
Human Dental Pulp Cells Responses to Apatite Precipitation from Dicalcium Silicates



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Specimen Preparation
2.2. Apatite Precipitate
2.3. Ion Concentration
2.4. HDPCs Isolation and Culture
2.5. COL and FN Secretion
2.6. Cell Adhesion and Proliferation Assay
2.7. Cell Morphology
2.8. Immunofluorescence Staining
2.9. Osteocalcin Formation
2.10. Statistical Analysis
3. Results
3.1. Characterization of CS in Different DMEM


3.2. Ion Concentration

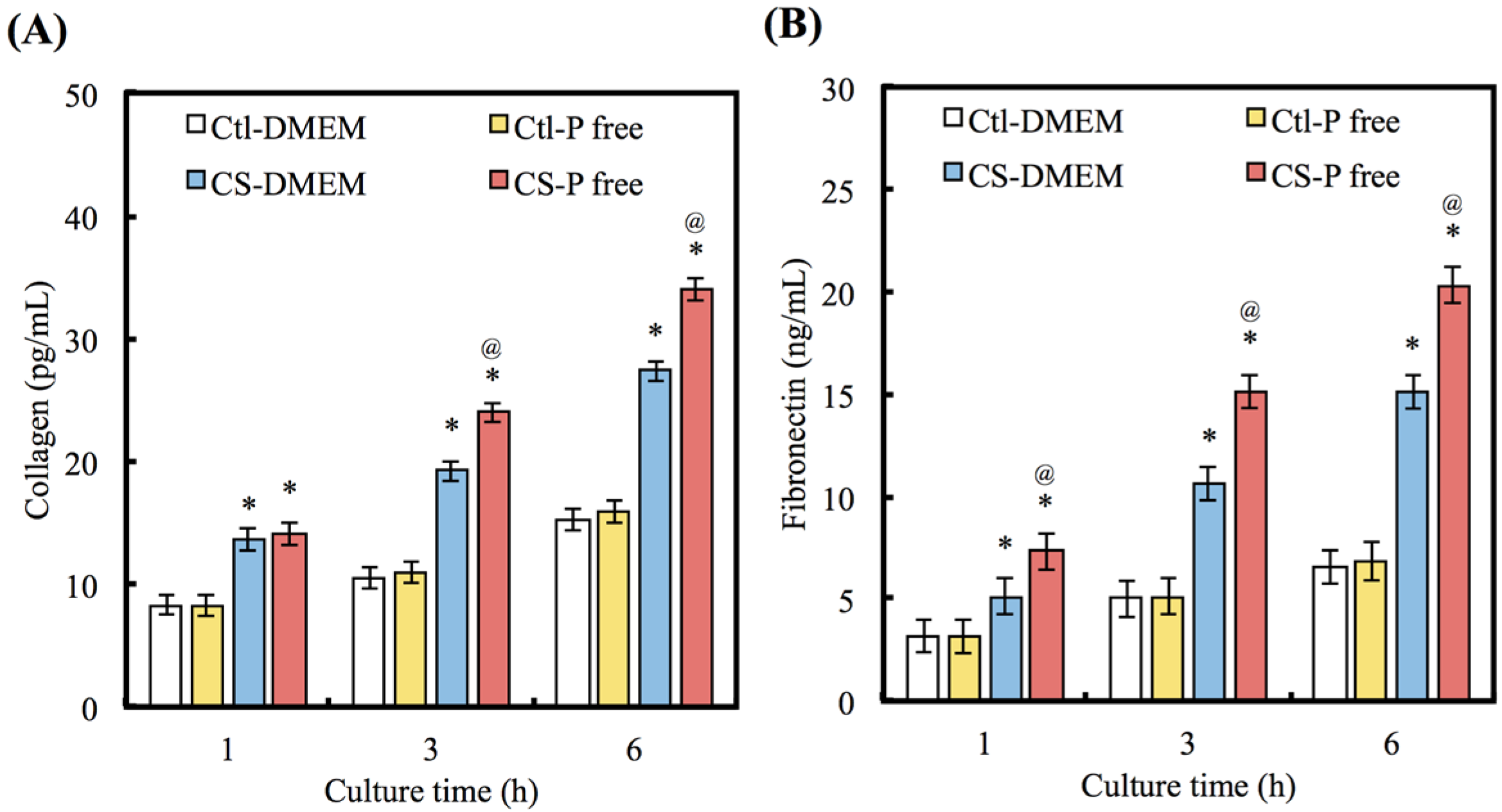
3.3. COL and FN Secretion
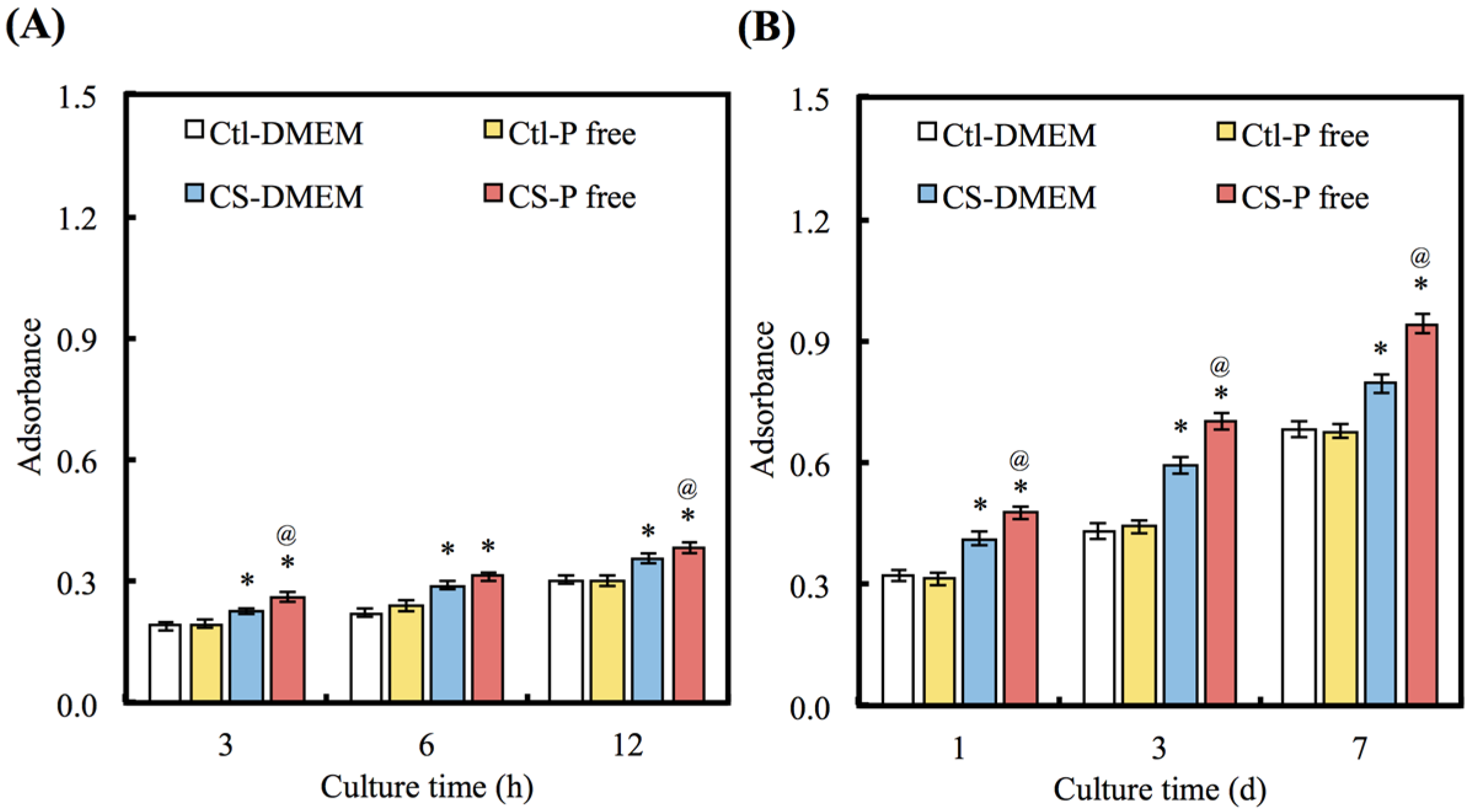
3.4. Cell Adhesion and Proliferation


3.5. Cell Morphology

3.6. Osteocalcin Expression

4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Torabinejad, M.; Watson, T.; Ford, T. Sealing ability of a mineral trioxide aggregate when used as a root end filling material. J. Endod. 1993, 19, 591–595. [Google Scholar] [CrossRef]
- Luo, Z.; Kohli, M.R.; Yu, Q.; Kim, S.; Qu, T.; He, W.X. Biodentine induces human dental pulp stem cell differentiation through mitogen-activated protein kinase and calcium-/calmodulin-dependent protein kinase II pathways. J. Endod. 2014, 40, 937–942. [Google Scholar] [CrossRef] [PubMed]
- Chang, S.W.; Lee, S.Y.; Kum, K.Y.; Kim, E.C. Effects of ProRoot MTA, Bioaggregate, and Micromega MTA on odontoblastic differentiation in human dental pulp cells. J. Endod. 2014, 40, 113–118. [Google Scholar] [CrossRef] [PubMed]
- Wei, W.; Qi, Y.; Nikonov, S.Y.; Niu, L.; Messer, R.L.W.; Mao, J.; Primus, C.M.; Pashbley, D.H.; Tay, F.R. Effects of an experimental calcium aluminosilicate cement on the viability of murine odontoblast-like cells. J. Endod. 2012, 38, 936–942. [Google Scholar] [CrossRef] [PubMed]
- Huang, T.H.; Shie, M.Y.; Kao, C.T.; Ding, S.J. The effect of setting accelerator on properties of mineral trioxide aggregate. J. Endod. 2008, 34, 590–593. [Google Scholar] [CrossRef] [PubMed]
- Kao, C.T.; Shie, M.Y.; Huang, T.H.; Ding, S.J. Properties of an accelerated mineral trioxide aggregate-like root-end filling material. J. Endod. 2009, 35, 239–242. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.H.; Hung, C.J.; Huang, T.H.; Lin, C.C.; Kao, C.T.; Shie, M.Y. Odontogenic differentiation of human dental pulp cells by calcium silicate materials stimulating via FGFR/ERK signaling pathway. Mater. Sci. Eng. C Mater. Biol. Appl. 2014, 43, 359–366. [Google Scholar] [CrossRef] [PubMed]
