Facilitating Adaptive Forest Management under Climate Change: A Spatially Specific Synthesis of 125 Species for Habitat Changes and Assisted Migration over the Eastern United States

Abstract
:1. Introduction
2. Material and Methods
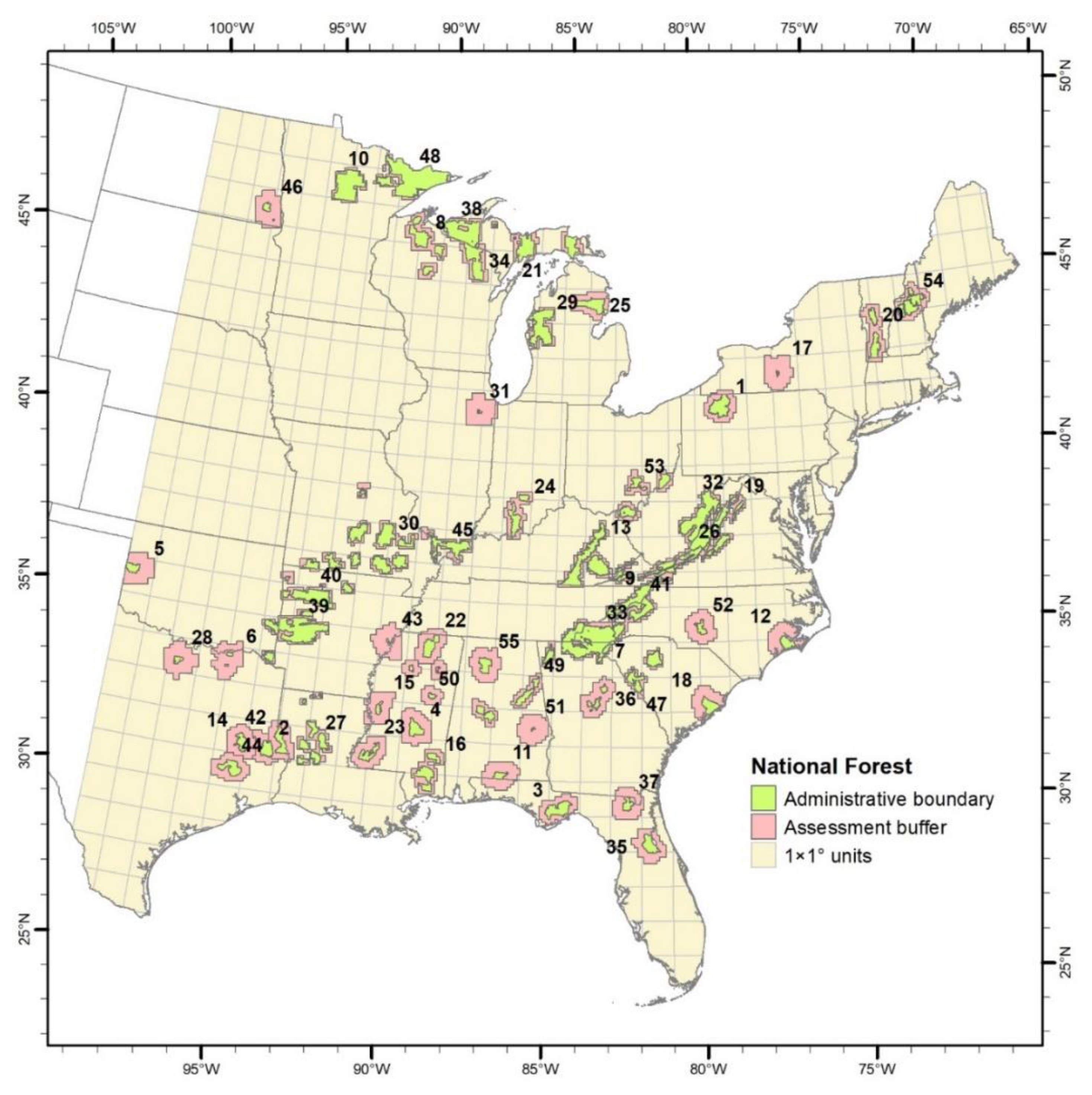
2.1. Study Area
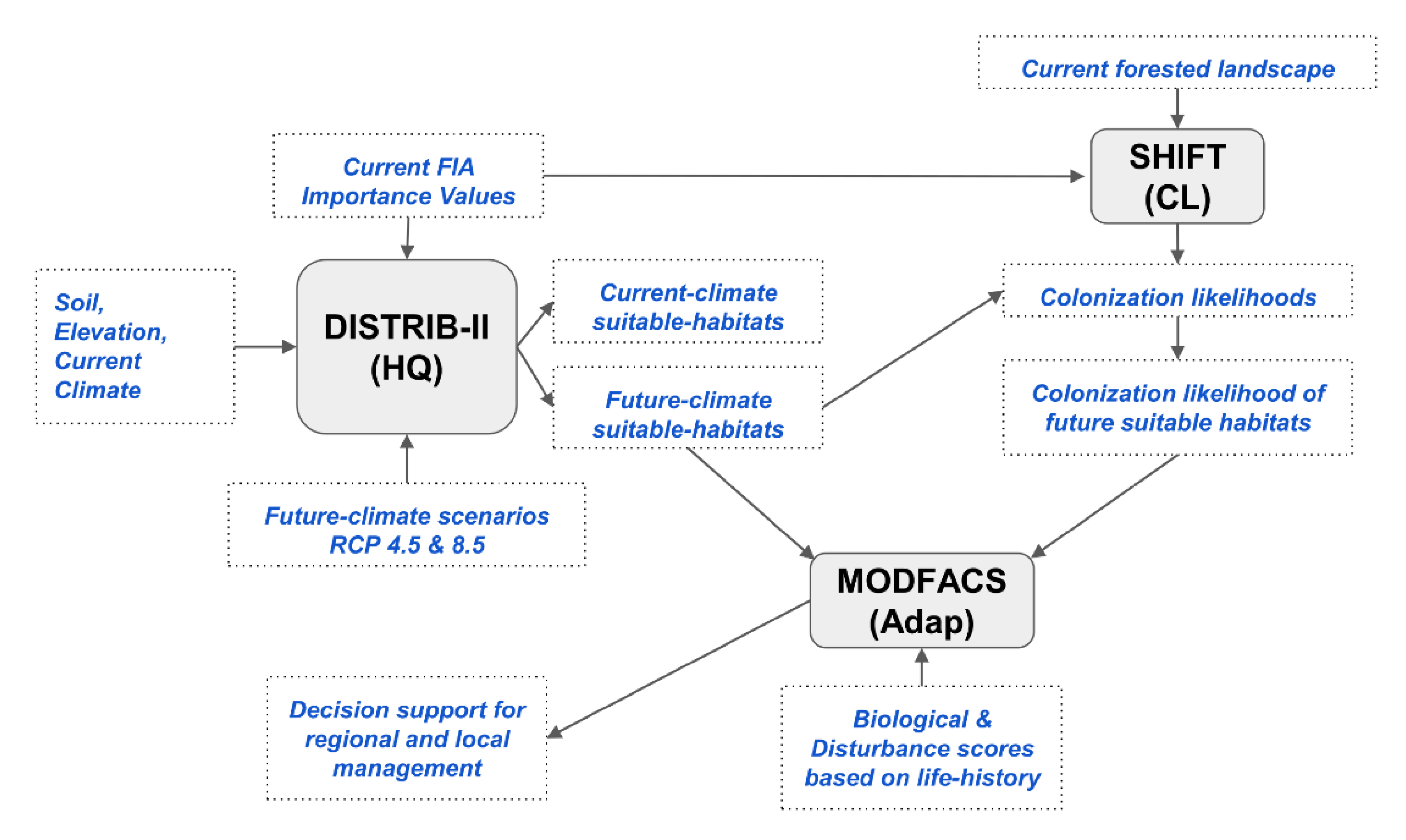
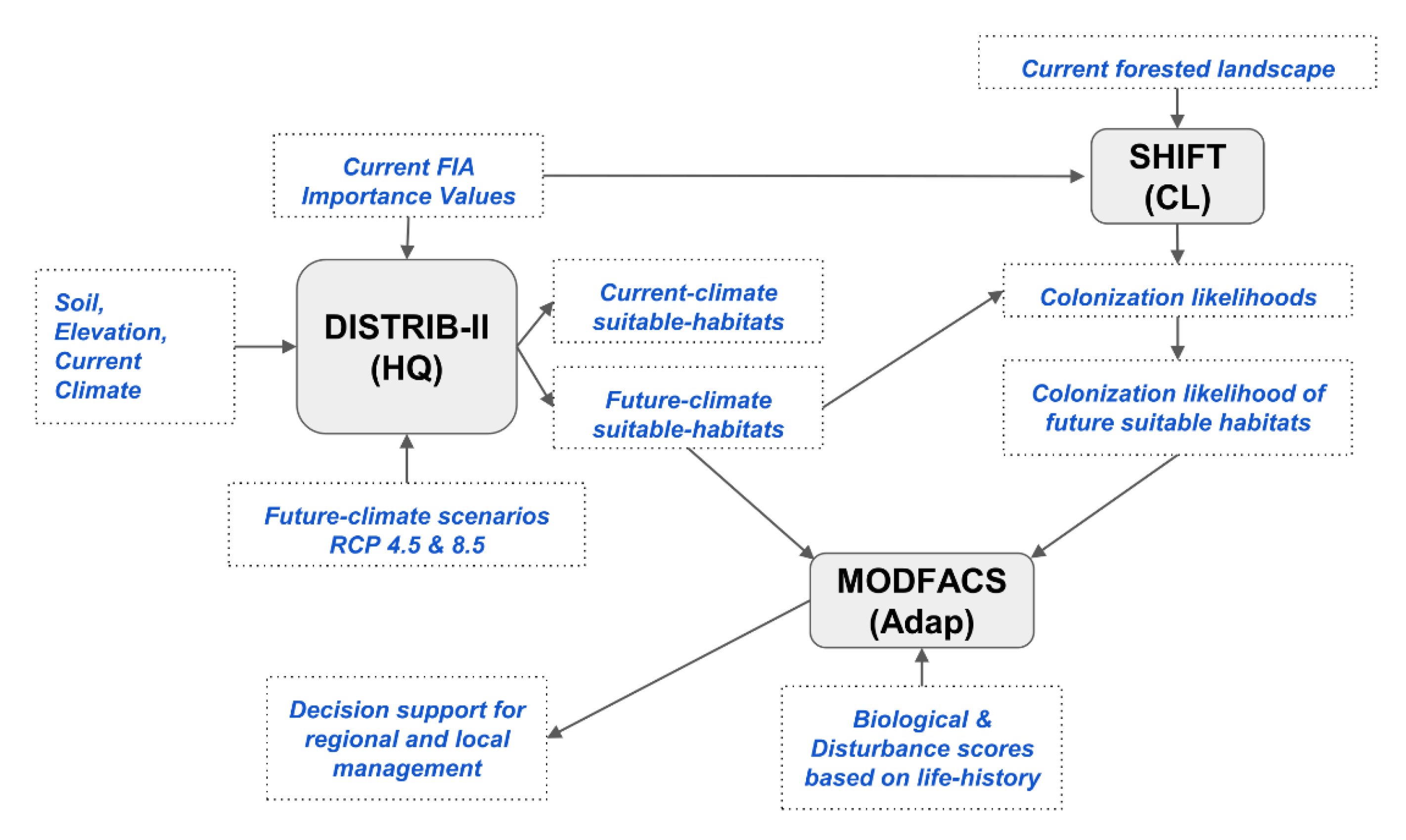
