1. Introduction
The loss of biodiversity continues on both regional and global scales across a wide range of ecosystems, due to the increasing intensity of disturbances and despite conservation efforts [
1]. Windstorms, wildfires, floods, drought, and insect epidemics are the main natural disturbances in forests [
2], creating so-called “early-successional forest ecosystems” (ESFEs) [
3]. The relevance of these disturbances has tended to increase in recent decades because of their increasing occurrence, severity, frequency, and intensity worldwide associated with changes in climate, which are becoming increasingly obvious [
4].
Successional changes begin immediately after a disturbance, due to the activities of the surviving organisms (e.g., animals, plants, and fungi), including plant growth and seed production. Initial conditions, however, generally vary, depending on the type of disturbance. Forest sites after severe disturbances are characterized by open environments not dominated by trees, which dramatically alter the site microclimate, including light conditions [
3]. These changes lead to increased exposure to sunlight, more extreme temperatures (ground and air), higher air velocities, and lower levels of relative humidity and moisture in the litter and surface soil. Shifts in these environmental metrics favor some species but create suboptimal or intolerable conditions for others [
3].
Windstorms in Europe were responsible for 53% of the total damage in forests from 1950–2000 [
5]. Most European forests are intensively managed, so windstorm damage decreases the yield of recoverable timber, increases the cost of unscheduled cuttings, creates problems in forestry planning, and releases considerable amounts of carbon (C), as recently demonstrated [
1,
6]. Ecological studies after such disturbances have therefore focused primarily on plant-community development, tree regeneration, and the re-establishment of closed forest canopies [
7,
8,
9]. Windstorms, however, have also destroyed forests in national parks and reserves across Europe that were developed to conserve the biodiversity of rare habitats, e.g., primeval forests or wetlands, and to encourage tourism and economic and social development in rural areas [
5,
10,
11,
12,
13]. Two strong windstorms in 2004 and 2014 destroyed thousands of hectares of spruce forest in the Tatra National Park, Slovakia, both leading to extensive outbreaks of bark beetles [
14,
15]. Extensive research in ESFEs with various management strategies has identified considerable changes to the microclimate, hydrology, stream morphology, slope erosion, plant communities, soil properties, and epigeic and edaphic macrofauna [
9,
16,
17,
18,
19] including the characteristics of soil microbial and nematode communities [
20,
21]. The soil microbial and nematode communities at a site vary with the tree species [
22,
23,
24], with the largest differences being between broadleaved and coniferous species [
22,
25,
26]. Information on the general response of these closely related groups of soil biota to such disturbances in a managed deciduous forest is nevertheless lacking, and the successional patterns and relationships of these groups of soil microfauna after a disturbance have not recently been surveyed.