- Wu, B.C.; Youn, S.C.; Kao, C.T.; Huang, S.C.; Hung, C.J.; Chou, M.Y.; Huang, T.H.; Shie, M.Y. The effects of calcium silicate cement/fibroblast growth factor-2 composite on osteogenesis accelerator in human dental pulp cells. J. Dent. Sci. 2015, 10, 145–153. [Google Scholar] [CrossRef]
- Chen, C.L.; Kao, C.T.; Ding, S.J.; Shie, M.Y.; Huang, T.H. Expression of the inflammatory marker cyclooxygenase-2 in dental pulp cells cultured with mineral trioxide aggregate or calcium silicate cements. J. Endod. 2010, 36, 465–468. [Google Scholar] [CrossRef] [PubMed]
- Shie, M.Y.; Ding, S.J. Integrin binding and MAPK signal pathways in primary cell responses to surface chemistry of calcium silicate cements. Biomaterials 2013, 34, 6589–6606. [Google Scholar] [CrossRef] [PubMed]
- Hung, C.J.; Kao, C.T.; Shie, M.Y.; Huang, T.H. Comparison of host inflammatory responses between calcium-silicate base material and intermediate restorative material. J. Dent. Sci. 2014, 9, 158–164. [Google Scholar] [CrossRef]
- Hung, C.J.; Kao, C.T.; Chen, Y.J.; Shie, M.Y.; Huang, T.H. Antiosteoclastogenic activity of silicate-based materials antagonizing receptor activator for nuclear factor kappaB ligand-induced osteoclast differentiation of murine marcophages. J. Endod. 2013, 39, 1557–1561. [Google Scholar] [CrossRef] [PubMed]
- Chou, M.Y.; Kao, C.T.; Hung, C.J.; Huang, T.H.; Huang, S.C.; Shie, M.Y.; Wu, B.C. Role of the p38 pathway in calcium silicate cement-induced cell viability and angiogenesis-related proteins of human dental pulp cell in vitro. J. Endod. 2014, 40, 818–824. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.C.; Wu, B.C.; Kao, C.T.; Huang, T.H.; Hung, C.J.; Shie, M.Y. Role of the p38 pathway in mineral trioxide aggregate-induced cell viability and angiogenesis-related proteins of dental pulp cell in vitro. Int. Endod. J. 2015, 48, 236–245. [Google Scholar] [CrossRef] [PubMed]
- Hung, C.J.; Hsu, H.I.; Lin, C.C.; Huang, T.H.; Wu, B.C.; Kao, C.T.; Shie, M.Y. The role of integrin αv in proliferation and differentiation of human dental pulp cell response to calcium silicate cement. J. Endod. 2014, 40, 1802–1809. [Google Scholar] [CrossRef] [PubMed]
