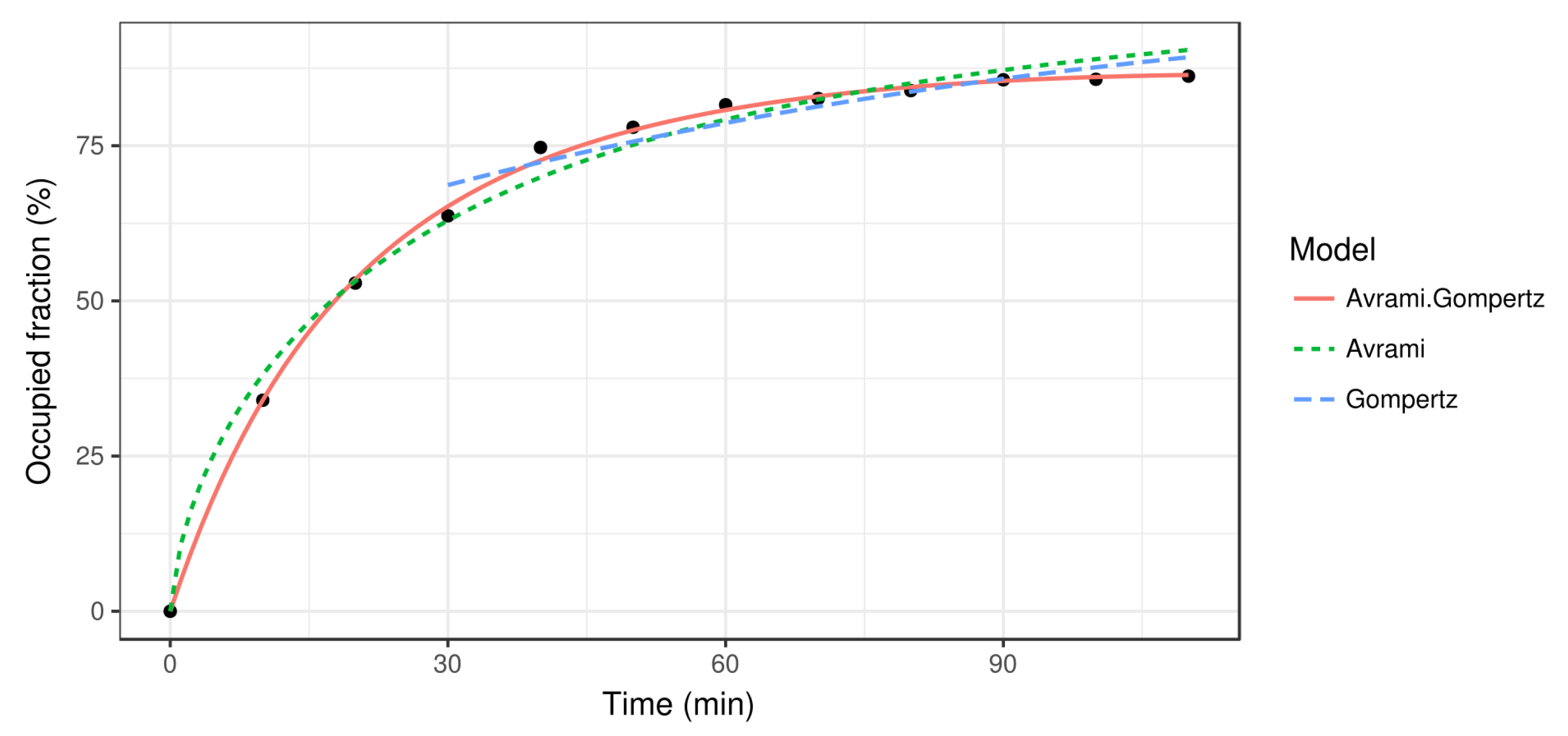
- : parameters of the Gompertz function in Equation (6) that models the last stages of the process.