2.2. The Synthetic Approach
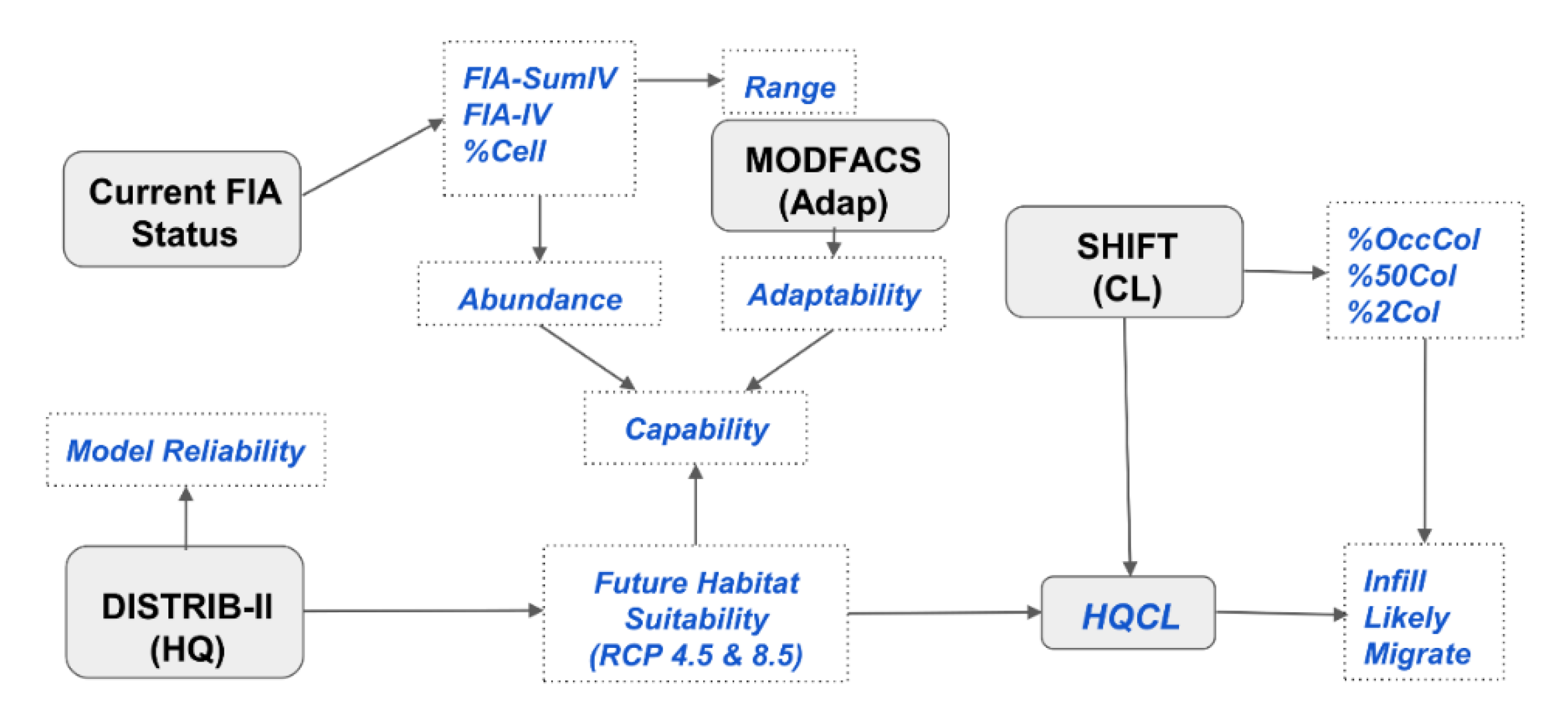
2.3. Rules for Capability to Cope with a Changing Climate
2.4. Rules for Expand, Migrate, and Likely, and Species Selection Options
2.5. Mapping Aggregated 1 × 1 Outputs
3. Results
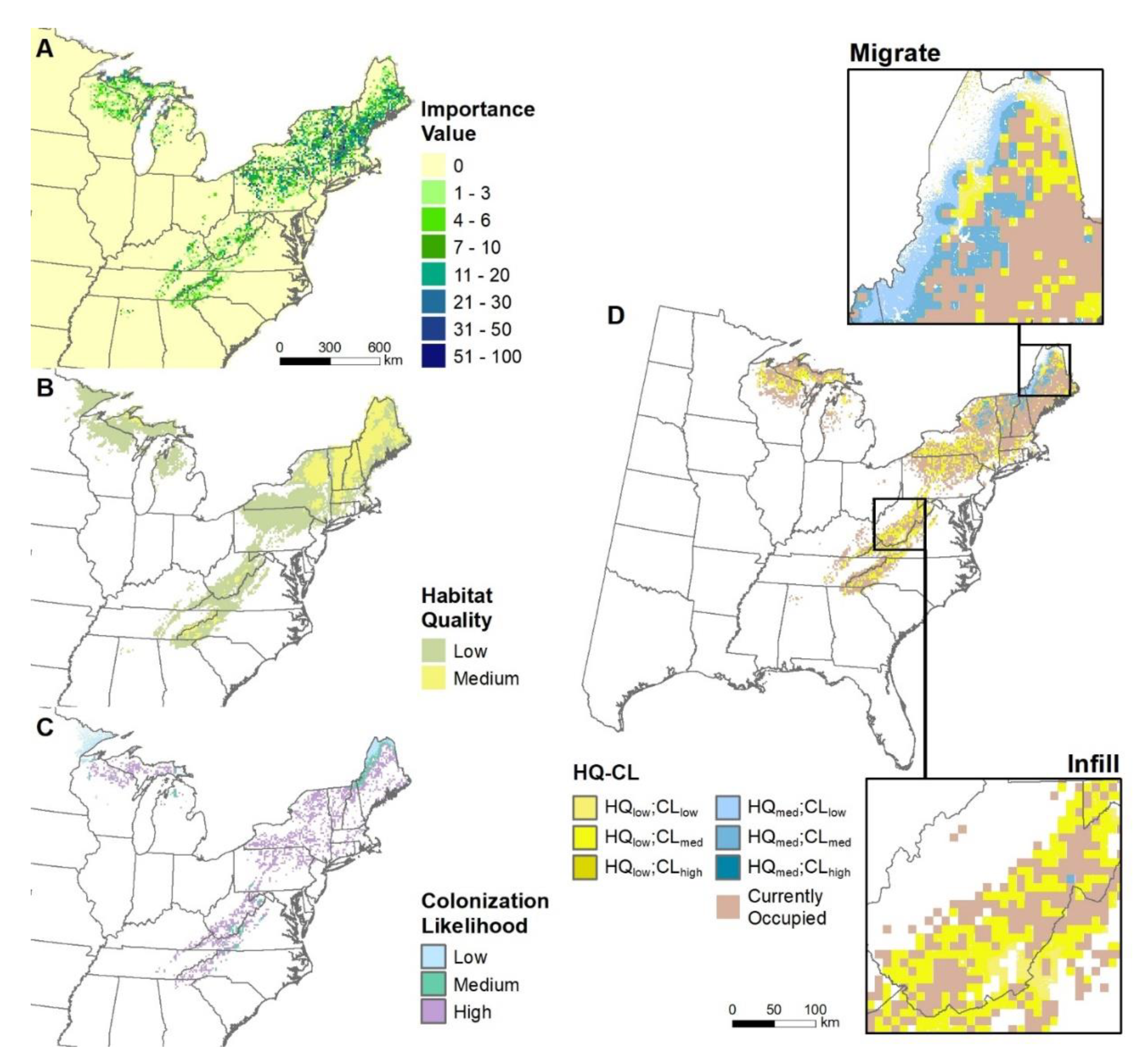
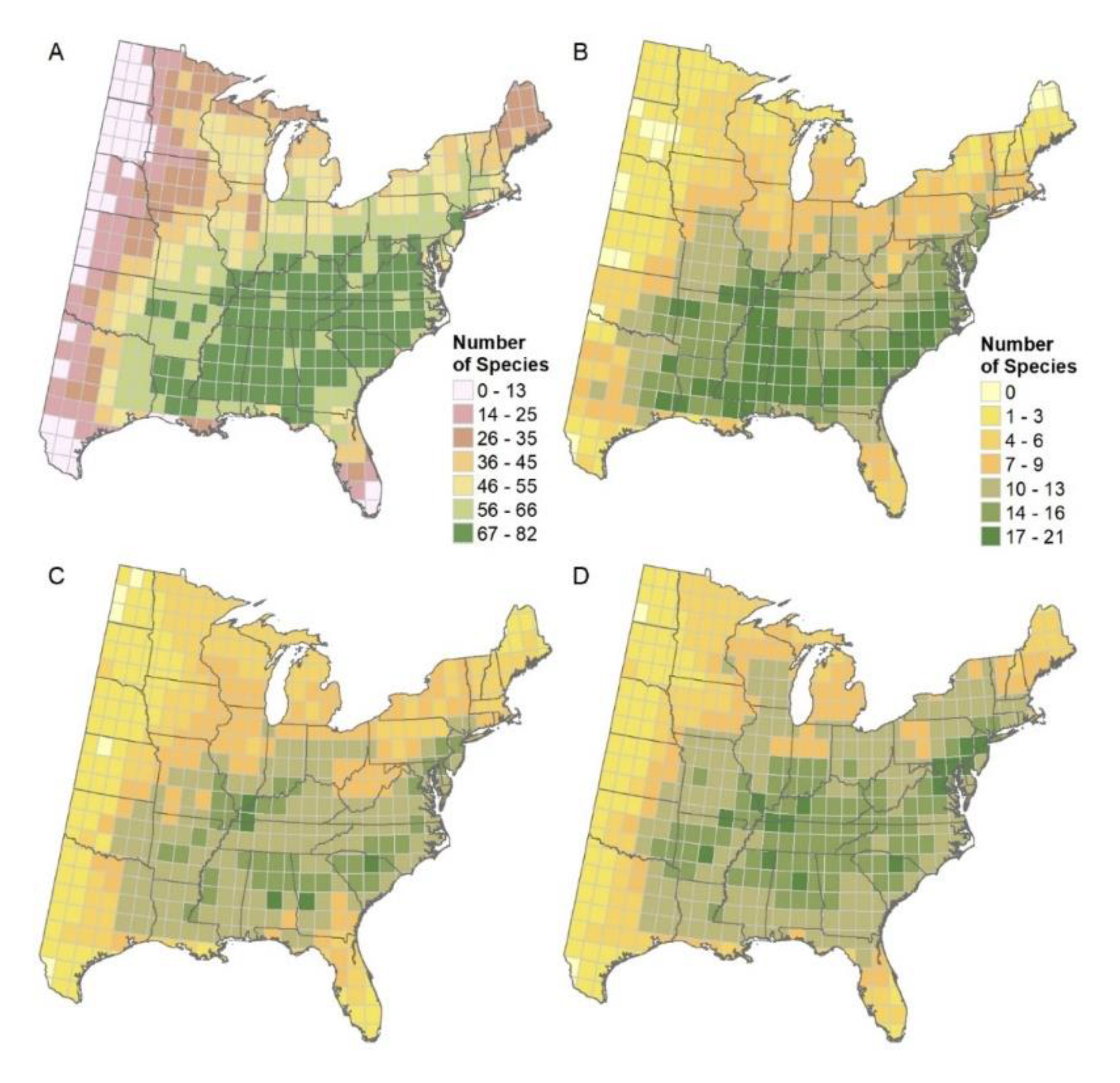
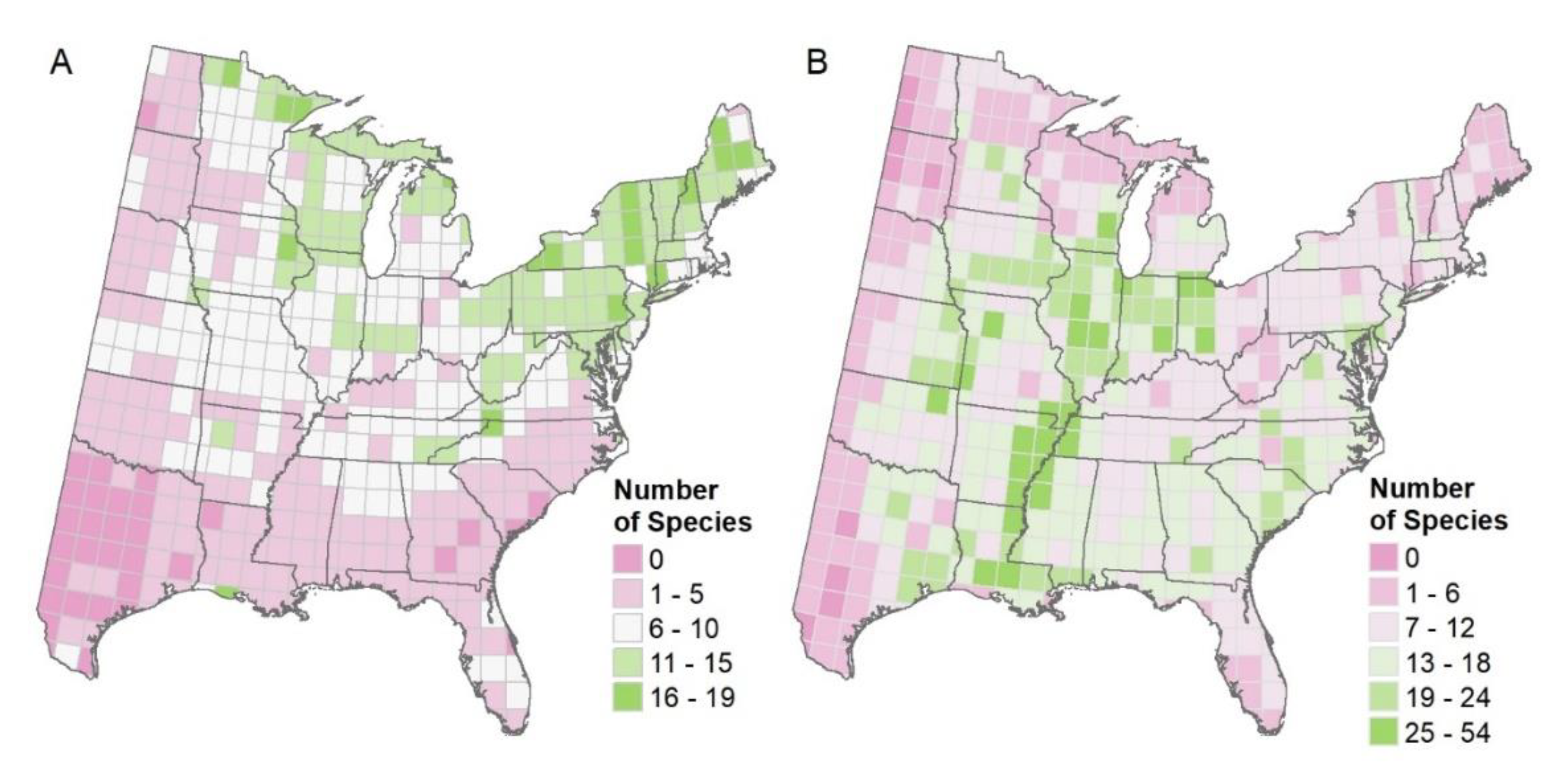
3.1. Mapping Habitat Quality and Colonization Likelihood Outputs
3.2. Species Summaries by National Forest
3.3. Trends in Area and Species Counts for National Forests and Grasslands
3.4. Questions and Answers for Example NF: Allegheny NF
- How many species are known (by FIA) to be present currently, what are these species, and how abundant are they?
