Conductance Changes in Bovine Serum Albumin Caused by Drug-Binding Triggered Structural Transitions


Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. CD Spectroscopy and Estimation of Secondary Structural Contents
2.3. BSA and Drug-BSA Monolayer Preparation and CAFM Measurement
3. Results and Discussion
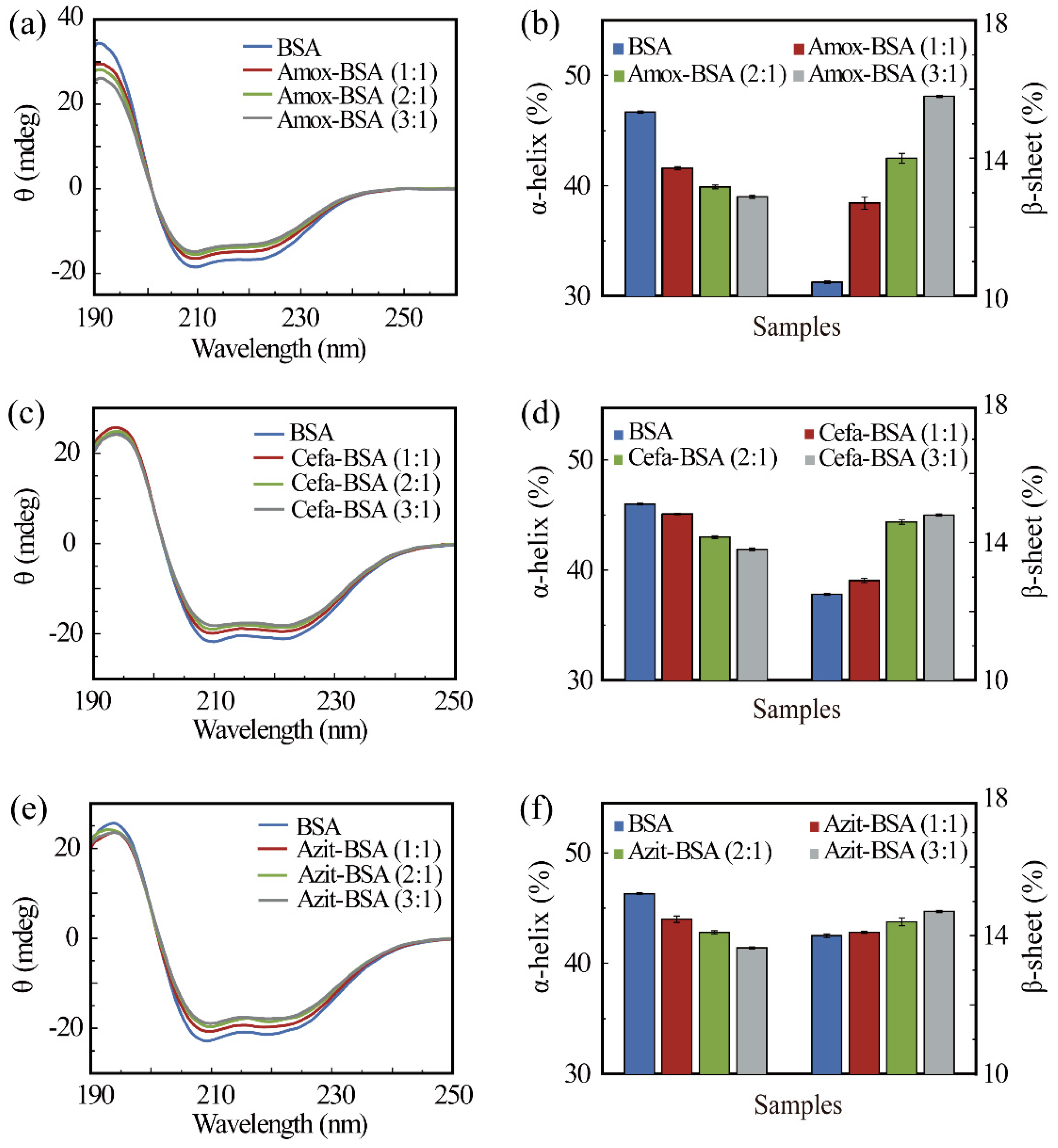
3.1. Drug-Binding Triggered Secondary Structure Transitions in BSA
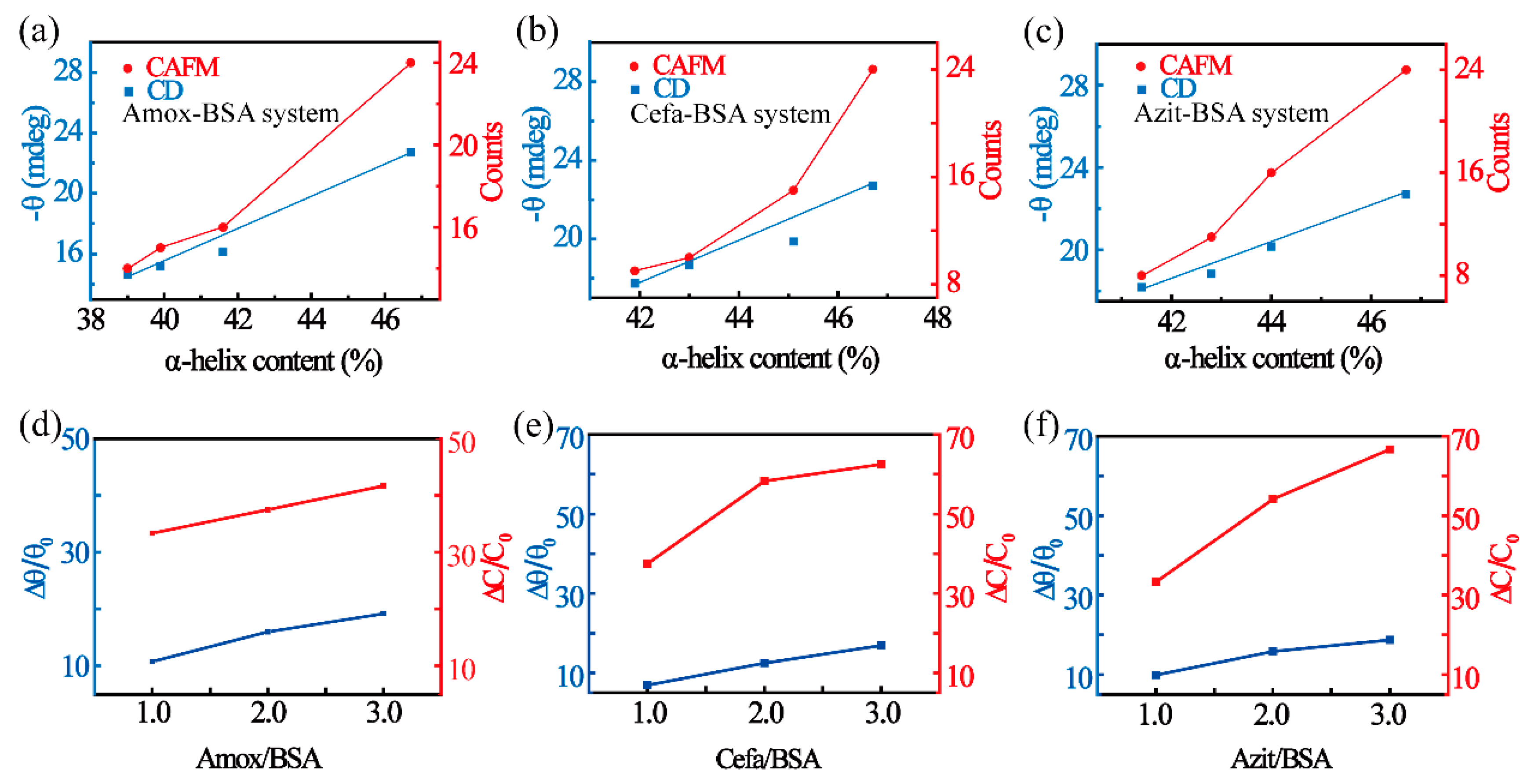
3.2. Change in the Conductance of BSA Due to Secondary Structure Transition
3.3. The Correlation Between BSA Conductance and Secondary Structural Content
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Aviram, A.; Ratner, M.A. Molecular rectifiers. Chem. Phys. Lett. 1974, 29, 277–283. [Google Scholar] [CrossRef]
- Davis, J.J.; Morgan, D.A.; Wrathmell, C.L.; Axford, D.N.; Zhao, J.; Wang, N. Molecular bioelectronics. J. Mater. Chem. 2005, 15, 2160–2174. [Google Scholar] [CrossRef]
- Guiseppi-Elie, A.; Brahim, S.; Slaughter, G.; Ward, K.R. Design of a subcutaneous implantable biochip for monitoring of glucose and lactate. IEEE Sens. J. 2005, 5, 345–355. [Google Scholar] [CrossRef]
- Farahi, R.H.; Ferrell, T.L.; Guiseppi-Elie, A.; Hansen, P. Integrated electronics platforms for wireless implantable biosensors. In Proceedings of the 2007 IEEE/NIH Life Science Systems and Applications Workshop, Bethesda, MD, USA, 8–9 November 2007; pp. 27–30. [Google Scholar]
- Carrara, S.; Cavallini, A.; Garg, A.; De Micheli, G. Dynamical spot queries to improve specificity in P450s based multi-drugs monitoring. In Proceedings of the 2009 ICME International Conference on Complex Medical Engineering, Tempe, AZ, USA, 9–11 April 2009; pp. 1–6. [Google Scholar]
- Bostick, C.D.; Mukhopadhyay, S.; Pecht, I.; Sheves, M.; Cahen, D.; Lederman, D. Protein bioelectronics: A review of what we do and do not know. Rep. Prog. Phys. 2018, 81, 026601. [Google Scholar] [CrossRef] [PubMed]
- Ligler, F.S.; Taitt, C.R.; Shriver-Lake, L.C.; Sapsford, K.E.; Shubin, Y.; Golden, J.P. Array biosensor for detection of toxins. Anal. Bioanal. Chem. 2003, 377, 469–477. [Google Scholar] [CrossRef]