4. Results
4.1. Protein Recrystallization
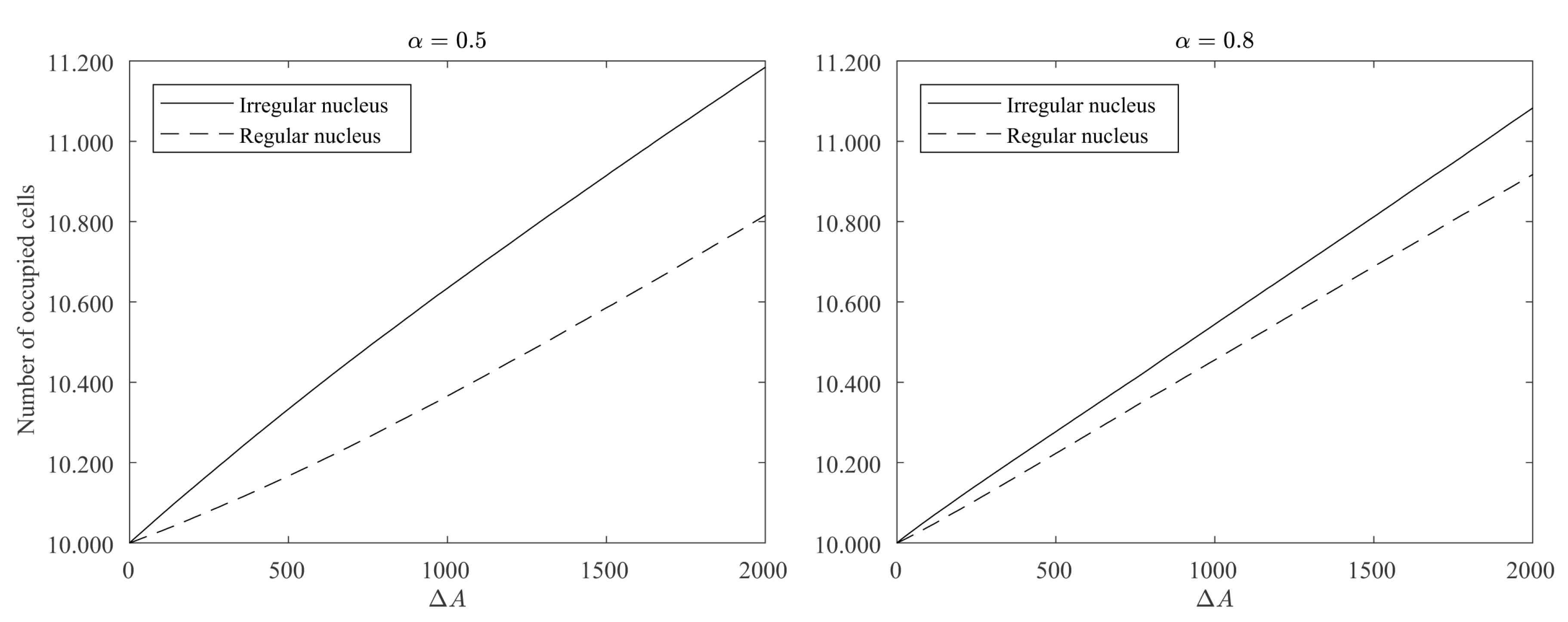
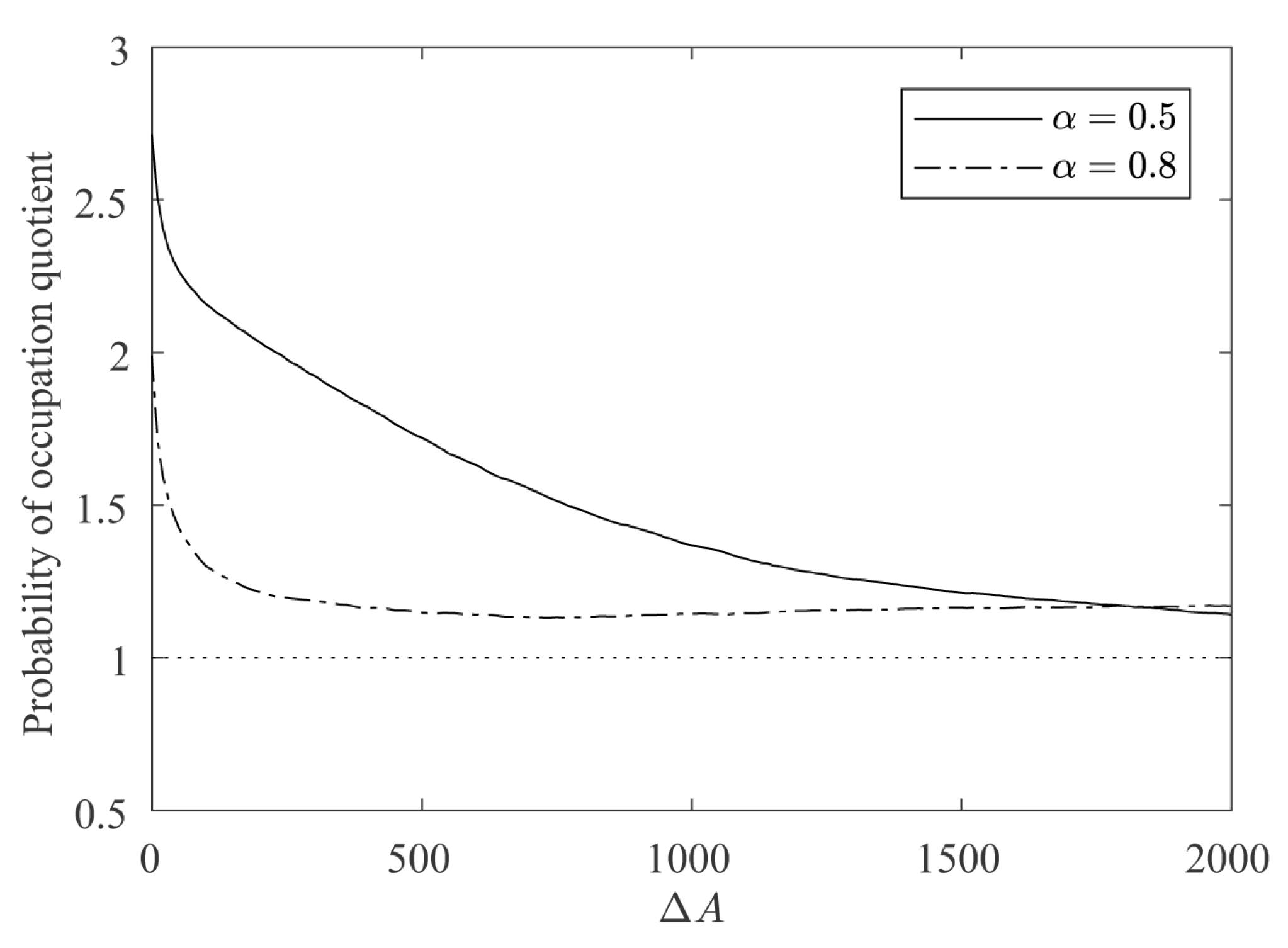
4.2. Influence of the Contour Regularity
5. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kittel, C. Introduction to Solid State Physics, 7th ed.; Wiley India Pvt. Limited: Noida, India, 2007. [Google Scholar]
- Veis, A.; Dorvee, J.R. Biomineralization Mechanisms: A New Paradigm for Crystal Nucleation in Organic Matrices. Calcif. Tissue Int. 2013, 93, 307–315. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.; Ruan, S.; Cai, T.; Yu, L. Fast Surface Diffusion and Crystallization of Amorphous Griseofulvin. J. Phys. Chem. B 2017, 121, 9463–9468. [Google Scholar] [CrossRef] [PubMed]
- Semenova, N.M.; Dick, A.A.; Skokov, V.N.; Koverda, V.P. Fractal structurization under crystallization of amorphous germanium in three-layer films Ag-Ge-Ag. Phys. Status Solidi (A) 1993, 139, 287–293. [Google Scholar] [CrossRef]
- Himawan, C.; Starov, V.; Stapley, A. Thermodynamic and kinetic aspects of fat crystallization. Adv. Colloid Interface Sci. 2006, 122, 3–33. [Google Scholar] [CrossRef]
- Zhou, D.; Zhang, G.G.Z.; Law, D.; Grant, D.J.W.; Schmitt, E.A. Thermodynamics, Molecular Mobility and Crystallization Kinetics of Amorphous Griseofulvin. Mol. Pharm. 2008, 5, 927–936. [Google Scholar] [CrossRef] [PubMed]
- Avrami, M. Kinetics of Phase Change. I General Theory. J. Chem. Phys. 1939, 7, 1103–1112. [Google Scholar] [CrossRef]
- Avrami, M. Kinetics of Phase Change. II Transformation-Time Relations for Random Distribution of Nuclei. J. Chem. Phys. 1940, 8, 212–224. [Google Scholar] [CrossRef]