- By 2100, how might suitable habitat change for trees under low and high emission scenarios?
- How adaptable are the tree species to conditions expected to change under climate change?
- How capable might the species be with regards to coping with the changing climate, and what species may be most (or least) vulnerable?
- Within the next 100 years, what species may be able to naturally migrate into the area?
- What species, now rare, are suited to expand or infill in importance over the century?
- What species are likely present in the area, though missed by FIA plots?
- What species might be useful for selecting as candidates for planting in the face of a changing climate?
3.5. Mapped Summaries for 1 × 1 Degree Grid
4. Discussion
4.1. Applications for Summary Tables
4.2. Mapped Summaries for 1 × 1 Degree Grids
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A
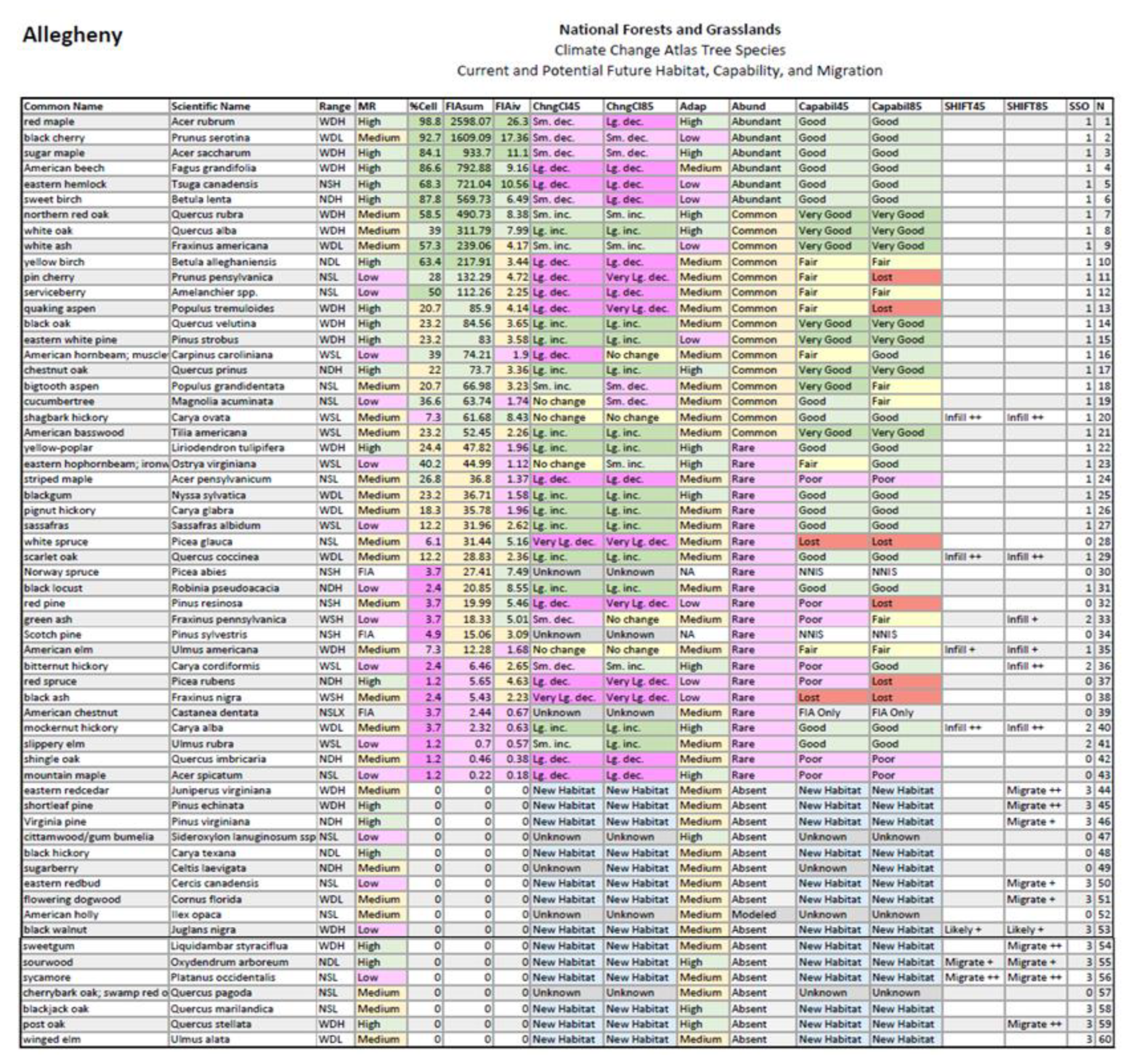
Appendix A.1. Questions and Answers for Example NF: Allegheny NF
Appendix A.1.1. Current Status of Species
- Of the species modeled, how many are known (by FIA) to be present in this NF?
- The column N counts the species in this table. Some are present and some are modeled to have suitable habitat appear by 2100. Those with %Cell > 0 are known by FIA to be present. Some others may be present but were not found on FIA plots.
- A total of 43 species identified by FIA for this NF.
- What species are present in this area (according to FIA)?
- Look at Common Name or Scientific Name, those with %Cell >0 have FIA indication of presence.
- Red maple, black cherry, sugar maple, and American beech top the list of 43 species.
- How common is each species, when it is found, i.e., it may be common only in certain areas within the unit, like bottomlands or plantations (according to FIA)?
- FIAiv shows the average importance of the species, when it is found.
- Eastern hemlock ranks 5th in overall importance across the NF (FIAsum), but 4th in importance when considering only cells where it is located—it has some particular habitats that it prefers within the NF.
- How abundant is each species, taken across the region of interest (according to FIA)?
- Abund classifies FIAsum into: Abundant, Common, Rare, Absent
- The Allegheny NF has 6 abundant species, 15 common species, and 22 rare species.
Appendix A.1.2. DISTRIB-II Model Outputs for Year 2100
- 5.
- How much confidence do we have in the models?
- ModRel gives classes of model reliability: Low, Medium, High
- The Allegheny NF table shows the following species ratings: 18 high, 24 medium, and 15 low model reliability. Three others are labeled FIA, meaning that we only provide information from FIA, and no models were developed because too few data were available for acceptable models.
- 6.
- What do the models suggest may happen to habitat suitability for the species by the year 2100, according to a high (or low) emissions scenario?
- ChngCl45 and ChngCl85 provide, for RCP 4.5 (low emissions) and RCP 8.5 (high emissions) respectively, the potential change in suitable habitat for the species by year 2100. It is important to note that a change in suitable habitat does not mean the species importance will actually change in that area by 2100, only that the habitat is expected to increase, decrease, or remain unchanged in suitability for that species over time.
- The Allegheny NF table shows 15 species with large or very large decrease in suitable habitat according to RCP 8.5, but also 4 species each for small decrease, small increase, or no change, 13 species for large increase, and 14 species with new habitat appearing by century’s end under high emissions.
- 7.
- According to literature, how adaptable are the species to the direct and indirect impacts of a changing climate?
- Adapt shows a color-coded rating (Low—pink, Medium—yellow, High—green) of a compilation of modification factors that estimate the adaptability of the species across their range.
- For Allegheny NF, 15 species are classed as highly adaptable, 35 as medium, and 8 as low in adaptability.
- 8.
- How capable might the species be for coping with the changing climate, within the region of interest, under a low (or high) emissions scenario?
- Capabil45 (or Capabil85) gives an estimate of the species capability to cope, based on its change in habitat suitability (ChngCl45 or ChngCl85), its Adapt, and its Abund in the region of interest. If the species is abundant locally, it is assumed to also be in refugia and niches somewhat buffered from the climate impacts. Ranks are coded Very Good, Good, Fair, Poor, Very Poor, FIA only (no model, no Capability assigned), NNIS (no model, non-native invasive species), Unknown (insufficient data to model), and New Habitat (potential to migrate into the region).
- Under a scenario of RCP 8.5, the Allegheny table shows 7 species with very good and 18 with good capability, 6 with fair capability, and 3 with poor and 6 with lost capability, along with 14 with new habitat, 2 non-native invasive species, and 1 species with only FIA data.
- 9.
- What species might be considered most (or least) vulnerable to the changing climate in this area?
- Those species listed under Capabil45 (or Capabil85) as Poor, Very Poor, or Lost will be vulnerable according to our assessments; those listed as Very Good, Good, or Fair are treated as less vulnerable.
- Most vulnerable (lost) species for the Allegheny NF under RCP 8.5 are red spruce, red pine, black ash, quaking aspen, pin cherry, and white spruce. Least vulnerable (very good) are: chestnut oak, northern red oak, white oak, white ash, black oak, eastern white pine, and American basswood. Of course, white ash is under severe threat of emerald ash borer, which the models could not adequately consider.
Appendix A.1.3. SHIFT Model Outputs for Year 2100
- 10.
- Natural migration of the species has been modeled with SHIFT—it predicts where the species may migrate within the next 100 years that is currently unoccupied by the species. Assuming a generous migration rate of ~50 km/century, where is the species likely to colonize?