- Carrara, S.; Shumyantseva, V.V.; Archakov, A.I.; Samorì, B. Screen-printed electrodes based on carbon nanotubes and cytochrome P450scc for highly sensitive cholesterol biosensors. Biosens. Bioelectron. 2008, 24, 148–150. [Google Scholar] [CrossRef]
- Baj-Rossi, C.; Micheli, G.D.; Carrara, S. Electrochemical detection of anti-breast-cancer agents in human serum by cytochrome P450-coated carbon nanotubes. Sensors 2012, 12, 6520–6537. [Google Scholar] [CrossRef]
- Martin, L.; Latypova, X.; Terro, F. Post-translational modifications of tau protein: Implications for Alzheimer’s disease. Neurochem. Int. 2011, 58, 458–471. [Google Scholar] [CrossRef]
- Permanne, B.; Adessi, C.; Saborio, G.P.; Fraga, S.; Frossaro, M.-J.; Dorpe, J.V.; Dewachter, I.; Banks, W.A.; Leuven, F.V.; Soto, C. Reduction of amyloid load and cerebral damage in a transgenic mouse model of Alzheimer’s disease by treatment with a β-sheet breaker peptide. FASEB J. 2002, 16, 860–862. [Google Scholar] [CrossRef]
- Olanow, C.W.; Brundin, P. Parkinson’s disease and alpha synuclein: Is Parkinson’s disease a prion-like disorder? Mov. Disord. 2013, 28, 31–40. [Google Scholar] [CrossRef]
- Haass, C.; Selkoe, D.J. Soluble protein oligomers in neurodegeneration: Lessons from the Alzheimer’s amyloid β-peptide. Nat. Rev. Mol. Cell Biol. 2007, 8, 101–112. [Google Scholar] [CrossRef]
- Zhao, X.C.; Liu, R.T.; Teng, Y.; Liu, X.F. The interaction between Ag+ and bovine serum albumin: A spectroscopic investigation. Sci. Total Environ. 2011, 409, 892–897. [Google Scholar] [CrossRef] [PubMed]
- Shao, J.; Zhang, X.Y.; Chen, Y.; Zheng, Y. Charge carrier transition in an ambipolar single-molecule junction: Its mechanical-modulation and reversibility. NPJ Comput. Mater. 2016, 2, 2. [Google Scholar] [CrossRef]
- Scullion, L.; Doneux, T.; Bouffier, L.; Fernig, D.G.; Higgins, S.J.; Bethell, D.; Nichols, R.J. Large conductance changes in peptide single molecule junctions controlled by pH. J. Phys. Chem. C 2011, 115, 8361–8368. [Google Scholar] [CrossRef]