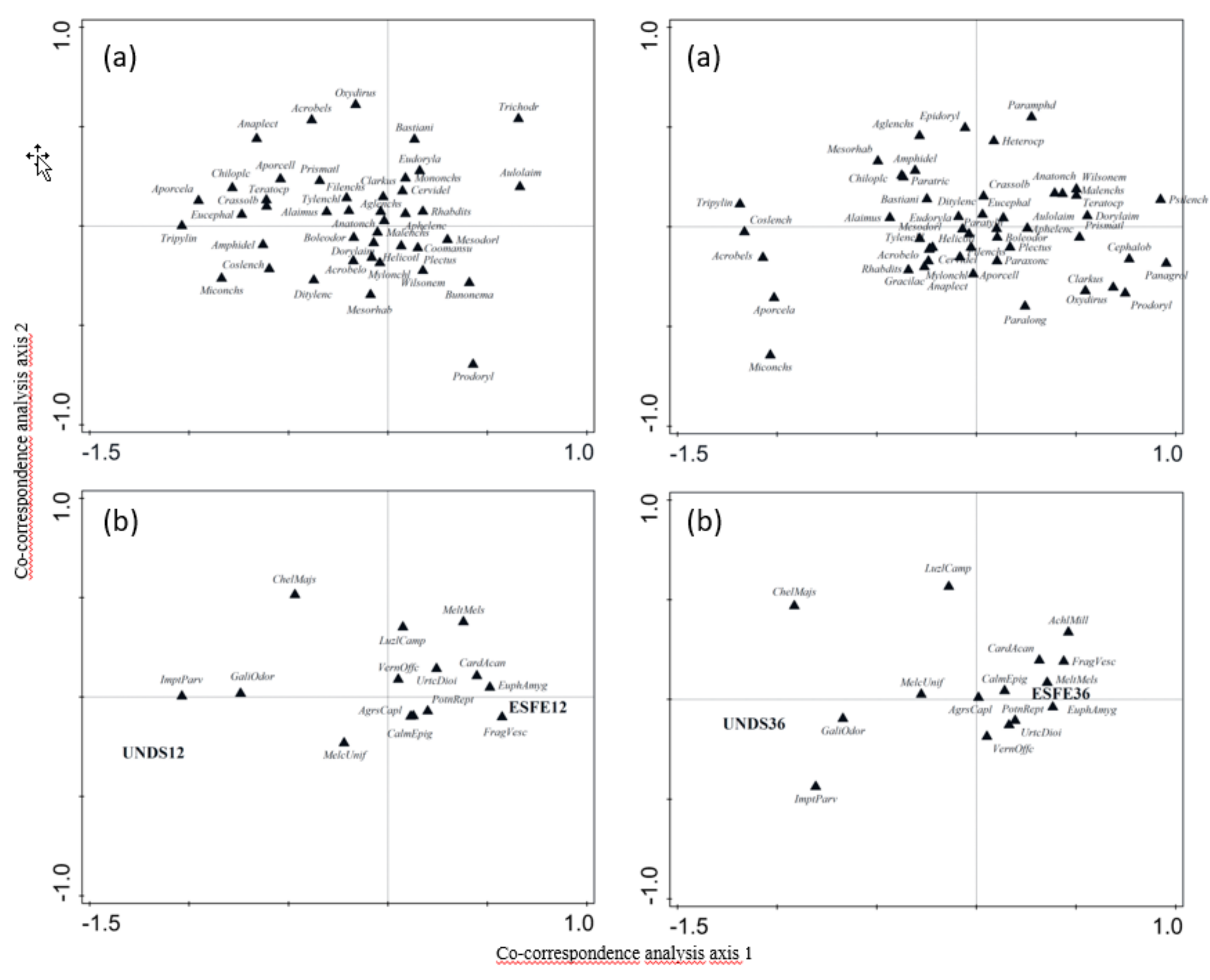
We studied the nematode communities and microbial characteristics at disturbed and undisturbed sites in deciduous beech forests 12 and 36 months after a windstorm to evaluate (1) whether soil nematode communities and microbial activity were affected by changes in the aboveground ecosystem, (2) relationships between biotic and abiotic soil features, (3) possible mutual links between specific nematode genera and plant species associated with secondary succession of understory vegetation, and (4) selection amongst various parameters that could serve as indicators of changes in soil state throughout forest development.
Our previous study of acidic soils at windstorm-disturbed sites in natural montane
Lariceto-Piceetum forests of the High Tatra National Park provided evidence that the windstorm strongly negatively influenced the activity and functional diversity of microbial communities [
16], but had little effect on the community structure, diversity, and abundance of nematodes [
21]. We tested the hypothesis that nematode communities and microbial activity would have the same pattern of change because the kind of natural disturbance was the same (windstorm), except for the neutral soils of the devastated deciduous productive
Fagus sylvatica L. forests.
4. Discussion
Change is a characteristic of all natural systems, usually beginning with disturbances to the existing community. Disturbances in forests such as windstorms, wildfires, or insect epidemics create open environments not dominated by trees, commonly known as early successional ecosystems. Areas damaged by windstorms are usually cleared; disturbed stands are harvested for economic (recouping financial losses before wood quality deteriorates) and sanitary (damaged trees can attract insects that can attack undisturbed trees in their neighborhood) reasons. Removal of the forest canopy dramatically alters the site microclimate, mainly light and water conditions [
3], that lead to changes in the composition, diversity, and habitat structure of understory plant species, which have potential implications for the soil biota [
48]. Any disturbance of the soil will lead to a succession in bacteria and fungi and the associated food web, with an initial decrease and then an increase in biodiversity [
50], in agreement with the authors of Reference [
16] who found that the characteristics of microbial activity (basal respiration, microbial biomass, N mineralization, and catalase activity) were significantly lower in the acidic soil of a cleared site of a
Lariceto-Piceetum coniferous forest immediately after a windstorm. In contrast, total nematode abundance, species diversity, and number of species were not altered soon after [
21] and nine years after the windstorm [
51]. The abundance of some trophic groups (bacterivores and herbivores) even slightly increased at cleared sites [
21]. We thus tested the hypothesis that the responses of soil nematode communities and microbial activity to windstorm disturbance in the neutral soil of a deciduous
F. sylvatica forest and secondary succession would have the same pattern as detected in the acidic soil of a coniferous
Lariceto-Piceetum forest.