- SHIFT45 or SHIFT85 highlights those species that have the best chance of migrating into the region within 100 years. Those species labeled ‘Migrate+’ have both new suitable habitat and some probability of colonization within 100 years, while those labeled ‘Migrate++’ have a stronger potential for colonization according to the models.
- Species with potential to migrate into the Allegheny NF within 100 years (under RCP 8.5) include: post oak, shortleaf pine, sweetgum, sycamore, and eastern redcedar (all Migrate++), flowering dogwood, sourwood, Virginia pine, and eastern redbud (all Migrate+). Under RCP 4.5, only sycamore and sourwood appear on the list.
- 11.
- What species are relatively rare in the region, but are poised to expand or infill within the region under the changing climate?
- SHIFT45 or SHIFT85 labels those relatively rare species that could expand within the region as ‘Infill+’ or ‘Infill++’, depending on the strength of the modeled indicators.
- The table for the Allegheny shows mockernut hickory, bitternut hickory, scarlet oak, and shagbark hickory as the best candidates for infilling (Infill++), followed by green ash and American elm (Infill+). Of course, these last two species would need to be resistant varieties to withstand current plagues against them.
- 12.
- What rare species are likely present in the region based on our model outputs, but the forest inventories missed them?
- SHIFT45 or SHIFT85 labels those relatively rare species are likely present within the region as ‘Likely+’ or ‘Likely++’, depending on the strength of the modeled indicators.
- The data predict that black walnut, not found in FIA plots, are now somewhere on the Allegheny NF or its buffer.
- 13.
- What species might be useful for selecting as candidates for planting in the face of a changing climate?
- SSO is the species selection option to assist in decisions regarding promoting the species, where 1 indicates the species is currently present and has at least a fair capability to cope, 2 indicates the species is rare or close to the NF boundary and has a good chance of spreading inside the NF, 3 indicates the species is not recorded in FIA plots but does have some chance of getting colonized within 100 years into the NF, and 0 indicates further evaluation may be required. Generally, managers might consider selecting species first with SSO = 1, then SSO = 2, then SSO = 3, then SSO = 0.
- For the Allegheny NF, 30 species present now have at least a fair capability to cope with the changing climate (SSO = 1); the models suggest these species are most suited for planting in the area. Three species are rare inside the NF (bitternut and mockernut hickory, slippery elm) and have potential to expand and could be suited for planting (SSO = 2) (green ash is also listed but of course would not be recommended due to emerald ash borer). Twelve additional species are marked as having some chance (>0% chance, SSO = 3) of naturally migrating there within 100 years and could be suited for planting, though with more risk of failure as compared with SSO 1 or 2. The remaining 14 species have SSO = 0, and are generally not suited for planting, though they still can be considered as candidates for planting under certain local conditions or manager preference.
- 14.
- What portion of the area has at least a 2% chance of getting colonized over 100 years? (note: these variables are only on the long file available as Supplementary File 2—Allegheny table)
- %2Col estimates, for any species, the percentage of the region of interest with at least 2% chance of colonization, and is not already occupied according to FIA. One cutoff we use for potential planting guidelines is that the species requires at least 5% of the region of interest to have >2% chance of being colonized; often coded “Migrate+” or “Migrate++” under SHIFT45/SHIFT85, and coded ‘3’ under SSO.
- The long form of the Allegheny table lists 49 species with at least 5% of the NF having at least a 2% chance of getting colonized over 100 years. These include species that are already present and potentially spreading to unoccupied locations within the NF.
- 15.
- What portion of the area has greater than 0% chance of getting colonized over 100 years? (note: these variables are only on the long file available as Supplementary File 2—Allegheny table)
- %0Col provides the percentage of the area, not already occupied according to FIA, that has >0% chance of getting colonized.
- For the Allegheny NF, 54 species (out of 60) show some area with at least some chance of getting colonized over 100 years. These also include species that are already present and potentially spreading to unoccupied locations within the NF.
- 16.
- What portion of the area has at least a 50% chance of getting colonized over 100 years? (note: these variables are only on the long file available as Supplementary File 2—Allegheny table)
- %50Col provides the percentage of the area, not already occupied according to FIA, that has >50% chance of getting colonized.
- Within the Allegheny, 41 species have at least some territory with >50% chance of colonization. These also include species that are already present and potentially spreading to unoccupied locations within the NF.
- 17.
- What species may have the highest (or at least some) chance of migrating and finding suitable habitat within 100 years (under low and high emissions)? (note: these variables are only on the long file available as Supplementary File 2—Allegheny table)
- HQCL45 (low emissions) and HQCL85 (high emissions) are indices that use a weighted average calculation (based on habitat quality and colonization likelihood), to assign a score for the potential to migrate into suitable habitat—the higher the value, the greater the chance, with values 1 or greater indicating the presence of both suitable and colonizable habitats.
- Within the Allegheny NF, of the 14 species with new habitat (question #8), 12 species have HQCL85 greater than or equal to 1.0 (indicating the presence of both suitable habitat and colonizable habitat), while for HQCL45, the number is only 3 species.
References
- IPCC Summary for Policymakers. In Global Warming of 1.5 °C. An IPCC Special Report on the Impacts of Global Warming of 1.5 °C Above Pre-Industrial Levels and Related Global Greenhouse Gas Emission Pathways, in the Context of Strengthening the Global Response to the Threat of Climate Change, Sustainable Development, and Efforts to Eradicate Poverty; World Meteorological Organization: Geneva, Switzerland, 2018; p. 32.
- USGCRP. Impacts, Risks, and Adaptation in the United States: Fourth National Climate Assessment; Global Change Research Program: Washington, DC, USA, 2018; Volume II: Report-In-Brief, p. 186.
- IPBES. Summary for Policymakers of the Global Assessment Report on Biodiversity and Ecosystem Services of the Intergovernmental Science-Policy Platform on Biodiversity and Ecosystem Services; Intergovernmental Science-Policy Platform on Biodiversity and Ecosystem Services: Bonn, Germany, 2019; p. 44. [Google Scholar]
- Urban, M.C.; Bocedi, G.; Hendry, A.P.; Mihoub, J.-B.; Pe’er, G.; Singer, A.; Bridle, J.R.; Crozier, L.G.; De Meester, L.; Godsoe, W.; et al. Improving the forecast for biodiversity under climate change. Science 2016, 353, aad8466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Araújo, M.B.; Anderson, R.P.; Márcia Barbosa, A.; Beale, C.M.; Dormann, C.F.; Early, R.; Garcia, R.A.; Guisan, A.; Maiorano, L.; Naimi, B.; et al. Standards for distribution models in biodiversity assessments. Sci. Adv. 2019, 5, eaat4858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Higgins, S.I.; O’Hara, R.B.; Römermann, C. A niche for biology in species distribution models. J. Biogeogr. 2012, 39, 2091–2095. [Google Scholar] [CrossRef]
- Dullinger, S.; Gattringer, A.; Thuiller, W.; Moser, D.; Zimmermann, N.E.; Guisan, A.; Willner, W.; Plutzar, C.; Leitner, M.; Mang, T.; et al. Extinction debt of high-mountain plants under twenty-first-century climate change. Nat. Clim. Chang. 2012, 2, 619. [Google Scholar] [CrossRef]
- Zurell, D.; Thuiller, W.; Pagel, J.; Cabral, J.S.; Münkemüller, T.; Gravel, D.; Dullinger, S.; Normand, S.; Schiffers, K.H.; Moore, K.A.; et al. Benchmarking novel approaches for modelling species range dynamics. Glob. Chang. Biol. 2016, 22, 2651–2664. [Google Scholar] [CrossRef] [PubMed]
- Reynolds, K.M. Integrated decision support for sustainable forest management in the United States: Fact or fiction? Comput. Electron. Agric. 2005, 49, 6–23. [Google Scholar] [CrossRef]
- Iverson, L.; McKenzie, D. Tree-Species range shifts in a changing climate—Detecting, modeling, assisting. Landsc. Ecol. 2013, 28, 879–889. [Google Scholar] [CrossRef]
- Prasad, A.M.; Iverson, L.R.; Matthews, S.N.; Peters, M.P. A multistage decision support framework to guide tree species management under climate change via habitat suitability and colonization models, and a knowledge-based scoring system. Landsc. Ecol. 2016, 31, 2187–2204. [Google Scholar] [CrossRef]
- Brandt, L.; He, H.; Iverson, L.; Thompson, F.; Butler, P.; Handler, S.; Janowiak, M.; Swanston, C.; Albrecht, M.; Blume-Weaver, R.; et al. Central Hardwoods Ecosystem Vulnerability Assessment and Synthesis: A Report from the Central Hardwoods Climate Change Response Framework Project; U.S. Department of Agriculture, Forest Service, Northern Research Station: Newtown Square, PA, USA, 2014; p. 254.
- Handler, S.; Duveneck, M.; Iverson, L.; Peters, E.; Scheller, R.; Wythers, K.; Brandt, L.; Butler, P.; Janowiak, M.; Swanston, C.; et al. Minnesota Forest Ecosystem Vulnerability Assessment and Synthesis: A Report from the Northwoods Climate Change Response Framework; Gen. Tech. Rep. NRS-133; U.S. Department of Agriculture, Forest Service, Northern Research Station: Newtown Square, PA, USA, 2014.