- Laird, A.K. Dynamics of tumor growth. Br. J. Cancer 1964, 18, 490–502. [Google Scholar] [CrossRef]
- Chernov, A.A. Protein crystals and their growth. J. Struct. Biol. 2003, 142, 3–21. [Google Scholar] [CrossRef]
- Siódmiak, J.; Gadomski, A. Growing lysozyme crystals under various physicochemical conditions: Computer modelling. J. Non-Cryst. Solids 2008, 354. [Google Scholar] [CrossRef]
- Durbin, S.; Feher, G. Simulation of lysozyme crystal growth by the Monte Carlo method. J. Cryst. Growth 1991, 110. [Google Scholar] [CrossRef]
- Sleytr, U.B.; Sára, M.; Pum, D.; Schuster, B. Characterization and use of crystalline bacterial cell surface layers. Prog. Surf. Sci. 2001, 68, 231–278. [Google Scholar] [CrossRef]
- López, A.E.; Moreno-Flores, S.; Pum, D.; Sleytr, U.B.; Toca-Herrera, J.L. Surface Dependence of Protein Nanocrystal Formation. Small 2010, 6. [Google Scholar] [CrossRef] [PubMed]
- Chung, S.; Shin, S.H.; Bertozzi, C.R.; De Yoreo, J.J. Self-catalyzed growth of S layers via an amorphous-to-crystalline transition limited by folding kinetics. Proc. Natl. Acad. Sci. USA 2010, 107, 16536–16541. [Google Scholar] [CrossRef] [PubMed]
- Lejardi, A.; López, A.E.; Sarasua, J.R.; Sleytr, U.B.; Toca-Herrera, J.L. Making novel bio-interfaces through bacterial protein recrystallization on biocompatible polylactide derivative films. J. Chem. Phys. 2013, 139, 121903. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2017. [Google Scholar]