- Wu, B.C.; Kao, C.T.; Huang, T.H.; Hung, C.J.; Shie, M.Y.; Chung, H.Y. Effect of verapamil, a calcium channel blocker, on the odontogenic activity of human dental pulp cells cultured with silicate-based materials. J. Endod. 2014, 40, 1105–1111. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Wu, C.; Chang, J.; Sun, J. Stimulation of osteogenic protein expression for rat bone marrow stromal cells involved in the ERK signalling pathway by the ions released from Ca7Si2P2O16 bioceramics. J. Mater. Chem. B 2014, 2, 885–891. [Google Scholar] [CrossRef]
- Zhou, Y.; Wu, C.; Zhang, X.; Han, P.; Xiao, Y. The ionic products from bredigite bioceramics induced cementogenic differentiation of periodontal ligament cells via activation of the Wnt/β-catenin signalling pathway. J. Mater. Chem. B 2013, 1, 3380–3389. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Han, P.; Jaiprakash, A.; Wu, C.; Xiao, Y. A stimulatory effect of Ca3ZrSi2O9 bioceramics on cementogenic/osteogenic differentiation of periodontal ligament cells. J. Mater. Chem. B 2014, 2, 1415–1423. [Google Scholar] [CrossRef]
- Huang, S.C.; Wu, B.C.; Ding, S.J. Stem cell differentiation-induced calcium silicate cement with bacteriostatic activity. J. Mater. Chem. B 2015, 3, 570–580. [Google Scholar] [CrossRef]
- Kao, C.T.; Huang, T.H.; Chen, Y.J.; Hung, C.J.; Lin, C.C.; Shie, M.Y. Using calcium silicate to regulate the physicochemical and biological properties when using β-tricalcium phosphate as bone cement. Mater. Sci. Eng. C Mater. Biol. Appl. 2014, 43, 126–134. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Lin, K.; Wu, C.; Liu, X.; Chang, J. Preparation of hierarchical enamel-like structures from nano- to macro-scale, regulated by inorganic templates derived from enamel. J. Mater. Chem. B 2015, 3, 65–71. [Google Scholar] [CrossRef]
- Su, C.C.; Kao, C.T.; Hung, C.J.; Chen, Y.J.; Huang, T.H.; Shie, M.Y. Regulation of physicochemical properties, osteogenesis activity, and fibroblast growth factor-2 release ability of β-tricalcium phosphate for bone cement by calcium silicate. Mater. Sci. Eng. C Mater. Biol. Appl. 2014, 37, 156–163. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.H.; Huang, T.H.; Hung, C.J.; Lai, W.Y.; Kao, C.T.; Shie, M.Y. The synergistic effects of fibroblast growth factor-2 and mineral trioxide aggregate on an osteogenic accelerator in vitro. Int. Endod. J. 2014, 47, 843–853. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.H.; Chen, Y.J.; Kao, C.T.; Lin, C.C.; Huang, T.H.; Shie, M.Y. Physicochemical properties and biocompatibility of silica doped β-tricalcium phosphate for bone cement. J. Dent. Sci. 2014. [Google Scholar] [CrossRef]
