Abstract
Using Hidden Markov Models (HMMs) as a recognition framework for automatic classification of animal vocalizations has a number of benefits, including the ability to handle duration variability through nonlinear time alignment, the ability to incorporate complex language or recognition constraints, and easy extendibility to continuous recognition and detection domains. In this work, we apply HMMs to several different species and bioacoustic tasks using generalized spectral features that can be easily adjusted across species and HMM network topologies suited to each task. This experimental work includes a simple call type classification task using one HMM per vocalization for repertoire analysis of Asian elephants, a language-constrained song recognition task using syllable models as base units for ortolan bunting vocalizations, and a stress stimulus differentiation task in poultry vocalizations using a non-sequential model via a one-state HMM with Gaussian mixtures. Results show strong performance across all tasks and illustrate the flexibility of the HMM framework for a variety of species, vocalization types, and analysis tasks.
1. Introduction
Within the field of bioacoustics, there is a growing need for accurate automatic signal classification for tasks such as call-type classification, individual identification, and assessment of correlation between vocalization patterns and specific social or behavioral contexts. There are a number of well-established techniques for vocalization classification, both template-based and parameter-based. Of the template-based methods, many require significant hand-tuning and have difficulty with temporal and individual variability. In recent years, there has been work toward application of more advanced pattern recognition methods such as those used in human speech and speaker recognition, in particular Hidden Markov Models (HMMs). The work presented here focuses on the use of HMMs as a robust recognition framework for bioacoustic signal classification across a variety of species and classification tasks.
Existing methods for bioacoustic signal classification include multivariate feature analysis, spectrogram cross-correlation, matched filtering, neural networks, dynamic time warping, and others. The most well-established of these is spectrogram cross-correlation (SPCC) [1,2], in which a spectrogram (time-sequence of short-time Fourier transforms) is directly correlated with test vocalizations. Spectrogram cross-correlation is simple to implement, requiring only a single vocalization example of each type of call to be recognized, and can be easily applied to either the isolated vocalization task, where recordings have been presegmented into separate files, or to continuous detection, where a sliding window is applied across a long recording, with correlation peaks indicating target detection. This SPCC method is well-suited to recognizing animal sounds consisting of tones and frequency sweeps [3]. However, the cross correlation approach is unable to adapt to changes in call duration and alignment, and is also significantly impacted by frequency variation such as shifts caused by vocal individuality across callers.
Matched filtering (MF) is a time-series correlation method that uses synthetic waveforms [4] or synthetic spectrograms [3] instead of recording examples. This approach is also easy to implement, although requiring more effort to construct pattern templates, and has many of the same strengths and weaknesses as spectrogram cross-correlation. Artificial Neural Network (ANN) approaches can be used for classification in many different applications, depending on the specific choice of input features and network topologies. If spectrogram values are used as input features, a common configuration, then ANNs can adjust the emphasis given to various portions of the time-frequency template, giving improved classification for some tasks such as species detection [5,6,7]. However, in this configuration, ANNs are subject to some of the same temporal nonlinearity and frequency shifting weaknesses as SPCC and MF. Dynamic time warping (DTW) [8], commonly used in small-vocabulary isolated word recognition for human speech, is able to handle temporal nonlinearity through optimal frame-to-frame alignment of the reference template with test vocalizations. However, it is difficult to implement DTW in a continuous detection framework, which is one of the reasons it is now rarely used for large-scale speech recognition.
HMMs, now used in nearly all state-of-the-art speech recognition systems, have begun to gain attention in bioacoustics as well [9,10,11,12]. An HMM is a statistical state machine model where each state represents a stationary spectral configuration, and transitions between states represent spectral changes over time. Benefits of HMMs include their ability to handle nonlinear temporal alignment, their statistical basis for classification, their extendibility to continuous recognition or detection, and the ease with which they can incorporate complex language or other recognition constraints.
The performance of HMM-based systems has been compared to other classification methods for bioacoustic tasks. Weisburn et al. [13] compared the performance of a matched filter and an HMM system for detecting bowhead whale (Balaena mysticetus) call notes. The matched filter used a spectrogram template, while the HMM used the three largest spectral peaks as features for an 18-stage model. The HMM detected 97% of the notes, whereas the MF detected 84%. Anderson [14] compared an HMM approach to a DTW-based system using a dataset consisting of two different species of bird song. His conclusion was that although the DTW system worked better with a small amount of training data, the HMM system was more robust to noise and to vocalization variability.
One of the issues that makes comparison across methods difficult is that of feature extraction. The selection of features is an essential component of any pattern recognition system, often having as much or more impact on the outcome as the choice of classification model. For example, SPCC by definition relies on short-time Fourier transform amplitudes as a feature, whereas MF uses a direct time-series correlation, and DTW can use any spectral distance measure between frames. HMMs, as statistical classification models, are able to use any frame-based feature vector giving meaningful discriminability across classes, most commonly cepstral coefficients. This use of cepstrum coefficients is somewhat different than the standard statistical analysis approaches common in the bioacoustics field, where typical features include whole-vocalization measures such as duration, energy, and maximum, minimum and average fundamental frequency values. Such measures work well with statistical test techniques such as the t-test, Chi-Squared test, MANOVA, and factor analysis. Frame-based features, however, are somewhat more difficult to use with traditional statistical approaches [8,15] because there are a variable number of features per vocalization. Another significant and often unavoidable problem in bioacoustic signal processing is the presence of background noise due to adverse recording environments, as well as convolutional noise due to microphone and sound propagation variability. Standard spectral features are highly sensitive to noise, which can decrease system accuracy and give misleading results. It is important, therefore, to incorporate signal enhancement and/or robust feature extraction techniques that enhance the vocalization while suppressing background noise. Common techniques to reduce noise artifacts in bioacoustic signals include basic bandpass filters and related frequency-based methods for spectrogram filtering and equalization, often incorporated directly into acquisition and analysis tools [16]. Other approaches in recent years have included spectral subtraction [17], minimum mean-squared error (MMSE) estimation [18,19], adaptive line enhancement [20], and perceptually motivated wavelet denoising [21].
In the study presented here, we examine the HMM recognition framework and look at its applicability to several different types of bioacoustic classification tasks. The framework is illustrated with three different datasets, each representing a different kind of task. While the same HMM framework and feature extraction methods are used across all tasks, front-end signal processing methods and implementation details are tailored to each application. Application tasks include call type classification of Asian elephant vocalizations, syllable-based song-type classification of ortolan bunting songs, and age-dependent stress stimulus classification of poultry vocalizations.
2. Methods
2.1. HMMs
An HMM is a stochastic finite state machine used to model a speech segment. The segments may be words, syllables, subword units, or even a complete phrase [22]. Fundamentally, an HMM is simply a set of states connected by transition arcs, as illustrated in Figure 1. When modeling time-sequential patterns, the states of the HMM are typically connected with state transitions from left to right, representing time.

Figure 1.
HMM with GMM state observations aligned to an Asian Elephant Squeak vocalization. The first and last state represent entry and exit points for the model, while internal states, called “emitting” states, are aligned to signal frames to maximize overall model likelihood.
Figure 1.
HMM with GMM state observations aligned to an Asian Elephant Squeak vocalization. The first and last state represent entry and exit points for the model, while internal states, called “emitting” states, are aligned to signal frames to maximize overall model likelihood.