- Zhang, X.Y.; Shao, J.; Jiang, S.X.; Wang, B.; Zheng, Y. Structure-dependent electrical conductivity of protein: Its differences between alpha-domain and beta-domain structures. Nanotechnology 2015, 26, 125702. [Google Scholar] [CrossRef] [PubMed]
- Ling, Z.; Zhou, X.; Ahmad, F.; Zhou, Y. Comprehensive spectroscopic probing the interaction and conformation impairment of bovine serum albumin (BSA) by herbicide butachlor. J. Photochem. Photobiol. B 2016, 162, 332–339. [Google Scholar]
- Manouchehri, F.; Izadmanesh, Y.; Aghaee, E.; Ghasemi, J.B. Experimental, computational and chemometrics studies of BSA-vitamin B6 interaction by UV-Vis, FT-IR, fluorescence spectroscopy, molecular dynamics simulation and hard-soft modeling methods. Bioorg. Chem. 2016, 68, 124–136. [Google Scholar] [CrossRef] [PubMed]
- Chovel-Sella, A.; Tov, A.B.; Lahav, E.; Mor, O.; Rudich, H.; Paret, G.; Reif, S. Incidence of rash after amoxicillin treatment in children with infectious mononucleosis. Pediatrics 2013, 131, E1424–E1427. [Google Scholar] [CrossRef] [PubMed]
- Dantzig, A.H.; Bergin, L. Uptake of the cephalosporin, cephalexin, by a dipeptide transport carrier in the human intestinal cell line, Caco-2. Biochim. Biophys. Acta BBA-Biomembr. 1990, 1027, 211–217. [Google Scholar] [CrossRef]
- Ray, W.A.; Murray, K.T.; Hall, K.; Arbogast, P.G.; Stein, C.M. Azithromycin and the risk of cardiovascular death. N. Engl. J. Med. 2012, 366, 1881–1890. [Google Scholar] [CrossRef]
- Kim, M.J.; Park, S.C.; Choi, S.O. Dual-nozzle spray deposition process for improving the stability of proteins in polymer microneedles. RSC Adv. 2017, 7, 55350–55359. [Google Scholar] [CrossRef] [Green Version]
- Ron, I.; Sepunaru, L.; Itzhakov, S.; Belenkova, T.; Friedman, N.; Pecht, I.; Sheves, M.; Cahen, D. Proteins as electronic materials: Electron transport through solid-state protein monolayer junctions. J. Am. Chem. Soc. 2010, 132, 4131–4140. [Google Scholar] [CrossRef] [PubMed]