- Shie, M.Y.; Ding, S.J.; Chang, H.C. The role of silicon in osteoblast-like cell proliferation and apoptosis. Acta. Biomater. 2011, 7, 2604–2614. [Google Scholar] [CrossRef] [PubMed]
- Lai, W.Y.; Kao, C.T.; Hung, C.J.; Huang, T.H.; Shie, M.Y. An evaluation of the inflammatory response of lipopolysaccharide-treated primary dental pulp cells with regard to calcium silicate-based cements. Int. J. Oral Sci. 2014, 6, 94–98. [Google Scholar] [CrossRef] [PubMed]
- Hsu, T.T.; Kao, C.T.; Chen, Y.W.; Huang, T.H.; Yang, J.J.; Shie, M.Y. The synergistic effects of CO2 laser treatment with calcium silicate cement of antibacterial, osteogenesis and cementogenesis efficacy. Laser Phys. Lett. 2015, 12. [Google Scholar] [CrossRef]
- Hsu, T.T.; Yeh, C.H.; Kao, C.T.; Chen, Y.W.; Huang, T.H.; Yang, J.J.; Shie, M.Y. Antibacterial and odontogenesis efficacy of mineral trioxide aggregate combined with CO2 laser treatment. J. Endod. 2015, 41, 1073–1080. [Google Scholar] [CrossRef] [PubMed]
- Vohra, S.; Hennessy, K.M.; Sawyer, A.A.; Zhuo, Y.; Bellis, S.L. Comparison of mesenchymal stem cell and osteosarcoma cell adhesion to hydroxyapatite. J. Mater. Sci. Mater. Med. 2008, 19, 3567–3574. [Google Scholar] [CrossRef] [PubMed]
- Shie, M.Y.; Chang, H.C.; Ding, S.J. Composition-dependent protein secretion and integrin level of osteoblastic cell on calcium silicate cements. J. Biomed. Mater. Res. Part. A 2014, 102, 769–780. [Google Scholar] [CrossRef] [PubMed]
- Yeh, C.H.; Chen, Y.W.; Shie, M.Y.; Fang, H.Y. Poly(dopamine)-assisted immobilization of Xu Duan on 3D printed poly(lactic acid) scaffolds to up-regulate osteogenic and angiogenic markers of bone marrow stem cells. Materials 2015, 8, 4299–4315. [Google Scholar] [CrossRef]
- Kao, C.T.; Lin, C.C.; Chen, Y.W.; Yeh, C.H.; Fang, H.Y.; Shie, M.Y. Poly(dopamine) coating of 3D printed poly(lactic acid) scaffolds for bone tissue engineering. Mater. Sci. Eng. C Mater. Biol. Appl. 2015, 56, 165–173. [Google Scholar] [CrossRef]