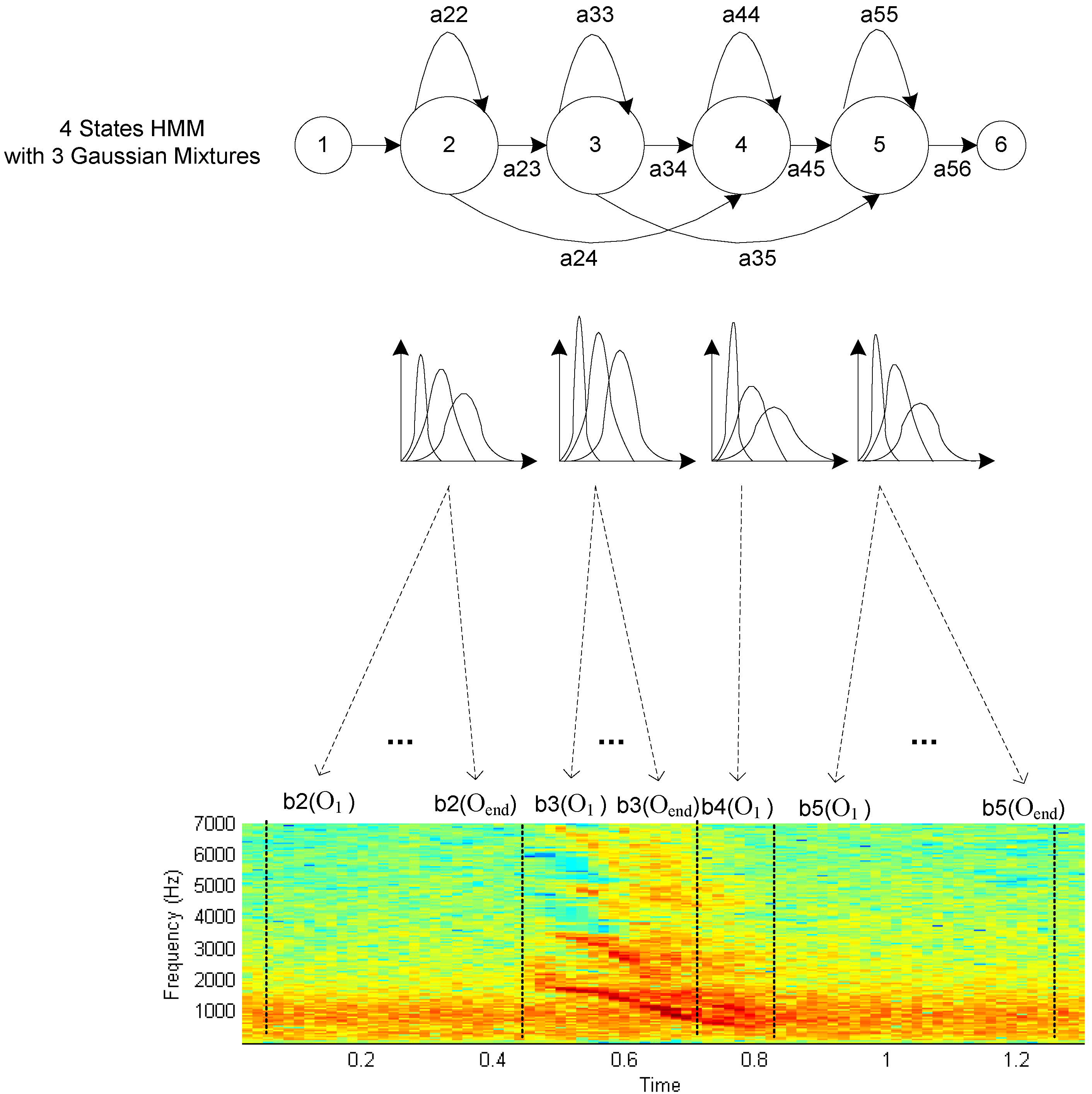
An HMM is defined by its transition matrix A, in which each element ai,j represents the probability of moving from state i to j, and state observation probabilities bi(ot), which represent the probability distributions for each state i. Observations ot represent the data being modeled, in this case the feature vectors of the vocalization frames. Because the transition matrix is two-dimensional, the system has the Markov property, i.e. the probability of the next state is dependent only on the current state. In human speech, Gaussian Mixture Models (GMMs), a weighted sum of Gaussians that can represent a wide variety of unimodal and multimodal distribution characteristics, are commonly used to model the state observation probability densities. The programming toolkit used here is the Hidden Markov Model Toolkit (HTK) from Cambridge University (2002) [23]. Parameter training is implemented using the Baum-Welch method (an implementation of the well known Expectation Maximization algorithm) for maximum likelihood estimation [24,25], and classification is accomplished using the Viterbi algorithm [26] to identify the most likely HMM or HMM sequence given an unknown vocalization.

Figure 2.
GMM model aligned to a non-stress condition poultry vocalization.
Figure 2.
GMM model aligned to a non-stress condition poultry vocalization.
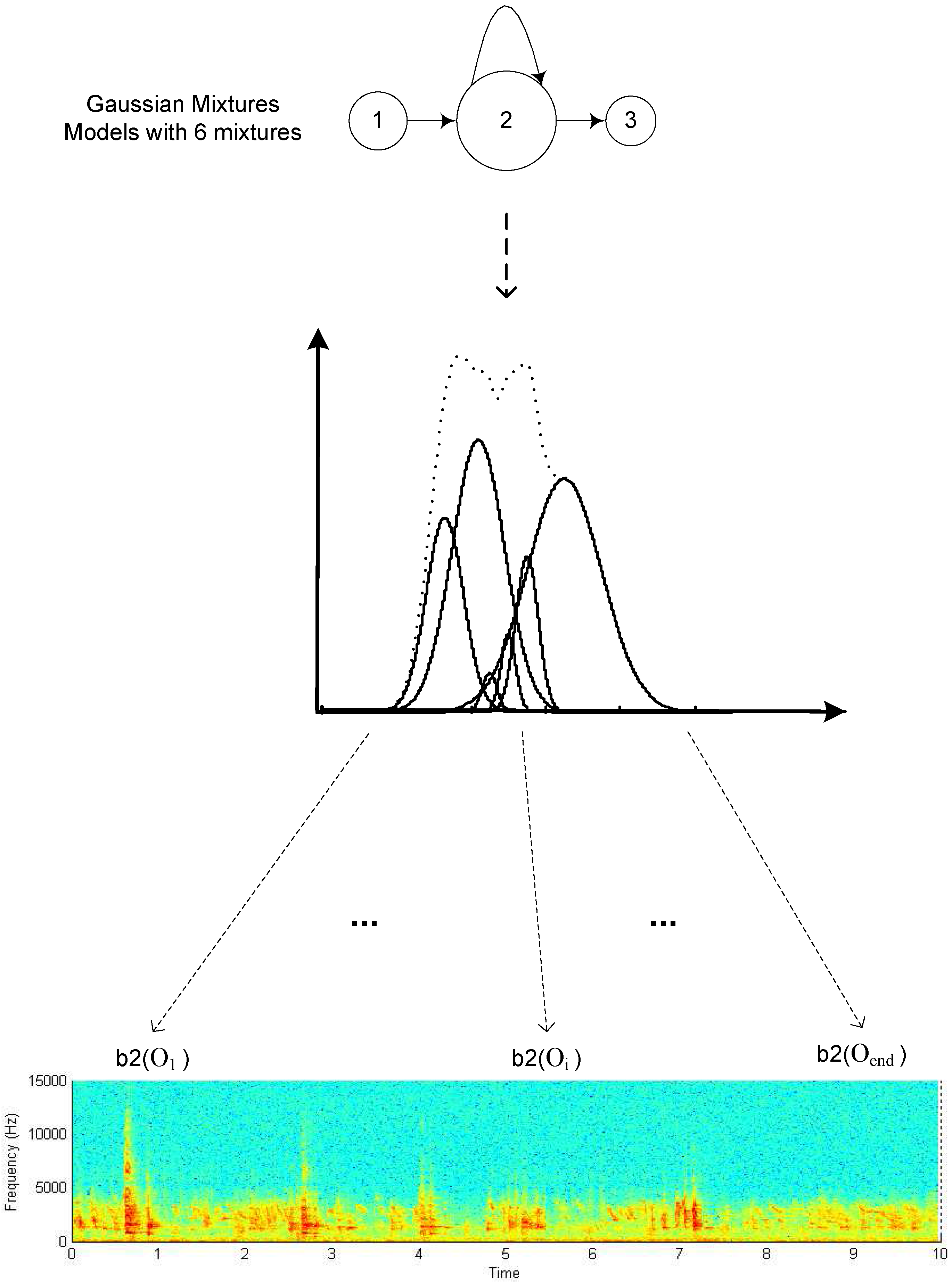
The left to right topology and number of states in the model may be adjusted to accommodate a wide variety of sequential patterns. Patterns with more variation and complexity are typically assigned models with more states to represent the different regimes. In contrast, it is possible to use HMMs to classify patterns without any temporal structure at all by utilizing a single state HMM with a GMM observation model, as illustrated in Figure 2, which reduces it to a simple statistical classifier. Transitions may also be added so that some states may be skipped completely. This is particularly useful in terms of modeling beginning and ending silence regions within vocalizations. Simply by including extra beginning and ending states, arbitrary amounts of surrounding silence regions are implicitly included in the model and there is no need for precise segmentation of individual vocalization start and end points. Accommodating the presence of silence and pauses within vocalizations is another example of temporal nonlinearity that can be easily handled by the HMM framework but is extremely difficult to incorporate into template based models.