- Yasmeen, S.; Rabbani, G. Calorimetric and spectroscopic binding studies of amoxicillin with human serum albumin. J. Therm. Anal. Calorim. 2017, 127, 1445–1455. [Google Scholar] [CrossRef]
- Li, Q.; Zhang, T.; Bian, L. Recognition and binding of β-lactam antibiotics to bovine serum albumin by frontal affinity chromatography in combination with spectroscopy and molecular docking. J. Chromatogr. B 2016, 1014, 90–101. [Google Scholar] [CrossRef]
- Wu, Y.; Ji, X.; Hu, S. Studies on electrochemical oxidation of azithromycin and its interaction with bovine serum albumin. Bioelectrochemistry 2004, 64, 91–97. [Google Scholar] [CrossRef] [PubMed]
- Sreerama, N.; Venyaminov, S.Y.U.; Woody, R.W. Estimation of the number of α-helical and β-strand segments in proteins using circular dichroism spectroscopy. Protein Sci. 1999, 8, 370–380. [Google Scholar] [CrossRef] [PubMed]
- Mikayama, T.; Miyashita, T.; Iida, K.; Suemori, Y.; Nango, M. Electron transfer mediated by photosynthetic reaction center proteins between two chemical-modified metal electrodes. Mol. Cryst. Liq. Cryst. 2006, 445, 291–296. [Google Scholar] [CrossRef]
- Khatun, S.; Riyazuddeen. Interaction of colchicine with BSA: Spectroscopic, calorimetric and molecular modeling approaches. J. Biomol. Struct. Dyn. 2018, 36, 3122–3129. [Google Scholar] [CrossRef]
- Nerli, B.; Romanini, D.; Picó, G. Structural specificity requirements in the binding of beta lactam antibiotics to human serum albumin. Chem. Biol. Interact. 1997, 104, 179–202. [Google Scholar] [CrossRef]
- Schneider, E.K.; Huang, J.X.; Carbone, V.; Baker, M.; Azad, M.A.; Cooper, M.A.; Li, J.; Velkov, T. Drug-drug plasma protein binding interactions of ivacaftor. J. Mol. Recognit. 2015, 28, 339–348. [Google Scholar] [CrossRef]
- Yang, S.T.; Lim, S.I.; Kiessling, V.; Kwon, I.; Tamm, L.K. Site-specific fluorescent labeling to visualize membrane translocation of a myristoyl switch protein. Sci. Rep. 2016, 6, 32866. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Henzler Wildman, K.A.; Lee, D.K.; Ramamoorthy, A. Determination of α-helix and β-sheet stability in the solid state: A solid-state NMR investigation of poly (L-alanine). Biopolym. Orig. Res. Biomol. 