- Handler, S.; Duveneck, M.J.; Iverson, L.; Peters, E.; Scheller, R.; Wythers, K.; Brandt, L.; Butler, P.; Janowiak, M.; Swanston, C.; et al. Michigan Forest Ecosystem Vulnerability Assessment and Synthesis: A Report from the Northwoods Climate Change Response Framework; Gen. Tech. Rep. NRS-129; U.S. Department of Agriculture, Forest Service, Northern Research Station: Newtown Square, PA, USA, 2014.
- Janowiak, M.K.; Iverson, L.R.; Mladenoff, D.J.; Peters, E.; Wythers, K.R.; Xi, W.; Brandt, L.A.; Butler, P.R.; Handler, S.D.; Shannon, P.D.; et al. Forest Ecosystem Vulnerability Assessment and Synthesis for Northern Wisconsin and Western Upper Michigan: A Report from the Northwoods Climate Change Response Framework Project; Gen. Tech. Rep. NRS-136; U.S. Department of Agriculture, Forest Service, Northern Research Station: Newtown Square, PA, USA, 2014; p. 247.
- Brandt, L.A.; Derby Lewis, A.; Scott, L.; Darling, L.; Fahey, R.T.; Iverson, L.; Nowak, D.J.; Bodine, A.R.; Bell, A.; Still, S.; et al. Chicago Wilderness Region Urban Forest Vulnerability Assessment and Synthesis: A Report from the Urban Forestry Climate Change Response Framework Chicago Wilderness Pilot Project; U.S. Department of Agriculture, Forest Service, Northern Research Station: Newtown Square, PA, USA, 2017; p. 142.
- Butler-Leopold, P.R.; Iverson, L.R.; Thompson, F.R., III; Brandt, L.A.; Handler, S.D.; Janowiak, M.K.; Shannon, P.D.; Swanston, C.W.; Bearer, S.; Bryan, A.M.; et al. Mid-Atlantic Forest Ecosystem Vulnerability Assessment and Synthesis: A Report from the Mid-Atlantic Climate Change Response Framework Project; Gen. Tech. Rep. NRS-181; U.S. Department of Agriculture, Forest Service, Northern Research Station: Newtown Square, PA, USA, 2018; p. 365.
- Janowiak, M.K.; D’Amato, A.W.; Swanston, C.W.; Iverson, L.; Thompson III, F.; Dijak, W.; Matthews, S.; Peters, M.; Prasad, A.; Fraser, J.; et al. New England and Northern New York Forest Ecosystem Vulnerability Assessment and Synthesis: A Report from the New England Climate Change Response Framework Project; Gen. Tech. Rep. NRS-173; U.S. Department of Agriculture, Forest Service, Northern Research Station: Newtown Square, PA, USA, 2018; p. 234.
- Matthews, S.N.; Iverson, L.R.; Prasad, A.M.; Peters, M.P.; Rodewald, P.G. Modifying climate change habitat models using tree species-specific assessments of model uncertainty and life history factors. For. Ecol. Manag. 2011, 262, 1460–1472. [Google Scholar] [CrossRef]
- Iverson, L.R.; Peters, M.P.; Prasad, A.M.; Matthews, S.N. Analysis of climate change impacts on tree species of the eastern US: Results of DISTRIB-II modeling. Forests 2019, 10, 302. [Google Scholar] [CrossRef]
- Peters, M.; Iverson, L.; Prasad, A.; Matthews, S. Utilizing the density of inventory samples to define a hybrid lattice for a macro-level species distribution model. Ecol. Evol. 2019, 9, 8876–8899. [Google Scholar] [CrossRef] [PubMed]
- Hagen-Zanker, A. Comparing Continuous Valued Raster Data: A Cross Disciplinary Literature Scan; Research Institute for Knowledge Systems: Bilthoven, The Netherlands, 2006. [Google Scholar]
- Iverson, L.R.; Prasad, A.M.; Matthews, S.N. Modeling potential climate change impacts on the trees of the northeastern United States. Mitig. Adapt. Strateg. Glob. Chang. 2008, 13, 487–516. [Google Scholar] [CrossRef]
- Prasad, A.M.; Iverson, L.R.; Liaw, A. Newer classification and regression tree techniques: Bagging and random forests for ecological prediction. Ecosystems 2006, 9, 181–199. [Google Scholar] [CrossRef]
- Iverson, L.R.; Prasad, A.M.; Matthews, S.N.; Peters, M. Estimating potential habitat for 134 eastern US tree species under six climate scenarios. For. Ecol. Manag. 2008, 254, 390–406. [Google Scholar] [CrossRef]
- Prasad, A.M.; Gardiner, J.; Iverson, L.; Matthews, S.; Peters, M. Exploring tree species colonization potentials using a spatially explicit simulation model: Implications for four oaks under climate change. Glob. Chang. Biol. 2013, 19, 2196–2208. [Google Scholar] [CrossRef]
- McLachlan, J.S.; Clark, J.S.; Manos, P.S. Molecular indicators of tree migration capacity under rapid climate change. Ecology 2005, 86, 2007–2017. [Google Scholar] [CrossRef]
- Davis, M.B.; Shaw, R.G. Range shifts and adaptive responses to quaternary climate change. Science 2001, 292, 673–679. [Google Scholar] [CrossRef]
- Morelli, T.L.; Daly, C.; Dobrowski, S.Z.; Dulen, D.M.; Ebersole, J.L.; Jackson, S.T.; Lundquist, J.D.; Millar, C.I.; Maher, S.P.; Monahan, W.B.; et al. Managing Climate Change Refugia for Climate Adaptation. PLoS ONE 2016, 11, e0159909. [Google Scholar] [CrossRef]
- Anderson, M.G.; Clark, M.; Sheldon, A.O. Estimating climate resilience for conservation across geophysical settings. Conserv. Biol. 2014, 28, 959–970. [Google Scholar] [CrossRef]
- Iverson, L.; Matthews, S.; Prasad, A.; Peters, M.; Yohe, G. Development of risk matrices for evaluating climatic change responses of forested habitats. Clim. Chang. 2012, 114, 231–243. [Google Scholar] [CrossRef] [Green Version]
- Iverson, L.; Prasad, A.M.; Matthews, S.; Peters, M. Lessons learned while integrating habitat, dispersal, disturbance, and life-history traits into species habitat models under climate change. Ecosystems 2011, 14, 1005–1020. [Google Scholar] [CrossRef]
- Iverson, L.; Prasad, A.M.; Matthews, S.; Peters, M. Chapter 2. Climate as an agent of change in forest landscapes. In Forest Landscapes and Global Change: Challenges for Research and Management; Azevedo, J., Perera, A., Pinto, M., Eds.; Springer: New York, NY, USA, 2014; pp. 29–50. [Google Scholar]
- Iverson, L.R.; Prasad, A.; Scott, C.T. Preparation of forest inventory and analysis (FIA) and state soil geographic data base (STATSGO) data for global change research in the eastern United States. In Proceedings, 1995 Meeting of the Northern Global Change Program; General Technical Report NE-214; Hom, J., Birdsey, R., O’Brian, K., Eds.; U.S. Department of Agriculture, Forest Service, Northeastern Forest Experiment Station: Radnor, PA, USA, 1996; pp. 209–214. [Google Scholar]
- Iverson, L.R.; Prasad, A.M. Potential changes in tree species richness and forest community types following climate change. Ecosystems 2001, 4, 186–199. [Google Scholar] [CrossRef]
- Iverson, L.R.; Prasad, A.M. Predicting abundance of 80 tree species following climate change in the eastern United States. Ecol. Monogr. 1998, 68, 465–485. [Google Scholar] [CrossRef]
- Iverson, L.R.; Prasad, A.M. Potential redistribution of tree species habitat under five climate change scenarios in the eastern US. For. Ecol. Manag. 2002, 155, 205–222. [Google Scholar] [CrossRef]
- Iverson, L.R.; Prasad, A.M.; Hale, B.J.; Sutherland, E.K. Atlas of Current and Potential Future Distributions of Common Trees of the Eastern United States; General Technical Report NE-265; Northeastern Research Station, USDA Forest Service: Radnor, PA, USA, 1999; p. 245.