2.2. GFCC/GPLP features
Mel-Frequency Cepstral Coefficients (MFCCs) [27] and Perceptual Linear Prediction (PLP) coefficients [28] are well-established feature representations for human speech analysis and recognition tasks. Both of these are representations of the vocal tract spectrum incorporating perceptual models of the human auditory system. The MFCC approach warps the linear frequency axis to match the Mel-scale cochlear frequency map, while the PLP method uses critical band filters, equal-loudness curve amplitude transformation, and cube-root power to intensity transformation. The use of MFCC and PLP coefficients have several beneficial characteristics and have been shown to be robust and highly effective for characterizing human speech for tasks such as speech and speaker recognition. These features, however, are specific to the human auditory system and need to be modified for application across a wider variety of animal species.
The features used in this work are a generalization of the MFCC and PLP features [29] based on the work of Greenwood [30], who showed that many terrestrial and aquatic mammals have a logarithmic cochlear-frequency map. MFCCs have been generalized to Greenwood Function Cepstral Coefficients (GFCCs) by the use of the Greenwood frequency warping function. PLPs have been generalized to generalized perceptual linear prediction (gPLP) coefficients, constructed by substituting a species-specific frequency warping function and equal loudness curve. It has been shown [10,31] that these generalized features are effective for acoustic pattern classification across a wide range of species and applications. The basic calculation method for GFCCs is to place triangular filters over the Fourier Transform of each frame, spaced according to the Greenwood frequency warping function, and then take the filter bank energies and use a Discrete Cosine Transform (DCT) to convert the filter bank energies into cepstral coefficients. In the experiments presented here, a 12-dimensional GFCC vector is used as the base feature, to which signal log energy is appended, after which velocity and acceleration coefficients (referred to as delta and delta-delta coefficients in the speech community) are calculated for each of the 13 original features, yielding an overall 39-element feature vector for each frame. In addition, the feature vectors are normalized using Cepstral Mean Subtraction (CMS), a common technique which simply subtracts the mean value across each vocalization. This compensates for fixed convolutional noise effects like microphone channel variation.
2.3. Language Models
During the recognition process, multiple HMMs can be connected together to form recognition networks. In continuous speech recognition, the fundamental HMM units are often individual phonemes, so that this recognition network represents phoneme-to-word mappings (i.e., a dictionary), pronunciation variants, pauses between words or phonemes, and word-to-word transitions (i.e., a language model). The recognition network also offers a mechanism for directly incorporating language model probabilities. For bioacoustic tasks, this recognition network framework can be used to represent information about the specific number and types of vocalization sequences expected.

Figure 3.
HMM recognition networks. Each unit represents an individual HMM, with the overall network representing allowable HMM sequences (a) Isolated vocalization network, exactly one HMM per vocalization and one vocalization per recording (b) Isolated vocalization network incorporating a constrained language model (c) Constrained model with optional inter-syllable short pauses and beginning/ending silence regions (d) Transformation into continuous detection network.
Figure 3.
HMM recognition networks. Each unit represents an individual HMM, with the overall network representing allowable HMM sequences (a) Isolated vocalization network, exactly one HMM per vocalization and one vocalization per recording (b) Isolated vocalization network incorporating a constrained language model (c) Constrained model with optional inter-syllable short pauses and beginning/ending silence regions (d) Transformation into continuous detection network.
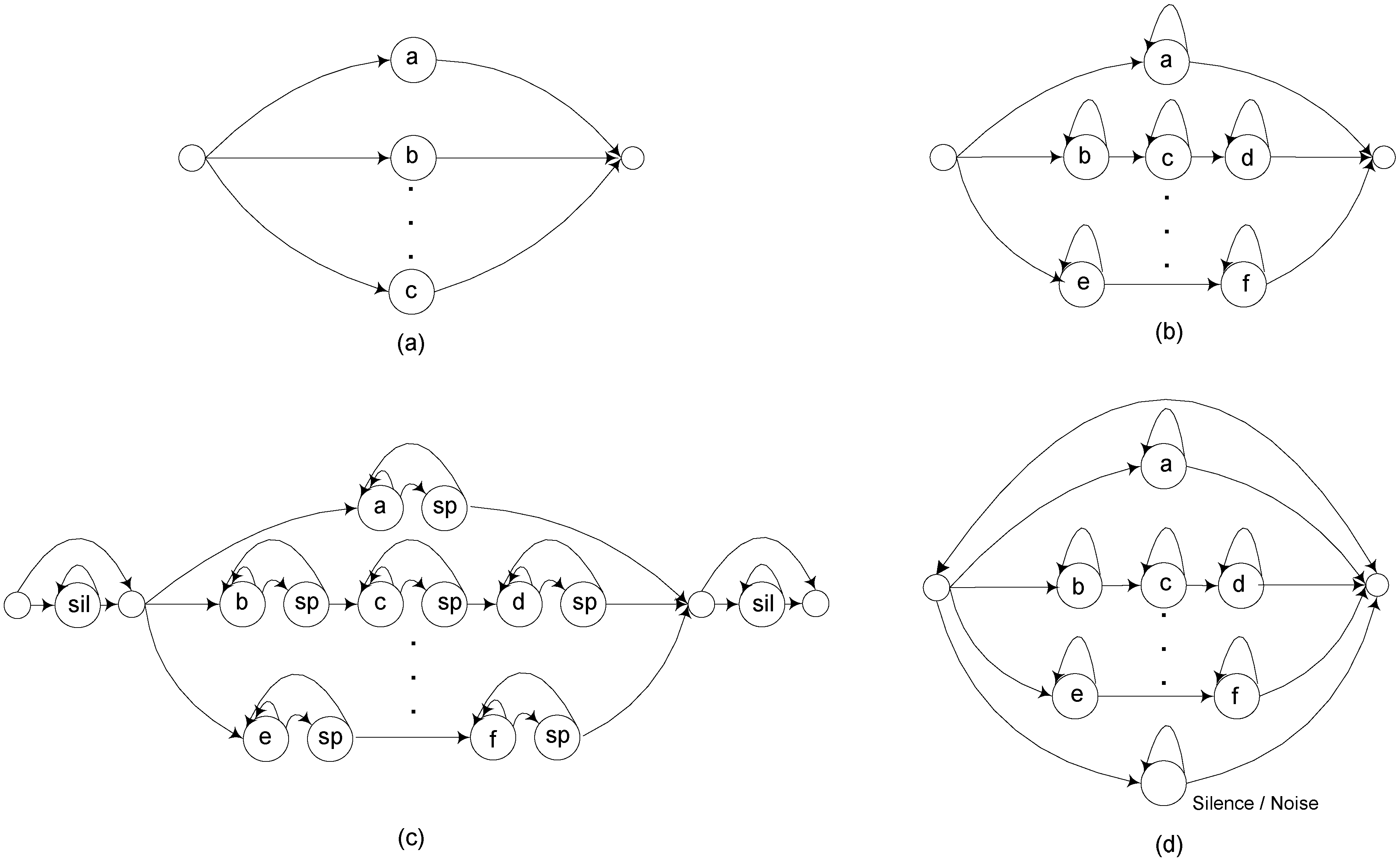
The flexibility of this framework is illustrated by the four different recognition networks shown in Figure 3. The first of these is just a simple single-vocalization classification network, where each HMM represents a vocalization pattern and there is exactly one vocalization per segmented file. The second adds substantial complexity, allowing repeated HMM sequences according to specific patterns. Here, each HMM would typically be a sub-vocalization unit (such as a syllable of a bird song). The third is similar to the second but adds optional short pauses between individual syllables as well as initial beginning and ending silence models for more robust alignment. The final network illustrates the addition of a feedback loop and an optional background noise model in parallel, which changes the individual vocalization recognition network into a continuous detection network that outputs both recognized vocalizations and timestamps across an entire recording. Vocalization and language probabilities can also be incorporated into the network to maximize the effective detection probability.