Secondary succession of the understory vegetation was visible at the ESFE sites 12 months after the windstorm, with progression over time, as expected. In contrast, plant composition did not change at the UNDS sites between the two sampling dates. Some herbs such as
G. odoratum,
C. majus, and
I. parviflora were suppressed at the ESFE sites, and two heliophilous grasses,
C. epigejos and
A. capillaris, benefited from the increase in light exposure caused by the removal of fallen trees. Communities dominated by
C. epigejos or
A. capillaris, which develop at forest sites after tree cutting or windstorm clearance, are common and widely distributed in European beech forests [
8,
52]. Sites disturbed by windstorms [
9] were rapidly colonized by the heliophilous species
Calamagrostis villosa and
Avenella flexuosa 18 months after the devastation of spruce forests in High Tatra National Park, and the abundance of the nitrophilous herb
Chamaerion angustifolium increased considerably one year later due to high N inputs. This vegetation succession was attributed to changes in light and moisture conditions, and the high N content was attributed to the absence of uptake by vegetation [
9].
C. villosa and
A. flexuosa also remained dominant at the disturbed sites nine years after the windstorm, but
C. angustifolium disappeared due to the succession of the forest floor [
51]. The succession of the understory plant species was positively correlated with the C and N contents at the ESFE sites, mainly for grasses.
F. sylvatica, however, has a very high competitive capacity against other tree species, and the understory vegetation is very species poor [
53], in agreement with our findings and those reported by [
8]. Such conditions, however, are favorable to soil nematodes, indicated by our records at the UNDS sites (relatively high abundance and high total biomass and number of nematode species), supporting the results of several previous studies of European deciduous forests [
54,
55,
56]. The overall changes at the ESFE sites caused by the windstorm therefore negatively altered the naturally rich nematode communities, i.e., nematode abundance of most species decreased, mainly the most abundant species and those from higher c–p groups (3–5). These changes led to a decrease in total nematode abundance and biomass, despite ongoing understory secondary succession 12 months after the storm (ESFE12). A PCA found a slight restoration of the nematode communities 24 months later (ESFE36).
Undisturbed sites in a montane
Lariceto-Piceetum forest with acidic soils [
51] naturally had ten-fold lower nematode biomass, five-fold fewer nematode individuals, and half the number of nematode species as the neutral soils of our
F. sylvatica forests, supporting the findings by [
57], who reported total numbers of nematodes about two- to three-fold higher at higher than lower pHs. The disturbance of the ecosystem by the windstorm thus did not have a detrimental impact on the nematode communities and even positively affected nematode abundance, attributed to an improvement of microclimatic conditions, mainly due to secondary plant succession and changes in the herbaceous cover, which can directly or indirectly affect food sources for different nematode trophic groups [
21,
51]. This contradiction may have been due to the identity of the tree species, differing in physiology, leaf-litter quality, or type of mycorrhiza, as a natural factor affecting the overall ecosystem environment and thus faunal biodiversity [
58] including soil nematodes [
23,
59] For example, native deciduous forests (mainly beech) have been replaced by coniferous forests (mainly spruce) in most parts of Europe, a common consequence of former European afforestation policies [
53]. Such conversions, however, have been problematic for ecosystem health by acidifying the soil [
58] stimulating pest abundance [
60] but also strongly negatively affecting the biomass of all nematode trophic groups, but also enchytraeids, earthworms, isopods, and mycetophilid or cecidomyiid Diptera [
58]. The re-establishment of pure coniferous and mixed forests, however, has had a positive impact on the soil food web, including nematode communities [
61,
62,
63].
The composition of the nematode trophic groups in our study was typical for deciduous-forest ecosystems, supporting the results of several previous studies of deciduous forests in Europe [
23,
56,
64]. The forest disturbance, however, significantly negatively affected the abundance of most nematode functional guilds in the trophic groups. None of the functional guilds, however, disappeared at the ESFE sites. The effect of the windstorm was thus quantitative rather than qualitative, partially supporting our findings from a
Lariceto-Piceetum forest [
21]. Bacterivore abundance decreased in the ESFEs, supporting the results by [
25,
65] who reported higher abundances of bacterivores in healthy than disturbed forests, but contradicting the results in Reference [
21]. Similarly, fungivores, plant parasites, and omnivores were less abundant at the disturbed sites of the
F. sylvatica forests in the present study. In contrast, disturbed sites in the
Lariceto-Piceetum forest had higher abundances of plant parasitic nematodes but similar abundances of fungivores and omnivores than at undisturbed sites [
21]. These contradictory results may have been due to the overall characteristics of the ecosystems, such as identity of the tree species, climatic conditions, soil physicochemical properties, and the diversity and composition of the plant species in the secondary succession [
25,
58].