- Elzhov, T.V.; Mullen, K.M.; Spiess, A.N.; Bolker, B. minpack.lm: R Interface to the Levenberg-Marquardt Nonlinear Least-Squares Algorithm Found in MINPACK, Plus Support for Bounds; R Package Version 1.2-1; 2016; Available online: https://rdrr.io/cran/minpack.lm/ (accessed on 4 February 2019).
- Barabási, A.L.; Stanley, H.E. Fractal Concepts in Surface Growth; Cambridge University Press: Cambridge, UK, 1995. [Google Scholar]
- Stel, B.; Cometto, F.; Rad, B.; De Yoreo, J.; Lingenfelder, M. Dynamically resolved self-assembly of S-layer proteins on solid surfaces. Chem. Commun. 2018, 54, 10264–10267. [Google Scholar] [CrossRef] [PubMed]












© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bolos, V.J.; Benitez, R.; Eleta-Lopez, A.; Toca-Herrera, J.L. A Probabilistic Model for Crystal Growth Applied to Protein Deposition at the Microscale. Materials 2019, 12, 479. https://doi.org/10.3390/ma12030479
Bolos VJ, Benitez R, Eleta-Lopez A, Toca-Herrera JL. A Probabilistic Model for Crystal Growth Applied to Protein Deposition at the Microscale. Materials. 2019; 12(3):479. https://doi.org/10.3390/ma12030479
Chicago/Turabian StyleBolos, Vicente J., Rafael Benitez, Aitziber Eleta-Lopez, and Jose L. Toca-Herrera. 2019. "A Probabilistic Model for Crystal Growth Applied to Protein Deposition at the Microscale" Materials 12, no. 3: 479. https://doi.org/10.3390/ma12030479
APA StyleBolos, V. J., Benitez, R., Eleta-Lopez, A., & Toca-Herrera, J. L. (2019). A Probabilistic Model for Crystal Growth Applied to Protein Deposition at the Microscale. Materials, 12(3), 479. https://doi.org/10.3390/ma12030479