3. Experiments
3.1. Asian Elephant (Elephas maximus): Repertoire Analysis
Elephants are intelligent, long-living animals that live in a complex and fluid society in which several modes of communication play a role in maintaining group cohesion and social order, and in locating and assessing reproductive state of potential mates [32,33]. Although the social and behavioral contexts of Asian elephant communication signals are well studied [34,35,36,37], the study of the elephant vocal repertoire through categorization of call types by acoustic parameters is still needed as a basis for future research on individuality and for comparisons of acoustic communication among elephant species and populations [32]. McKay [38] described ten basic vocalizations for Asian elephants, divided into three major categories. However, only the low frequency rumble has been described by spectral and temporal features [39].
This study focuses on call type classification for Asian elephant repertoire analysis and validation. Vocalizations were recorded from captive elephants housed at the Oregon Zoo in Portland, OR (USA) and from domesticated elephants in Thailand [40,41]. Vocalizations were collected in a variety of social contexts, time frames, and environmental noise conditions in both urban and rural settings. Data was recorded from seven known individuals and between 10 and 25 unknown individuals, at distances ranging from 1 to 100 meters. The primary sampling rate was 44.1 kHz, although a few calls were alternatively recorded at 32 kHz. Call types were manually classified using perceptual aural cues and visual inspection of spectrograms for differentiation of fundamental frequency contour, tonality, and signal duration. Glaeser et al. [40,41] defined an acoustic repertoire of Asian elephants based on acoustic parameters, and validated structural distinction among six basic call types (Bark, Roar, Rumble, Squeak, Squeal, and Trumpet) and the non-laryngeal Blow. In addition to these basic call types, five call combinations with these basic calls forming their constituent parts are defined (Roar-Rumble, Squeal-Squeak, Squeak train, Squeak-Bark, and Trumpet-Roar). The frequency range of the signal energy ranges from 14 Hz up to 9 kHz, and call durations range from 0.1 to 14 seconds. Trumpets, Squeaks, Squeals have energy in the higher frequency range (54 Hz to 9 kHz) and are differentiated perceptually and through visual inspection by frequency modulation and duration. Rumbles, Roars, and Barks are limited to the lower frequency range (14 Hz to 5 kHz), and are differentiated by amplitude modulation and duration. A total of 2,044 vocalizations were manually segmented into all call categories. All vocalizations are included in the data set, regardless of signal degradation by noise or presence of overlapping signals.
To maximize training set size given the limited number of examples, 5-fold cross validation is implemented. In the cross-validation methodology, the data set is divided into multiple equal subsets (five for this experiment), with one set used for testing and the remaining for training. The process is then repeated once for each test subset, so that each example is used as a test point exactly once.
For analysis, GFCC features are extracted from the vocalizations using a moving Hamming window with a window size of 25 ms with 15 ms overlap. A 39 element feature vector (standard in human speech recognition) is used, with 12 GFCC coefficients with cepstral mean normalization plus log-energy, appended by velocity and acceleration, as described previously. The Greenwood frequency warping constants are calculated using 10 Hz−10 kHz to encompass the signal energy [40,41]. A total of 60 triangular filterbanks (more than is usually used in human speech) are spaced in accordance with the Greenwood warping function across that frequency range to guarantee enough filterbanks in the low frequencies, where a large amount of signal energy is located. One Hidden Markov Model is used to model each of the 11 different call types discussed above, with 12 emitting states per HMM and GMM state distributions with three Gaussian mixtures per state. A range of parameter variations was also implemented for comparative evaluation, with generally similar performance present across a broad range of states and mixtures.

Table 1.
Confusion matrix for 12 state HMM with 3 mixtures. Basic call types are shown in bold. BRK=Bark, RRM=Roar-Rumble, ROR=Roar, RUM=Rumble, SKC=SqueakTrain, SQG=Squeak-Bark, SQK=Squeak, SQL=Squeal, SQS=Squeak-Squeal, TMP=Trumpet, TRR=Trumpet-Roar. Overall accuracy is 60.1% (vs. chance accuracy 28.0%).
| Classification | ||||||||||||
| BRK | RRM | ROR | RUM | SKC | SQG | SQK | SQL | SQS | TMP | TRR | ||
| Label | BRK | 42 | 0 | 10 | 20 | 0 | 0 | 1 | 5 | 0 | 6 | 0 |
| RRM | 0 | 5 | 8 | 4 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |
| ROR | 21 | 5 | 134 | 32 | 0 | 0 | 0 | 11 | 2 | 8 | 0 | |
| RUM | 7 | 1 | 20 | 87 | 1 | 0 | 0 | 32 | 10 | 23 | 0 | |
| SKC | 0 | 0 | 2 | 0 | 64 | 5 | 0 | 10 | 22 | 3 | 0 | |
| SQG | 0 | 0 | 1 | 0 | 8 | 13 | 3 | 2 | 1 | 0 | 0 | |
| SQK | 11 | 0 | 0 | 5 | 26 | 2 | 253 | 16 | 11 | 7 | 0 | |
| SQL | 12 | 0 | 2 | 38 | 17 | 0 | 11 | 323 | 103 | 66 | 0 | |
| SQS | 0 | 0 | 0 | 7 | 26 | 4 | 13 | 75 | 75 | 1 | 0 | |
| TMP | 6 | 0 | 1 | 22 | 2 | 1 | 17 | 24 | 4 | 227 | 0 | |
| TRR | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 0 | 4 | |
The confusion matrix over the full data set conditions, including all call types with all overlapping and noisy calls, are shown in Table 1. The overall accuracy for the full set is 60.1%. The pattern shown by the confusion matrix indicates that a significant number of the errors are made between highly confusable call types and between basic call types and their corresponding call-combinations. By roughly ordering the columns according to confusability between calls, similarity can be observed by looking along the diagonal for blocks with significant error rates. Some of these patterns are highlighted in the confusion matrix for reference. Similarity between RRM (Roar-Rumble) and RUM (Rumble) and to a lesser extent ROR (Roar) is noted, as well as substantial similarity between SQS (Squeak-Squeal) and SQL (Squeal). These similarities are expected because combination calls are made up of the basic call types with which they are confused, and the error patterns help confirm the repertoire categories.