- Iverson, L.R.; Prasad, A.M.; Schwartz, M.W. Predicting potential changes in suitable habitat and distribution by 2100 for tree species of the eastern United States. J. Agric. Meteorol. 2005, 61, 29–37. [Google Scholar] [CrossRef]
- Matthews, S.N.; Iverson, L.R.; Peters, M.P.; Prasad, A.M.; Subburayalu, S. Assessing and comparing risk to climate changes among forested locations: Implications for ecosystem services. Landsc. Ecol. 2014, 29, 213–228. [Google Scholar] [CrossRef]
- Iverson, L.R.; Schwartz, M.W.; Prasad, A. How fast and far might tree species migrate under climate change in the eastern United States? Glob. Ecol. Biogeogr. 2004, 13, 209–219. [Google Scholar] [CrossRef]
- Iverson, L.R.; Schwartz, M.W.; Prasad, A.M. Potential colonization of new available tree species habitat under climate change: An analysis for five eastern US species. Landsc. Ecol. 2004, 19, 787–799. [Google Scholar] [CrossRef]
- Iverson, L.R.; Prasad, A.M.; Schwartz, M.W. Modeling potential future individual tree-species distributions in the Eastern United States under a climate change scenario: A case study with Pinus virginiana. Ecol. Model. 1999, 115, 77–93. [Google Scholar] [CrossRef]
- Bouchard, M.; Aquilué, N.; Périé, C.; Lambert, M.-C. Tree species persistence under warming conditions: A key driver of forest response to climate change. For. Ecol. Manag. 2019, 442, 96–104. [Google Scholar] [CrossRef]
- Swanston, C.W.; Janowiak, M.; Brandt, L.; Butler, P.; Handler, S.D.; Shannon, P.D.; Lewis, A.D.; Hall, K.R.; Fahey, R.T.; Scott, L.; et al. Forest Adaptation Resources: Climate Change Tools and Approaches for Land Managers, 2nd ed.; Gen. Tech. Rep. NRS-GTR-87-2; U.S. Department of Agriculture, Forest Service, Northern Research Station: Newtown Square, PA, USA, 2016; p. 161.
- Millar, C.; Stephenson, N.L. Climate change and forests of the future: Managing in the face of uncertainty. Ecol. Appl. 2007, 17, 2145–2151. [Google Scholar] [CrossRef] [PubMed]
- Nagel, L.M.; Palik, B.J.; Battaglia, M.A.; D’Amato, A.W.; Guldin, J.M.; Swanston, C.W.; Janowiak, M.K.; Powers, M.P.; Joyce, L.A.; Millar, C.I.; et al. Adaptive silviculture for climate change: A national experiment in manager-scientist partnerships to apply an adaptation framework. J. For. 2017, 115, 167–178. [Google Scholar] [CrossRef]
- Woodall, C.W.; Westfall, J.A.; D’Amato, A.W.; Foster, J.R.; Walters, B.F. Decadal changes in tree range stability across forests of the eastern U.S. For. Ecol. Manag. 2018, 429, 503–510. [Google Scholar] [CrossRef]
- Catanzaro, P.; D’Amato, A.W.; Huff, E.S. Increasing Forest Resiliency for an Uncertain Future; University of Massachusetts, Cooperative Extension Landowner Outreach Pamphlet: Boston, MA, USA, 2016; p. 32. [Google Scholar]
- Guldin, J.M. Silvicultural options in forests of the southern United States under changing climatic conditions. New For. 2019, 50, 71–87. [Google Scholar] [CrossRef]
- Loarie, S.R.; Duffy, P.B.; Hamilton, H.; Asner, G.P.; Field, C.B.; Ackerly, D.D. The velocity of climate change. Nature 2009, 462, 1052–1055. [Google Scholar] [CrossRef] [PubMed]
- Serra-Diaz, J.M.; Franklin, J.; Ninyerola, M.; Davis, F.W.; Syphard, A.D.; Regan, H.M.; Ikegami, M. Bioclimatic velocity: The pace of species exposure to climate change. Divers. Distrib. 2014, 20, 169–180. [Google Scholar] [CrossRef]
- Carroll, C.; Lawler, J.J.; Roberts, D.R.; Hamann, A. Biotic and climatic velocity identify contrasting areas of vulnerability to climate change. PLoS ONE 2015, 10, e0140486. [Google Scholar] [CrossRef] [PubMed]
- Sittaro, F.; Paquette, A.; Messier, C.; Nock Charles, A. Tree range expansion in eastern North America fails to keep pace with climate warming at northern range limits. Glob. Chang. Biol. 2017, 23, 3292–3301. [Google Scholar] [CrossRef]
- Pedlar, J.H.; McKenney, D.W.; Aubin, I.; Beardmore, T.; Beaulieru, J.; Iverson, L.R.; O’Neill, G.A.; Winder, R.S.; Ste-Marie, C. Placing forestry in the assisted migration debate. BioScience 2012, 62, 835–842. [Google Scholar] [CrossRef]
- Scheffers, B.R.; Pecl, G. Persecuting, protecting or ignoring biodiversity under climate change. Nat. Clim. Chang. 2019, 9, 581–586. [Google Scholar] [CrossRef]
- Currie, D.J.; Paquin, V. Large-Scale biogeographical patterns of species richness of trees. Nature 1987, 329, 326–327. [Google Scholar] [CrossRef]
- Currie, D.J. Energy and large-scale patterns of animal-and plant-species richness. Am. Nat. 1991, 137, 27–49. [Google Scholar] [CrossRef]
- Hanberry, B.B.; Nowacki, G.J. Oaks were the historical foundation genus of the east-central United States. Quat. Sci. Rev. 2016, 145, 94–103. [Google Scholar] [CrossRef]
- McShea, W.; Healy, W. Oak Forest Ecosystems: Ecology and Management for Wildlife; The Johns Hopkins University Press: Baltimore, MD, USA, 2002. [Google Scholar]
- McShea, W.J.; Healy, W.M.; Devers, P.; Fearer, T.; Koch, F.H.; Stauffer, D.; Waldon, J. Forestry matters: Decline of oaks will impact wildlife in hardwood forests. J. Wildl. Manag. 2007, 71, 1717–1728. [Google Scholar] [CrossRef]
- Johnson, P.; Shifley, S.; Rogers, R. The Ecology and Silviculture of Oaks; CABI Publishing: New York, NY, USA, 2009. [Google Scholar]
- Tallamy, D.W.; Shropshire, K.J. Ranking lepidopteran use of native versus introduced plants. Conserv. Biol. 2009, 23, 941–947. [Google Scholar] [CrossRef]
- Loftis, D.L. Upland Oak Regeneration and Management; U.S. Department of Agriculture, Forest Service, Southern Research Station: Asheville, NC, USA, 2004; pp. 163–167.
- Nowacki, G.J.; Abrams, M.D. The demise of fire and ‘’mesophication’’ of forests in the eastern United States. Bioscience 2008, 58, 123–138. [Google Scholar] [CrossRef]
- Knott, J.A.; Desprez, J.M.; Oswalt, C.M.; Fei, S. Shifts in forest composition in the eastern United States. For. Ecol. Manag. 2019, 433, 176–183. [Google Scholar] [CrossRef]
- Hutchinson, T.F.; Yaussy, D.A.; Long, R.P.; Rebbeck, J.; Sutherland, E.K. Long-Term (13-year) effects of repeated prescribed fires on stand structure and tree regeneration in mixed-oak forests. For. Ecol. Manag. 2012, 286, 87–100. [Google Scholar] [CrossRef]
- Iverson, L.R.; Hutchinson, T.F.; Peters, M.P.; Yaussy, D.A. Long-Term response of oak-hickory regeneration to partial harvest and repeated fires: Influence of light and moisture. Ecosphere 2017, 8, e01642. [Google Scholar] [CrossRef]
- Clark, J.; Iverson, L.; Woodall, C.W.; Allen, C.D.; Bell, D.M.; Bragg, D.C.; D’Amato, A.W.; Davis, F.W.; Hersh, M.H.; Ibanez, I.; et al. Impacts of increasing drought on forest dynamics, structure, diversity, and management. In Effects of Drought on Forests and Rangelands in the United States: A Comprehensive Science Synthesis; Gen. Tech. Rep. WO-93b; Vose, J., Clark, J., Luce, C., Patel-Weynand, T., Eds.; U.S. Department of Agriculture, Forest Service, Washington Office: Washington, DC, USA, 2016; pp. 59–96. [Google Scholar]
- Vose, J.; Elliott, K.J. Oak, fire, and global change in the Eastern USA: What might the future hold? Fire Ecol. 2016, 12, 160–179. [Google Scholar] [CrossRef]