2002, 64, 246–254. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Wang, X.; Wang, L.; Hou, X.; Liu, W.; Chen, C. Interaction of gold nanoparticles with proteins and cells. Sci. Technol. Adv. Mater. 2015, 16, 034610. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Jager, H.J.; Prinsloo, L.C. The dehydration of phosphates monitored by DSC/TGA and in situ Raman spectroscopy. Thermochim. Acta 2001, 376, 187–196. [Google Scholar] [CrossRef]
- K Abou-Zied, O. Understanding the physical and chemical nature of the warfarin drug binding site in human serum albumin: Experimental and theoretical studies. Curr. Pharm. Des. 2015, 21, 1800–1816. [Google Scholar] [CrossRef]
- Sun, Y.; Huang, Y. Disulfide-crosslinked albumin hydrogels. J. Mater. Chem. B 2016, 4, 2768–2775. [Google Scholar] [CrossRef]
- Orata, D.; Yusuf Amir, C.N.; Mukabi, M. Electrochemical Characterization of Amoxycillin, a Broad Spectrum Antibiotic on a Bentonite Host Matrix, Using Cyclic Voltammetry. IOSR Appl. Chem. 2014, 7. [Google Scholar] [CrossRef]
- Mott, N.F. Conduction in non-crystalline materials. Philos. Mag. 1969, 19, 835–852. [Google Scholar] [CrossRef] [Green Version]
- Coward-Kelly, G.; Agbogbo, F.K.; Holtzapple, M.T. Lime treatment of keratinous materials for the generation of highly digestible animal feed: 2. Animal hair. Bioresour. Technol. 2006, 97, 1344–1352. [Google Scholar] [CrossRef]
- Wilting, J.; van der Giesen, W.F.; Janssen, L.H.; Weideman, M.M.; Otagiri, M.; Perrin, J.H. The effect of albumin conformation on the binding of warfarin to human serum albumin. The dependence of the binding of warfarin to human serum albumin on the hydrogen, calcium, and chloride ion concentrations as studied by circular dichroism, fluorescence, and equilibrium dialysis. J. Biol. Chem. 1980, 255, 3032–3037. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| System | a-Helix | β-Sheet | β-Turn | Unordered |
|---|---|---|---|---|
| BSA | 46.7% | 10.4% | 16.3% | 26.3% |
| Amox-BSA (1:1) | 41.6% | 12.7% | 17.7% | 28.3% |