More accurate classification is possible by including only the basic call types, and by excluding calls with high noise degradation and calls that overlap with other calls or sporadic sounds. This improvement is shown in Table 2, where the accuracy for a 12 state HMM classifier with three Gaussian mixtures is 85.0%, across 186 clean vocalizations.
Table 2.
Call type classification accuracy over reduced-set clean Asian Elephant data. Overall accuracy is 85.0% (vs. chance accuracy 32.8%).
| Classification | |||||||
|---|---|---|---|---|---|---|---|
| BRK | ROR | RUM | SQK | SQL | TMP | ||
| Label | BRK | 15 | 0 | 0 | 0 | 4 | 3 |
| ROR | 3 | 22 | 0 | 0 | 1 | 0 | |
| RUM | 1 | 2 | 4 | 0 | 0 | 1 | |
| SQK | 0 | 0 | 0 | 60 | 1 | 0 | |
| SQL | 0 | 1 | 0 | 1 | 26 | 2 | |
| TMP | 0 | 2 | 0 | 3 | 3 | 31 | |
3.2. Ortolan Bunting (Emberiza Hortulana): Syllable, Song Variant, and Song Type Classification
The ortolan bunting is an endangered migratory passerine distributed from Western Europe to Mongolia [42], which has undergone a major population decline in recent years [43,44]. This bird vocalizes in a range between 1.9 kHz and 6.7 kHz and has a relatively simple song and a repertoire size that varies between just a few songs per individual to as many as 24 [45]. The ortolan bunting vocalizations examined for this study were collected from County Hedmark, Norway in May of 2001 and 2002 [46]. The male vocalizations were recorded on 11 out of 25 sites within an area of about 500 km2. The total number of males in the covered area during 2001 and 2002 was approximately 150. As described by Osiejuk [46], these ortolan vocalizations were recorded between 04:00 and 11:00 am with a professional DAT recorder at the sampling rate of 48 kHz.
Songs of ortolan buntings are composed of base units referred to as syllables. Each syllable is described using letter notation, as illustrated in Figure 4. Songs are categorized by song type, defined according to the general type of syllable pattern, and further into specific song variants, defined by exact repetition pattern. For example, common song types include ab, cd, and huf, with common song variants such as aaaab, aaaaabb, cccd, and hhhhuff. Although individual examples of syllables differ in length and frequency across individuals, they maintain the same sonogram shape and underlying temporal pattern.

Figure 4.
Ortolan bunting syllables (from [46]).
Figure 4.
Ortolan bunting syllables (from [46]).
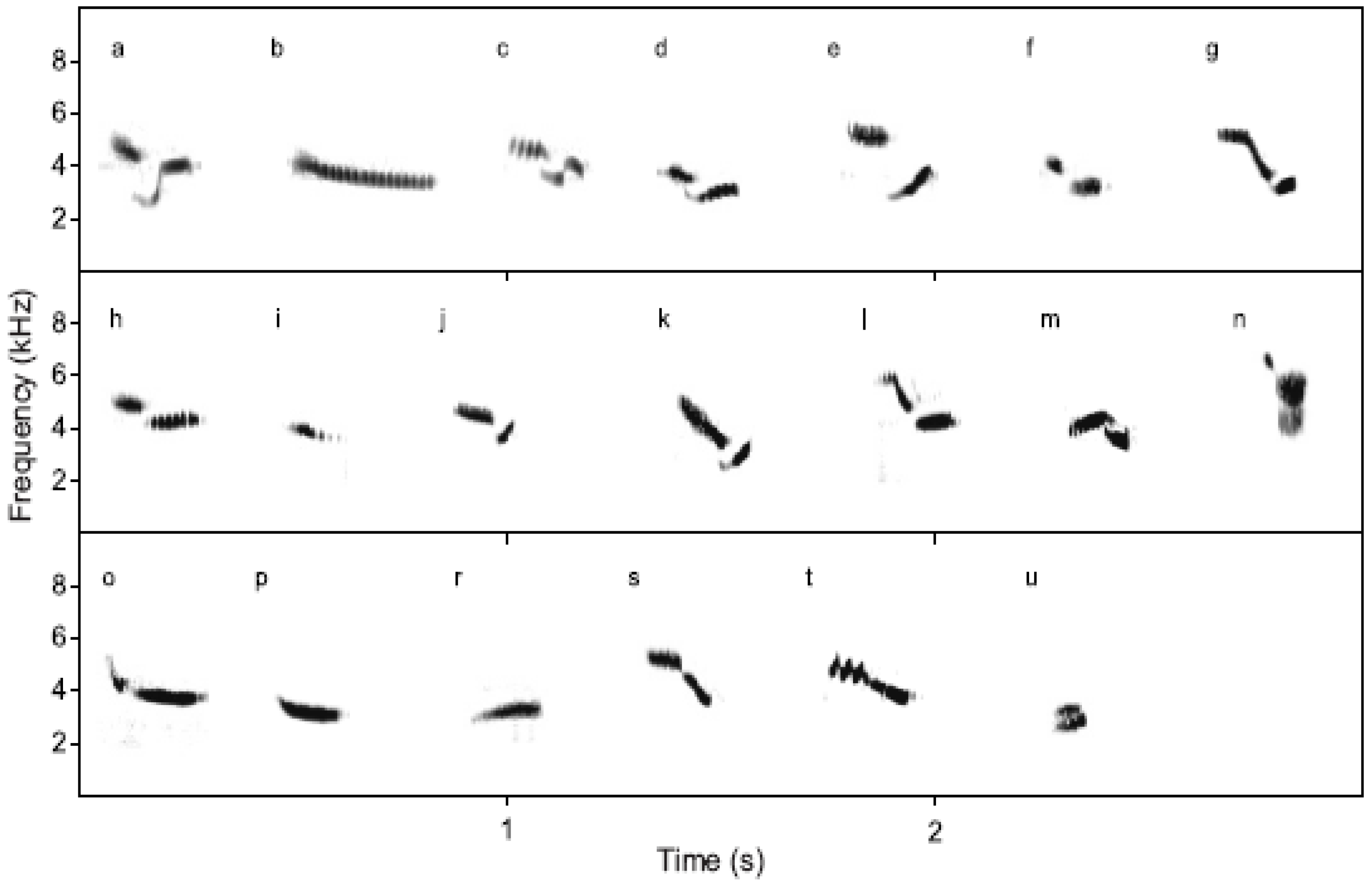
This study focuses on automatic song classification in the ortolan bunting. Modeling is done at the syllable level, so that each syllable type has a corresponding HMM, and then classifying into a syllable sequence to match individual songs with a specific song type and song variant.
To evaluate classification accuracy using HMMs, a subset of 10 frequently produced call types is used, with 25 calls in each type used for training and 25 for testing, and data across multiple individuals included in both the training and test sets. The call types selected include ab, cb, cd, eb, ef, gb, guf, h, huf, and jufb, which consist of the ten common syllables a, b, c, d, e, f, g, h, j, and u. In addition a silence model is trained to represent beginning and ending silence and pauses between syllables.