- Lee, M.H.; You, C.; Kim, K.H. Combined effect of a microporous layer and type I collagen coating on a biphasic calcium phosphate scaffold for bone tissue engineering. Materials 2015, 8, 1150–1161. [Google Scholar] [CrossRef]
- Su, Y.F.; Lin, C.C.; Huang, T.H.; Chou, M.Y.; Yang, J.J.; Shie, M.Y. Osteogenesis and angiogenesis properties of dental pulp cell on novel injectable tricalcium phosphate cement by silica doped. Mater. Sci. Eng. C Mater. Biol. Appl. 2014, 42, 672–680. [Google Scholar] [CrossRef] [PubMed]
- Kang, Y.; Kim, S.H.; Bishop, J.; Khademhosseini, A.; Yang, Y.J. The osteogenic differentiation of human bone marrow MSCs on HUVEC-derived ECM and β-TCP scaffold. Biomaterials 2012, 33, 6998–7007. [Google Scholar] [CrossRef] [PubMed]
- Huang, M.H.; Kao, C.T.; Chen, Y.W.; Hsu, T.T.; Shieh, D.E.; Huang, T.H.; Shie, M.Y. The synergistic effects of chinese herb and injectable calcium silicate/b-tricalcium phosphate composite on an osteogenic accelerator in vitro. J. Mater. Sci. Mater. Med. 2015, 26. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.J.; Shie, M.Y.; Hung, C.J.; Wu, B.C.; Liu, S.L.; Huang, T.H.; Kao, C.T. Activation of focal adhesion kinase induces extracellular signal-regulated kinase-mediated osteogenesis in tensile force-subjected periodontal ligament fibroblasts but not in osteoblasts. J. Bone Miner. Metab. 2013, 32, 671–682. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Li, D.; Li, T.; Ding, J.; Liu, J.; Li, B.; Chen, X. Gelatin tight-coated poly(lactide-co-glycolide) scaffold incorporating rhBMP-2 for bone tissue engineering. Materials 2015, 8, 1009–1026. [Google Scholar] [CrossRef]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lai, W.-Y.; Chen, Y.-W.; Kao, C.-T.; Hsu, T.-T.; Huang, T.-H.; Shie, M.-Y. Human Dental Pulp Cells Responses to Apatite Precipitation from Dicalcium Silicates. Materials 2015, 8, 4491-4504. https://doi.org/10.3390/ma8074491
Lai W-Y, Chen Y-W, Kao C-T, Hsu T-T, Huang T-H, Shie M-Y. Human Dental Pulp Cells Responses to Apatite Precipitation from Dicalcium Silicates. Materials. 2015; 8(7):4491-4504. https://doi.org/10.3390/ma8074491
Chicago/Turabian StyleLai, Wei-Yun, Yi-Wen Chen, Chia-Tze Kao, Tuan-Ti Hsu, Tsui-Hsien Huang, and Ming-You Shie. 2015. "Human Dental Pulp Cells Responses to Apatite Precipitation from Dicalcium Silicates" Materials 8, no. 7: 4491-4504. https://doi.org/10.3390/ma8074491