| Amox-BSA (2:1) | 39.9% | 14.0% | 17.7% | 28.5% |
| Amox-BSA (3:1) | 39.0% | 15.8% | 18.9% | 28.7% |
| BSA | 46.0% | 12.5% | 17.2% | 25.7% |
| Cefa-BSA (1:1) | 45.1% | 12.9% | 17.1% | 27.4% |
| Cefa-BSA (2:1) | 43.0% | 14.6% | 17.4% | 26.3% |
| Cefa-BSA (3:1) | 41.9% | 14.8% | 17.5% | 26.3% |
| BSA | 46.3% | 14.0% | 16.7% | 25.5% |
| Azit-BSA (1:1) | 44.0% | 14.1% | 17.8% | 27.1% |
| Azit-BSA (2:1) | 42.8% | 14.4% | 17.2% | 26.8% |
| Azit-BSA (3:1) | 41.4% | 14.7% | 17.0% | 25.5% |
| I (nA) | 0–1 | 1–2 | 2–3 | 3–4 | 4–5 | 5–6 | >6 | |
|---|---|---|---|---|---|---|---|---|
| Counts | ||||||||
| BSA | 13 | 8 | 2 | 2 | 0 | 1 | 24 | |
| Amox-BSA (1:1) | 16 | 9 | 2 | 2 | 2 | 3 | 16 | |
| Amox-BSA (2:1) | 22 | 4 | 5 | 2 | 2 | 0 | 15 | |
| Amox-BSA (3:1) | 23 | 6 | 4 | 2 | 0 | 1 | 14 | |
| Cefa-BSA (1:1) | 20 | 7 | 6 | 1 | 0 | 0 | 15 | |
| Cefa-BSA (2:1) | 21 | 11 | 3 | 1 | 1 | 3 | 10 | |
| Cefa-BSA (3:1) | 25 | 10 | 1 | 2 | 1 | 2 | 9 | |
| Azit-BSA (1:1) | 23 | 4 | 4 | 1 | 0 | 2 | 16 | |
| Azit-BSA (2:1) | 25 | 6 | 2 | 4 | 2 | 0 | 11 | |
| Azit-BSA (3:1) | 28 | 6 | 3 | 1 | 1 | 3 | 8 | |
| System | Molar Ratio Change | Relative θ Change (%) | Relative Counts Change (%) |
|---|---|---|---|
| Amox-BSA | 0:1 to 3:1 | 19.16 | 41.67 |
| Cefa-BSA | 0:1 to 3:1 | 16.88 | 62.50 |
| Azit-BSA | 0:1 to 3:1 | 18.70 | 66.70 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, J.; Chen, Y.; Xiong, L.; Zhang, X.; Zheng, Y. Conductance Changes in Bovine Serum Albumin Caused by Drug-Binding Triggered Structural Transitions. Materials 2019, 12, 1022. https://doi.org/10.3390/ma12071022
Yu J, Chen Y, Xiong L, Zhang X, Zheng Y. Conductance Changes in Bovine Serum Albumin Caused by Drug-Binding Triggered Structural Transitions. Materials. 2019; 12(7):1022. https://doi.org/10.3390/ma12071022
Chicago/Turabian StyleYu, Jing, Yun Chen, Liqun Xiong, Xiaoyue Zhang, and Yue Zheng. 2019. "Conductance Changes in Bovine Serum Albumin Caused by Drug-Binding Triggered Structural Transitions" Materials 12, no. 7: 1022. https://doi.org/10.3390/ma12071022