Due to substantial recording noise, all waveforms are enhanced using simple band-pass filtering to eliminate noise outside the active vocalization range (2,500 to 7,500 Hz) of these calls, followed by Ephraim-Malah filtering [18], a statistical signal enhancement method common in human speech processing applications. Analysis is then done using Hamming windows with a window size of 5 ms with 2.5 ms overlap, accounting for the much faster temporal patterns present in the bunting vocalizations as compared to the Asian elephants. Features for classification are the same 39 element GFCC feature vector described previously for the Asian elephant data, with a few minor differences. The main difference is that the lower and upper frequency limits of the Greenwood function used for frequency warping are adjusted to 2,500 and 7,500 Hz, respectively, to represent the frequency range of these calls. In addition, a simple baseline curve normalization is added to the log energy feature. This consists of performing a second order polynomial fit to the log energy curve for each song, then subtracting the baseline curve from the log energy on a per-frame basis. This is done because the amplitude of the individual syllables is commonly the loudest in the middle portion of calls, and the normalization compensates for the longer-term amplitude changes while still allowing shorter-term signal amplitude modulations to be represented by the energy feature and thus used for classification.
Each syllable model consists of a 15 emitting state HMM with a single Gaussian, designed to track the time-evolution of each syllable’s frequency pattern. Multiple Gaussian mixtures are not necessary because of the simplicity of the frequency characteristics along each step in the time-frequency pattern. Recognition is implemented using a constrained language model describing the allowable song patterns, as illustrated previously in Figure 3.
Classification is evaluated in terms of song-type accuracy, syllable accuracy, and song-variant accuracy. Song-type and song-variant accuracy can be computed directly as the percent of songs with the correct classification. However, since there are multiple syllables in each song, syllable accuracy requires a detailed alignment process, where the recognized song is aligned against the correct song transcription and a dynamic programming method is used to determine the minimal number of insertions, deletions, and substitutions differentiating the recognized song or correct song [23]. Final results are shown in Table 3 below. In addition, Table 4 shows the increase in syllable accuracy as the various system improvements described above are added sequentially, including language modeling constraints, feature normalization and signal enhancement, short pause models between syllables, and energy normalization. It can be seen that language model constraints, feature normalization (which reduces individual vocal variability), and front-end signal enhancement each make very substantial contributions to accuracy improvement.
Table 3.
Song type, syllable, and song variant classification accuracy for overall system.
| Measure | Accuracy |
|---|---|
| Song Type | 100% |
| Syllable | 97.5% |
| Song Variant | 77.6% |
Table 4.
Increase in accuracy due to sequential system improvements. Constrained Recognition Network (CRN) refers to the song-type language model of Figure 3b. Cepstral Mean Subtraction (CMS) is feature normalization as described in Section 2.2. Signal Enhancement (SE) is Bandpass plus Ephraim-Malah filtering as described above. Short Pause (SP) is the song-type language model of Figure 3c, and Energy normalization (EN) is baseline curve energy normalization as described above.
| Methodology (15 state HMMs with 1 Gaussian) | SyllableAccuracy |
|---|---|
| Baseline 39 element feature vector | 61.0% |
| Baseline + CRN (Constrained Recognition Network ) | 74.0% |
| Baseline + CMS (Cepstral Mean Subtraction) | 88.0% |
| Baseline + CRN +CMS + SE (Signal Enhancement) | 95.9% |
| Baseline + CRN + CMS + SE + SP (Short Pause) | 96.7% |
| Baseline + CRN + CMS + SE + SP + EN (Energy Normalization) | 97.5% |
3.3. Chicken (Gallus Domesticus): Age-Dependent Stress Condition Classification
Maintaining low-stress living conditions is extremely important to the care of domesticated poultry used for agricultural food production. However, currently the only clinical tool for directly measuring stress levels requires measuring hormonal indicators in blood samples, which itself stresses the animals. This study is focused on investigating the correlation between vocalization patterns in agricultural chickens and various stress stimuli in this environment, for the purpose of assessing whether vocalizations could be reliably used as a stress indicator. Two separate stress-related task experiments are implemented, one focused on directly detecting living condition stress in vocalizations and one focused on evaluating the connection among stress induced by human presence, diet and age.
Vocalizations were collected at the University of Connecticut Poultry Barn, which houses 25 animals. A stationary microphone to which the animals were accustomed was used. Long (multiple-minute) DAT recordings at a 44.1 kHz sampling rate were made after acclimatization to each condition under test, and then divided into 10 second duration segments for analysis and classification. The vocalizations used in these experiments vary from previous studies in that there are large numbers of individuals vocalizing simultaneously, so that individual vocalizations cannot be separated and there is no discernible time patterning. This is accommodated within the HMM framework by reducing the number of states to 1 because there is no time sequence to be modeled, and increasing the number of mixtures to 12 to get a finer resolution discrimination in terms of overall spectral characteristics across the population. Thus, the classification method is spectral as opposed to sequential, and the HMM approach becomes equivalent to a much simpler GMM statistical classifier.
Feature extraction is again similar to that of the previous tasks, using a base 39-element GFCC feature vector, with Hamming frames of 25 ms each, frame overlap of 15 ms, and 26 filter banks for analysis. The lower and upper frequency limits are set to 125 Hz and 2,000 Hz [47], respectively, based on the approximate hearing range. However, the log energy coefficient is not included since there is no need for a loudness pattern due to the non-temporal model, and to avoid any bias that might be introduced by overall vocalization amplitude in different conditions,
3.3.1. Age dependent living condition detection
The goal of the first task is age-dependent living condition classification. Newly hatched chicks were recorded at the ages of 1, 2 and 3 months, in six different stress factor conditions. These conditions include: No stimulus (Control), Heat (H), Human Presence (HP), Heat + Human Presence (H + HP), Crowding (C), Heat + Crowding (H + C). The baseline Control condition was without any humans present and an ambient temperature of 60 degrees Fahrenheit, the Heat condition was conducted at an ambient temperature of 100 degrees Fahrenheit, the Human Presence condition was conducted with one individual present in the barn, and the Crowding condition was conducted with space per chicken reduced by 1/3 over standard recommended conditions. A total of 1,355 vocalization segments were recorded across the three age groups and six conditions.
Classification is run over each age category separately using 10-fold cross validation. An example confusion matrix for the 3 month old data subset is shown in Table 6, and overall accuracies are given in Table 5. Chance accuracy in all cases is between 25−35%. The results suggest that vocalizations are affected by condition and also that vocal production patterns become more consistent over time. It can be seen that, as might be expected, the highest confusion is between conditions with overlapping stimuli, for example Heat vs. Heat + Human Presence vs. Heat + Crowding. Accuracy between the disjoint stimuli Heat, Human Presence, and Crowding is much higher.
Table 5.
Sample condition classification confusion matrix, for 3 month old age group. Accuracy 72.4% (vs. chance accuracy 25.3%).
| Classification | ||||||||
|---|---|---|---|---|---|---|---|---|
| Control | H | HP | H + HP | C | H + C | |||
| Label | Control | 97 | 0 | 2 | 0 | 0 | 0 | |
| H | 0 | 39 | 7 | 12 | 7 | 0 | ||
| HP | 5 | 1 | 70 | 0 | 20 | 0 | ||
| H + HP | 0 | 12 | 0 | 24 | 0 | 6 | ||
| C | 7 | 2 | 2 | 0 | 34 | 1 | ||
| H + C | 3 | 1 | 0 | 11 | 9 | 19 | ||
Table 6.