We expected a positive relationship between SM content and nematode abundance, because nematodes depend on the film of water surrounding soil particles for movement and to prevent desiccation [
66]. Total nematode abundance in our study, however, was not correlated with SM content, supporting our previous records from spruce forest [
51], indicating a more likely important role of secondary succession. Several nematode functional guilds were nevertheless positively correlated with SM content (Pp
2,3,4, Fu
2 and Ba
2) and soil pH (Ca
3,4). These findings agree with those by [
67]. It was found, that soil organic C and total N contents were relatively important factors influencing soil nematode communities in forest habitats, partially supporting our findings, especially for bacterivorous and fungivorous guilds, those negatively correlated with C and N contents [
68]. EI, SI, CI, and BI (except CI in ESFE36) could not distinguish between disturbed and undisturbed ESFE sites, contradicting the findings from the
Lariceto-Piceetum forest [
21]. These results partially agree with those by [
25] who found that forest disturbance did not affect CI or SI but that EI was significantly higher in the soil of a disturbed forest.
Soil microorganisms tend to respond quickly to environmental stress, because they have a higher surface-to-volume ratio than higher organisms, so the exchange of matter and energy with their environment is much more intense [
69]. The size, composition, and activity of microbial communities can frequently change before changes in soil physical and chemical properties become detectable, supporting the results found in Reference [
16] for soils at disturbed sites of the
Lariceto-Piceetum forest shortly after a windstorm. The large impact of the windstorm on soil microbial activity, biomass, and community composition in our study, however, was not apparent 12 months after the storm, in agreement with the results in Reference [
70] who reported no differences in fungal or bacterial biomass after the removal of forest residues from harvesting cleared sites.
Basal respiration, N mineralization, and microbial biomass, however, had decreased significantly in the ESFEs 36 months after the storm, supporting the findings in Reference [
71] but contradicting the statement by the authors of Reference [
72] that changes in microbial activity due to environmental changes can be independent of possible changes in microbial biomass. Soil pH may influence the biomass of soil microorganisms but not their activity [
20,
57], in agreement with our finding that soil pH was positively correlated with microbial biomass but not with microbial activity. Surprisingly, the functional diversity of the microbial community was negatively correlated with SM content, supporting the results in Reference [
20] but was positively correlated with C and N contents or the C/N ratio, contradicting the findings in Reference [
20].
5. Conclusions
Communities of microorganisms, especially those of forest soils with well-developed litter and humus layers, are characterized by a large diversity of species. Such communities are particularly sensitive to disturbance, because the long lifespan of the trees does not allow rapid adaptation to environmental changes. Our study found considerably different responses of soil nematode communities and microbial activity to the same type of disturbance in productive deciduous forests than in our previous studies in a natural coniferous forest shortly after a windstorm.
The structures of the soil nematode communities were significantly negatively altered by the windstorm in the beech forests, and the total abundance and numbers of plant parasites, bacterivores, and omnivores increased after a similar disturbance in spruce forest. In contrast, changes in microbial activity and functional diversity of the microbial communities appeared long after the disturbance in the beech forests but immediately after the disturbance in spruce forest.
These differences could be attributed to a set of environmental factors determining the natural degree of species diversity or abundance. For example, the identity of the tree species, differing in physiology, leaf-litter quality, or type of mycorrhiza may alter the physicochemical environment inhabited by nematodes and microbes. Climatic conditions or altitude differentiates the growth of tree species, forest types, and early-successional plant communities that have been injured by windstorms. Spruce forests under natural conditions are generally restricted to upper montane regions and wet acidic habitats, but natural beech forests grow optimally at lower altitudes with a wide range of soil physicochemical conditions and types.
The duration of our investigation was too short to expect significant changes in soil chemical properties between sites, but we nevertheless recorded significant increases in the C and N contents and a higher C/N ratio in the ESFEs in the third year after the storm. We conclude that the nematodes of the neutral beech forest soils are more sensitive to initial environmental changes from harvesting timber after a disturbance than nematodes from acidic spruce soils, contradicting the responses of microbial activity to such disturbances in these different forest types.






{kind=link}
{kind=link}
{kind=link}
{kind=link}