- Iverson, L.R.; Thompson, F.R.; Matthews, S.; Peters, M.; Prasad, A.; Dijak, W.D.; Fraser, J.; Wang, W.J.; Hanberry, B.; He, H.; et al. Multi-model comparison on the effects of climate change on tree species in the eastern U.S.: Results from an enhanced niche model and process-based ecosystem and landscape models. Landsc. Ecol. 2017, 32, 1327–1346. [Google Scholar] [CrossRef]
- Iverson, L.; Peters, M.; Matthews, S.; Prasad, A.; Hutchinson, T.; Bartig, J.; Rebbeck, J.; Yaussy, D.A.; Stout, S.; Nowacki, G. Adapting oak management in an age of ongoing mesophication but warming climate. In Oak Symposium: Sustaining Oak Forests in the 21st Century Through Science-Based Management; e-Gen. Tech. Rep. SRS-237; Clark, S.L., Schweitzer, C.J., Eds.; U.S. Department of Agriculture Forest Service, Southern Research Station: Asheville, NC, USA, 2019; pp. 35–45. [Google Scholar]
- Riitters, K.H.; Wickham, J.D.; O’Neill, R.V.; Jones, K.B.; Smith, E.R.; Coulston, J.W.; Wade, T.G.; Smith, J.H. Fragmentation of continental United States forests. Ecosystems 2002, 5, 815–822. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Change Class | Adaptability | ||
|---|---|---|---|
| High | Medium | Low | |
| Large increase | Very Good | Very Good | Good |
| Small increase | Very Good | Good | Fair |
| No change | Good | Fair | Poor |
| Small decrease | Fair | Poor | Poor |
| Large decrease | Fair | Poor | Very Poor |
| Other capability classes: | |||
| Lost | All suitable habitat lost | ||
| New Habitat | New habitat appearing | ||
| FIA Only | Unacceptable model for future, only FIA reported | ||
| NNIS | Non-native invasive species, only FIA reported | ||
| Unknown | Modeled as present, unknown | ||
| Class | Capability | HQCL | %OccCol | %2Col |
|---|---|---|---|---|
| Infill+ | Fair, Poor | ≥1 | >0–50 | any |
| Infill++ | Good, Very Good | ≥1 | >10–50 | any |
| Likely+ | New Habitat, Unknown | any | >0 | any |
| Likely++ | New Habitat, Unknown | ≥1 | >2 | any |
| Migrate+ | New Habitat | =1 | 0 | >0 |
| Migrate++ | New Habitat | >1 | 0 | >0 |
| Identifier | Name | Longitude | Latitude | Area, km2 | Buffer Area, km2 | Number Species | Number Infill | Number Migrate |
|---|---|---|---|---|---|---|---|---|
| 1 | Allegheny | −79 | 41.7 | 2996.5 | 8200 | 43 | 6 | 11 |
| 2 | Angelina | −94.4 | 31.3 | 1602.8 | 8800 | 61 | 22 | 1 |
| 3 | Apalachicola | −84.7 | 30.2 | 2555.9 | 8044 | 64 | 18 | 1 |
| 4 | Bienville | −89.5 | 32.3 | 1572 | 8900 | 67 | 16 | 2 |
| 5 | Black_Kettle NG | −99.5 | 35.7 | 982 | 8196 | 18 | 2 | 6 |
| 6 | Caddo NG | −96 | 33.6 | 277.3 | 9100 | 46 | 13 | 0 |
| 7 | Chattahoochee | −84.2 | 34.7 | 5950.2 | 9700 | 72 | 24 | 9 |
| 8 | Chequamegon | −90.8 | 46 | 4209.7 | 11,815 | 40 | 5 | 9 |
| 9 | Cherokee | −83.1 | 35.8 | 5950.2 | 9600 | 74 | 35 | 9 |
| 10 | Chippewa | −94.1 | 47.4 | 6465.5 | 8800 | 34 | 6 | 11 |
| 11 | Conecuh | −86.6 | 31.1 | 692.9 | 8500 | 60 | 18 | 4 |
| 12 | Croatan | −77.1 | 35 | 1243.7 | 8034 | 62 | 17 | 2 |
| 13 | Daniel_Boone | −83.9 | 37.3 | 8282.7 | 14,100 | 77 | 10 | 4 |
| 14 | Davy_Crockett | −95.1 | 31.3 | 1602.8 | 8400 | 55 | 15 | 1 |
| 15 | Delta | −90.8 | 32.8 | 490.7 | 8000 | 62 | 30 | 3 |
| 16 | DeSoto | −89 | 31.1 | 3252.4 | 8700 | 67 | 18 | 0 |
| 17 | Finger_Lakes | −76.8 | 42.6 | 58.4 | 8300 | 62 | 11 | 8 |
| 18 | Francis_Marion | −79.8 | 33.3 | 1696.1 | 8221 | 70 | 24 | 1 |
| 19 | George_Washington | −79.3 | 38.2 | 7270.9 | 14,300 | 73 | 16 | 10 |
| 20 | Green_Mountain | −73 | 43.4 | 2543 | 8800 | 42 | 6 | 14 |
| 21 | Hiawatha | −86 | 46.2 | 5197.3 | 9549 | 35 | 7 | 14 |
| 22 | Holly_Springs | −89.2 | 34.7 | 1906 | 8500 | 70 | 13 | 4 |
| 23 | Homochitto | −90.9 | 31.5 | 1540.6 | 8400 | 72 | 17 | 2 |
| 24 | Hoosier | −86.5 | 38.5 | 2612 | 8100 | 65 | 11 | 4 |
| 25 | Huron | −84 | 44.6 | 2787.3 | 8078 | 39 | 6 | 12 |
| 26 | Jefferson | −81.2 | 37.1 | 4926.7 | 15,100 | 76 | 11 | 8 |
| 27 | Kisatchie | −92.8 | 31.7 | 4195.7 | 9800 | 74 | 12 | 2 |
| 28 | Lyndon_B_Johnson NG | −97.6 | 33.4 | 465.5 | 9200 | 32 | 14 | 1 |
| 29 | Manistee | −85.9 | 43.9 | 5377.5 | 8566 | 47 | 7 | 6 |
| 30 | Mark_Twain | −91.7 | 37.3 | 12,237.4 | 23,900 | 66 | 15 | 5 |
| 31 | Midewin NG | −88.1 | 41.4 | 104.3 | 8350 | 31 | 19 | 12 |
| 32 | Monongahela | −79.9 | 38.6 | 7270.9 | 10,500 | 61 | 8 | 11 |
| 33 | Nantahala | −83.6 | 35.2 | 5950.2 | 9700 | 65 | 17 | 9 |
| 34 | Nicolet | −88.7 | 45.7 | 3890.7 | 8200 | 39 | 8 | 6 |
| 35 | Ocala | −81.8 | 29.2 | 1791.6 | 8177 | 42 | 4 | 5 |
| 36 | Oconee | −83.5 | 33.4 | 1049.8 | 8700 | 62 | 10 | 3 |
| 37 | Osceola | −82.5 | 30.4 | 932.4 | 8400 | 44 | 12 | 6 |
| 38 | Ottawa | −89.2 | 46.5 | 3890.7 | 9559 | 36 | 7 | 11 |
| 39 | Ouachita | −93.9 | 34.6 | 9682.3 | 15,000 | 62 | 16 | 11 |
| 40 | Ozark | −93.5 | 35.7 | 9682.3 | 11,800 | 70 | 13 | 11 |
| 41 | Pisgah | −82.4 | 35.8 | 5950.2 | 10,100 | 70 | 12 | 12 |
| 42 | Sabine | −93.9 | 31.5 | 1602.8 | 8500 | 63 | 22 | 3 |
| 43 | Saint_Francis | −90.7 | 34.7 | 120.5 | 8700 | 63 | 45 | 3 |
| 44 | Sam_Houston | −95.4 | 30.5 | 2005.3 | 8200 | 63 | 18 | 1 |
| 45 | Shawnee | −88.9 | 37.5 | 3501 | 8000 | 73 | 13 | 5 |
| 46 | Sheyenne NG | −97.2 | 46.4 | 551.6 | 8400 | 9 | 6 | 7 |
| 47 | Sumter | −82.1 | 34.3 | 5950.2 | 8200 | 72 | 24 | 1 |
| 48 | Superior | −91.6 | 47.8 | 13,204.5 | 16,415 | 32 | 11 | 14 |
| 49 | Talladega | −86.5 | 33.2 | 3042.6 | 9700 | 68 | 15 | 7 |
| 50 | Tombigbee | −89.2 | 33.7 | 713.1 | 8700 | 69 | 20 | 2 |
| 51 | Tuskegee | −85.6 | 32.5 | 63.2 | 8100 | 70 | 21 | 3 |
| 52 | Uwharrie | −79.9 | 35.4 | 889.2 | 8200 | 73 | 18 | 4 |
| 53 | Wayne | −82 | 39.3 | 3451.8 | 10,800 | 71 | 13 | 5 |
| 54 | White_Mountain | −71.4 | 44.2 | 3511.8 | 8800 | 37 | 9 | 14 |
| 55 | WB_Bankhead | −87.3 | 34.2 | 1409.3 | 8900 | 71 | 0 | 0 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Iverson, L.R.; Prasad, A.M.; Peters, M.P.; Matthews, S.N. Facilitating Adaptive Forest Management under Climate Change: A Spatially Specific Synthesis of 125 Species for Habitat Changes and Assisted Migration over the Eastern United States. Forests 2019, 10, 989. https://doi.org/10.3390/f10110989
Iverson LR, Prasad AM, Peters MP, Matthews SN. Facilitating Adaptive Forest Management under Climate Change: A Spatially Specific Synthesis of 125 Species for Habitat Changes and Assisted Migration over the Eastern United States. Forests. 2019; 10(11):989. https://doi.org/10.3390/f10110989
Chicago/Turabian StyleIverson, Louis R., Anantha M. Prasad, Matthew P. Peters, and Stephen N. Matthews. 2019. "Facilitating Adaptive Forest Management under Climate Change: A Spatially Specific Synthesis of 125 Species for Habitat Changes and Assisted Migration over the Eastern United States" Forests 10, no. 11: 989. https://doi.org/10.3390/f10110989
APA StyleIverson, L. R., Prasad, A. M., Peters, M. P., & Matthews, S. N. (2019). Facilitating Adaptive Forest Management under Climate Change: A Spatially Specific Synthesis of 125 Species for Habitat Changes and Assisted Migration over the Eastern United States. Forests, 10(11), 989. https://doi.org/10.3390/f10110989