Overall 6 condition classification accuracy for 1, 2, and 3 month old age groups.
| Accuracy | |
|---|---|
| 1 month | 60.5% |
| 2 months | 62.0% |
| 3 months | 72.4% |
To examine the change in vocalization patterns as a function of age, a cross testing experiment is done by training and testing on disjoint age groups. An example of these results, training on one month old data and testing on two and three month old data, is shown in Table 7. Accuracy drops in proportion to age differences between training and test set conditions, dropping from 60.5% within the same age group (from Table 6), to 49.1% on 2 month olds (a one month age differential), to 34.9% on 3 months (a two month age differential).
Table 7.
Accuracy of 2 and 3 month old chick vocalizations using models trained on 1 month old data.
| Accuracy | |
|---|---|
| 2 months | 49.1% |
| 3 months | 34.9% |
3.3.2. Stress and Diet detection
A second study on poultry vocalizations is also implemented, focused on determining whether vocalization patterns could be used to differentiate dietary conditions (meat vs. vegetable fed), stress vs. nonstress conditions (human presence being the stress stimulus), and age of the animals (young, 0–4 weeks, vs. mature, 5−6 weeks).
Results for stress classification are shown in Table 8. All accuracies are above 90%, and indicate that within these conditions, human presence stress is relatively easy to detect. The slightly higher accuracy for mature vocalizations may be simply correlated with age, in that vocalization patterns tend to be more stable and established in older individuals, or may be due to learned reactions or preferences.
Table 8.
Stress (S) vs. Nonstress (NS) classification as a function of age and diet conditions.
| Meat fed 0–4 weeks old 98.6% | Classification | Veg fed 0–4 weeks old 94.40% | Classification | |||||
| NS | S | NS | S | |||||
| Label | NS | 176 | 0 | Label | NS | 224 | 1 | |
| S | 5 | 190 | S | 22 | 164 | |||
| Meat fed 5–6 weeks old 100% | Classification | Veg fed 5–6 weeks old 98.9% | Classification | |||||
| NS | S | NS | S | |||||
| Label | NS | 72 | 0 | Label | NS | 92 | 0 | |
| S | 0 | 89 | S | 2 | 82 | |||
Table 9.
Meat vs. Vegetarian diet classification as a function of age and stress conditions.
| Nonstress 0–4 weeks old 61.6% | Classification | Stress 0–4 weeks old 69.6% | Classification | |||||
| Meat | Veg | Meat | Veg | |||||
| Label | Meat | 157 | 140 | Label | Meat | 128 | 46 | |
| Veg | 24 | 106 | Veg | 62 | 119 | |||
| Nonstress 5–6 weeks old 89.2% | Classification | Stress 5–6 weeks old 78.4% | Classification | |||||
| Meat | Veg | Meat | Veg | |||||
| Label | Meat | 63 | 9 | Label | Meat | 71 | 19 | |
| Veg | 9 | 85 | Veg | 18 | 63 | |||
In comparison, determining whether or not diet has impact on vocalization patterns is a much more difficult classification task. Table 9 presents the results on this question. The main observation is that diet does impact vocalization patterns with accuracies high enough to confirm significance in all cases. Additionally, it can be seen that the nonstressed condition vocalizations are much easier to discriminate than the stress condition vocalizations, and that more mature animals are easier to differentiate than young animals. This indicates that diet-related vocalization differences are diminished by stress but increase with age. Maximum accuracy, for mature animals in the nonstressed condition, are relatively easy to discriminate on the basis of diet, at 89.2%. Looking comprehensively at all the poultry experiments, the main overall conclusion is that while vocalization patterns increase in consistency and differentiability with age, stress conditions can be differentiated across all age levels.
4. Conclusions
Overall, the experimental results show consistent performance across a diverse set of species and application tasks. The use of a robust automatic classification model can offer insight into repertoire variation, individual vocal variability, social context correlations, stress conditions, and many other important animal behavior questions. Hidden Markov Models, with a flexible recognition network topology and probabilistic framework for classification, coupled with generalized spectral features such as GFCCs for application across many species, offer an extremely robust and powerful framework for many bioacoustic applications.
Acknowledgements
This material is based on work supported by the National Science Foundation under Grant Numbers IIS-0326395 and OIA-0528598 and by the Polish State Committee for Scientific Research Grant No. 3-P04C-083-25. The authors would also like to acknowledge Portland State University, the Oregon Zoo, the OSU/NOAA Cooperative Institute for Marine Resource Studies, Royal Elephant Kraal, and the Elephant Nature Park.
References and Notes
- Clark, C.W.; Marler, P.; Beeman, K. Quantitative analysis of animal vocal phonology: An application to swamp sparrow song. Ethology 1987, 76, 101–115. [Google Scholar] [CrossRef]
- Chabot, D. A quantitative technique to compare and classify humpback whale (Megaptera novaeangliae) sounds. Ethology 1988, 77, 89–102. [Google Scholar] [CrossRef]
- Mellinger, D.K.; Clark, C.W. Recognizing transient low-frequency whale sounds by spectrogram correlation. In Proceedings of the IEEE Oceans '93, Victoria, BC, Canada, October 1993.
- Stafford, K.M.; Fox, C.G.; Clark, D.S. Long-range acoustic detection and localization of blue whale calls in the northeast Pacific Ocean. J. Acoust. Soc. Am. 1998, 104, 3616–3625. [Google Scholar] [CrossRef] [PubMed]
- Ramani, N.; Patrick, P.H. Fish detection and identification using neural networks. IEEE J. Ocean Eng. 1992, 17, 364–368. [Google Scholar] [CrossRef]
- Gaetz, W.; Jantzen, K.; Weinberg, H.; Spong, P.; Symonds, H. A neural network mechanism for recognition of indivudual Orcinus orca based on their acoustic behavior: Phase 1. In Proceedings of the IEEE Oceans '93, Victoria, BC, Canada, October 1993.
- Moore, P.W.B.; PRoitblat, H.L.; Penner, R.H.; Nachtigall, P.E. Recognizing successive dolphin echoes with an integrator gateway network. Neural Netw. 1991, 4, 701–709. [Google Scholar] [CrossRef]
- Buck, J.R.; Tyack, P.L. A quantitative measure of similarity for tursiops truncatus signature whistles. J. Acoust. Soc. Am. 1993, 94, 2497–2506. [Google Scholar] [CrossRef] [PubMed]
- Clemins, P.J.; Johnson, M.T.; Leong, C.M.; Savage, A. Automatic classification and speaker idenfication of African elephant (Loxodonta africana) vocalizations. J. Acoust. Soc. Am. 2005, 117, 1–8. [Google Scholar] [CrossRef]
- Clemins, P.J.; Trawicki, M.B.; Adi, K.; Tao, J.; Johnson, M.T. Generalized perceptual features for vocalization and analysis across multiple species. In Proceedings of the IEEE International Conference on Acoustic, Speech and Signal Processing, Toulouse, France, May 2006.
- Adi, K.; Sonstrom, K.E.; Scheifele, P.M.; Johnson, M.T. Unsupervised validity measures for vocalization clustering. In Proceedings of the Internatinal Conference on Acoustics Speech and Signal Processing, Las Vegas, NV, USA, April 2008.
- Tao, J.; Johnson, M.T.; Osiejuk, T. Acoustic model adaptation for ortolan bunting (Emberiza Hortulana L.) song type classification. J. Acoust. Soc. Am. 2008, 123, 1582–1590. [Google Scholar] [CrossRef] [PubMed]
- Weisburn, B.A.; Mitchell, S.G.; Clark, C.W.; Parks, T.W. Isolating biological acoustic transient signals. In Proceedings of the 1993 International Conference on Acoustics, Speech, and Signal Processing, Minneapolis, MN, USA, April 1993.
- Anderson, S.E. Speech recognition meets bird song: A comparison of statistics-based and template-based techniques. J. Acoust. Soc. Am. 1999, 106, 2130. [Google Scholar] [CrossRef]
- Murray, S.O.; Mercado, E.; Roitblat, H.L. The neural network classification of false killer whale (Pseudorca crassidents) vocalizations. J. Acoust. Soc. Am. 1998, 104, 3626–3633. [Google Scholar] [CrossRef] [PubMed]
- Mellinger, D.K. Ishmael 1.0 User's Guide; Pacific Marine Enviromental Laboratory: Seattle, WA, USA, 2002. [Google Scholar]
- Liu, R.C.; Miller, K.D.; Merzenich, M.N.; Schreiner, C.E. Acoustic variability and distinguishability among mouse ultrasound vocalizations. J. Acoust. Soc. Am. 2003, 114, 3412–3422. [Google Scholar] [CrossRef] [PubMed]
- Ephraim, Y.; Malah, D. Speech Enhancement using a minimum mean-square error log-spectral amplitude estimator. IEEE Trans Acoust 1985, ASSP–33, 443–445. [Google Scholar] [CrossRef] [PubMed]
- Alvarez, B.D.; Garcia, C.F. System architecture for pattern recognition in eco systems. In Proceedings of the ESA-EUSC, Madrid, Spain, September 2004.
- Yan, Z.; Niezrecki, C.; Cattafesta, L.N.I.; Beusse, O.D. Background noise cancellation of manatee vocalizations using an adaptive line enhancer. J. Acoust. Soc. Am. 2006, 120, 145–152. [Google Scholar] [CrossRef] [PubMed]
- Ren, Y.; Johnson, M.T.; Tao, J. Perceptually motivated wavelet packet transform for bioacoustic signal enhancement. J. Acoust. Soc. Am. 2008, 124, 316–327. [Google Scholar] [CrossRef] [PubMed]
- Deller, J.R., Jr; Hansen, J.H.L.; Proakis, J.G. Discrete-Time Processing of Speech Signals; IEEE: Piscataway, NJ, USA, 2000. [Google Scholar]
- Department, C.U.E. Hidden Markov Model Toolkit (HTK) Version 3.2.1 User's Guide; Cambridge: Cambridge, MA, USA, 2002. [Google Scholar]
- Baum, L.E.; Petrie, T.; Soules, G.; Weiss, N. A maximization technique occurring in the statistical analysis of probabilistic functions of Markov chains. Ann. Math. Stat. 1970, 41, 164–171. [Google Scholar] [CrossRef]
- Baum, L.E. An inequality and associated maximization technique in statistical estimation for probabilistic functions of Markov processes. Inequalities 1972, 3, 1–8. [Google Scholar]
- Forney, G.D. The viterbi algorithm. IEEE 1973, 61, 268–278. [Google Scholar] [CrossRef]
- Davis, S.B.; Mermelstein, P. Comparison of parametric representations for monosyllabic word recognition in continuously spoken sentences. IEEE Trans. Acoust. 1980, 28, 357–366. [Google Scholar] [CrossRef]
- Hermansky, H. Perceptual Linear predictive (PLP) analysis for speech recognition. J. Acoust. Soc. Am. 1990, 87, 1738–1752. [Google Scholar] [CrossRef] [PubMed]
- Clemins, P.J. Automatic Speaker Identification and Classification of Animal Vocalizations. Dissertation, Electrical and Computer Engineering Department, Marquette University, Milwaukee, WI, USA, May 2005. [Google Scholar]
- Greenwood, D.D. Critical bandwidth and the frequency coordinates of the basilar membrane. J. Acoust. Soc. Am. 1961, 33, 1344–1356. [Google Scholar] [CrossRef]
- Trawicki, M.B.; Johnson, M.T. Automatic song-type classification and speaker identification of Norwegian Ortolan Bunting (Emberiza Hurtulana). In Proceedings of the IEEE International Conference on Machine Learning in Signal Processing (MLSP), Mystic, CT, USA; 2005. [Google Scholar]
- Langbauer, W.R.J. Elephant communication. Zoo Biol. 2000, 19, 425–455. [Google Scholar] [CrossRef]
- Eisenberg, J.F.; McKay, G.M.; Jainudeen, M.R. Reproductive behavior of the Asiatic elephant (Elephas maximus). Behav. Ecol. Sociobiol. 1971, 38, 193–225. [Google Scholar]
- Douglas-Hamilton, I. On the ecology and behaviour of the Lake Manyara elephants. E Afr. Wildl. J. 1973, 11, 401–403. [Google Scholar] [CrossRef]
- Rasmussen, L.E.L.; Gunawardena, R.A.; Rasmussen, R.A. Do Asian elephants, especially males in musth, chemically signal via volatiles in breath? Chem. Sens. 1997, 22, 775. [Google Scholar]
- Schulte, B.; Rasmussen, L.E.L. Signal-receiver interplay in the communication of male condition by Asian elephants. Anim. Behav. 1999, 57, 1265–1274. [Google Scholar] [CrossRef] [PubMed]
- Poole, J.H. Signal and Assessment in African Elephants: Evidence from playback experiments. Anim. Behav. 1999, 58, 185–193. [Google Scholar] [CrossRef] [PubMed]
- McKay, G.M. Behavior and ecology of the Asiatic elephant in Southeastern Ceylon. Smithsonian Contrib. Zool. 1973, 125, 1–113. [Google Scholar] [CrossRef]
- Payne, K.B.; Langbauer, J.W.R.; Thomas, E.M. Infrasonic calls of the Asian elephant. Behav. Ecol. Sociobiol. 1986, 18, 297–301. [Google Scholar] [CrossRef]
- Glaeser, S.S.; Klinck, H.; Mellinger, D.K.; Ren, Y.; Clemins, P.J.; Johnson, M.T.; Cook, M.L.H.; Zelick, R. A vocal repertoire of Asian elephant and comparison of call classification methods. J. Acoust. Soc. Am. 2009, 125, 2710. [Google Scholar] [CrossRef]
- Glaeser, S.S. Analysis and Classification of Sounds Produced by Asian Elephants (Elephas Maximus). MSc Thesis, Department of Biology, Portland State University, Portland, OR, USA, 2009. [Google Scholar]
- Cramp, S.; Perrins, C.M. The birds of the western palearctic; Oxford University Press: Oxford, UK, 1994. [Google Scholar]
- Steinberg, B.D. Microwave Imaging with Large Antenna Arrays; John Wiley & Sons: New York, NY, USA, 1983. [Google Scholar]
- Dale, S. Causes of Population Decline in Ortolan Bunting in Norway. In Proceedings of the 3rd International Ortolan Symposium, Poznan, Poland; 2001. [Google Scholar]
- Losak, K. A Comparative Analysis of Song Variation in Ortolan Bunting (Emberiza Hortulana) from Populations of Different Status and Quality. Dissertation, Adam Mickiewicz University, Poznan, Poland, 2007. [Google Scholar]
- Osiejuk, T.S.; Ratynska, K.; Cygan, J.P.; Svein, D. Song structure and repertoire variation in ortolan bunting (Emberiza hortulana L.) from isolated Norwegian population. Ann Zool Fenn 2003, 40, 3–19. [Google Scholar]
- Michael, C.A.; Joy, A.M.; Hughes, B.O. Poultry Behaviour and Welfare; CABI: Cambridge, MA, USA, 2004. [Google Scholar]
© 2009 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).