The Soil Microbiome of the Laurel Forest in Garajonay National Park (La Gomera, Canary Islands): Comparing Unburned and Burned Habitats after a Wildfire

 , , and
, , and
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Sampling Localities
2.2. Physicochemical Analysis of the Soils
2.3. DNA Extraction, PCR Amplification, and Pyrosequencing of Bacterial Soil DNA
2.4. Pyrosequencing Data Analysis
3. Results
3.1. Chemical Characteristics of Soils from Laurel Forest in Garajonay NP
3.2. Alpha Diversity of the Microbial Communities
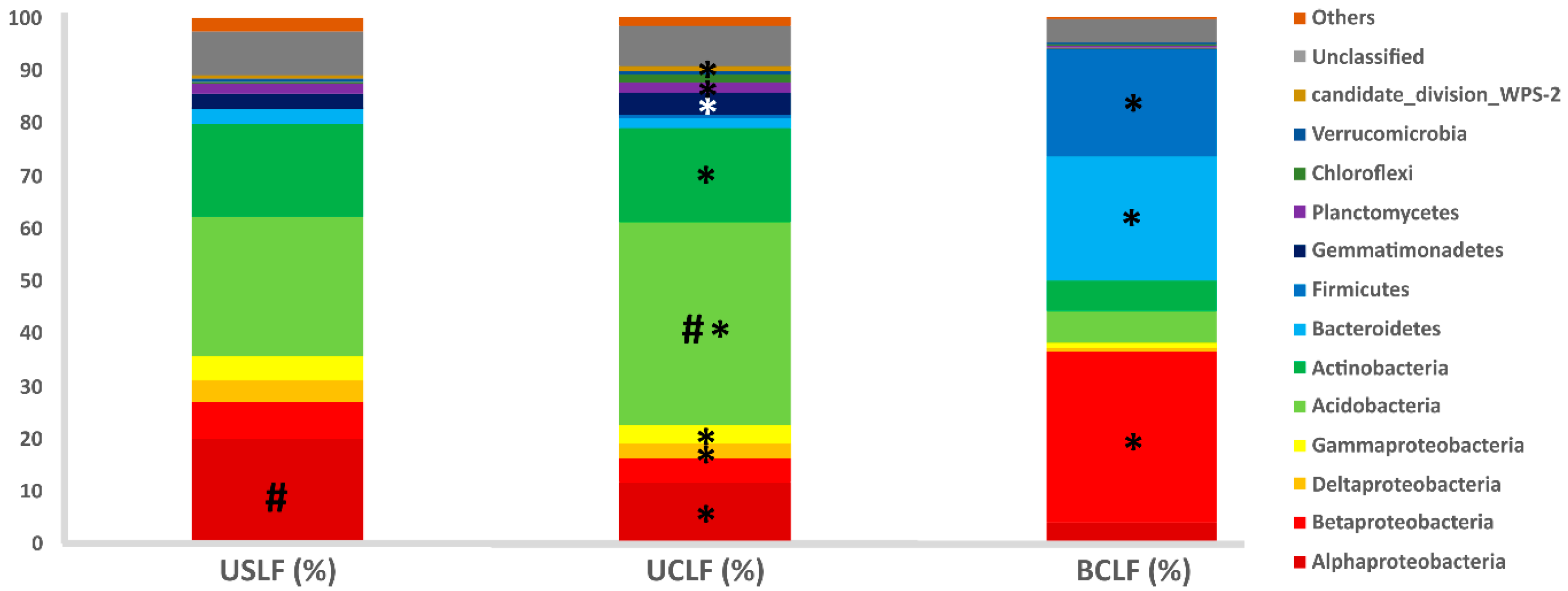
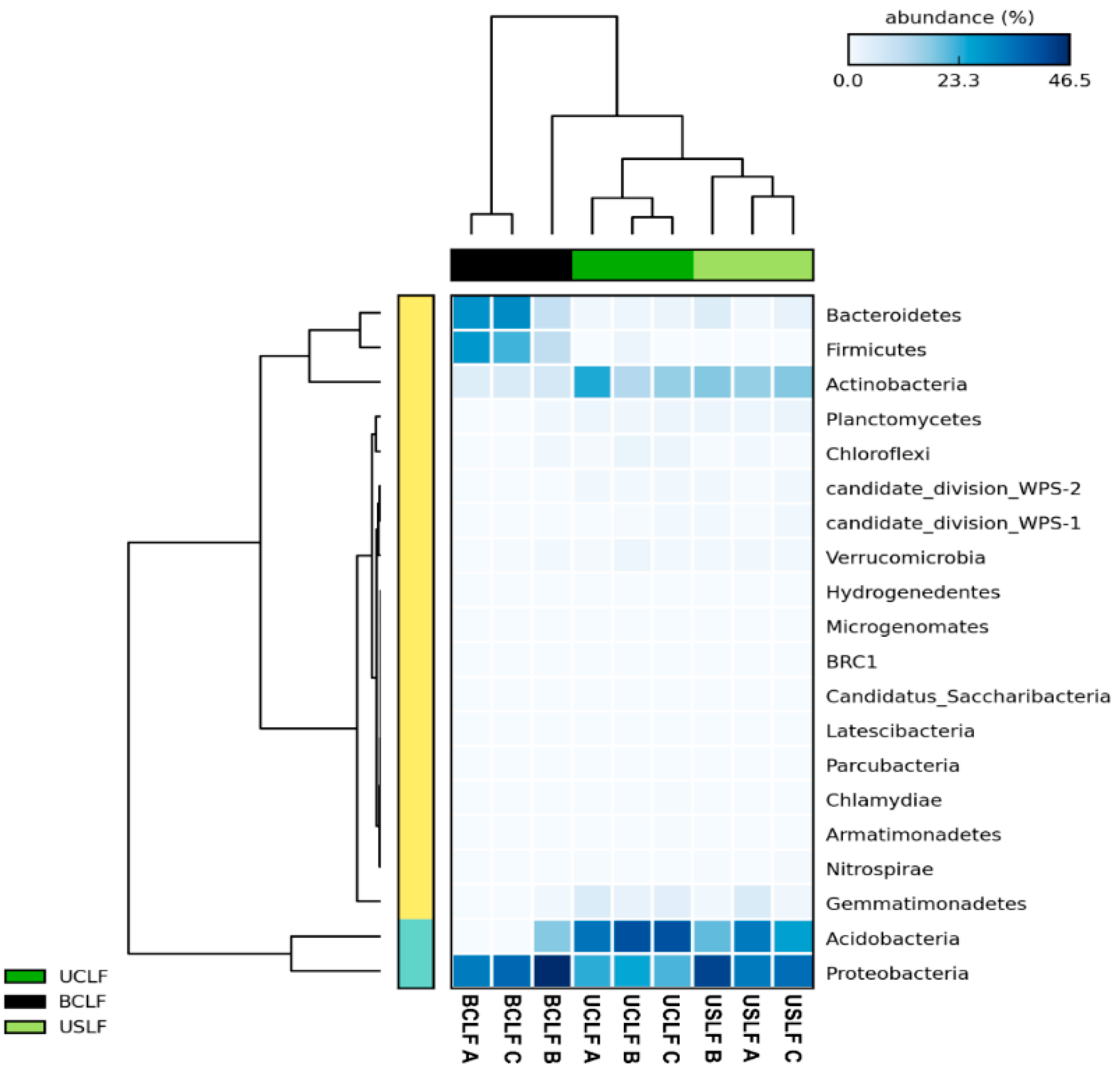
3.3. Bacterial Community Composition of the Soil Microbiome in Undisturbed Laurel Forests
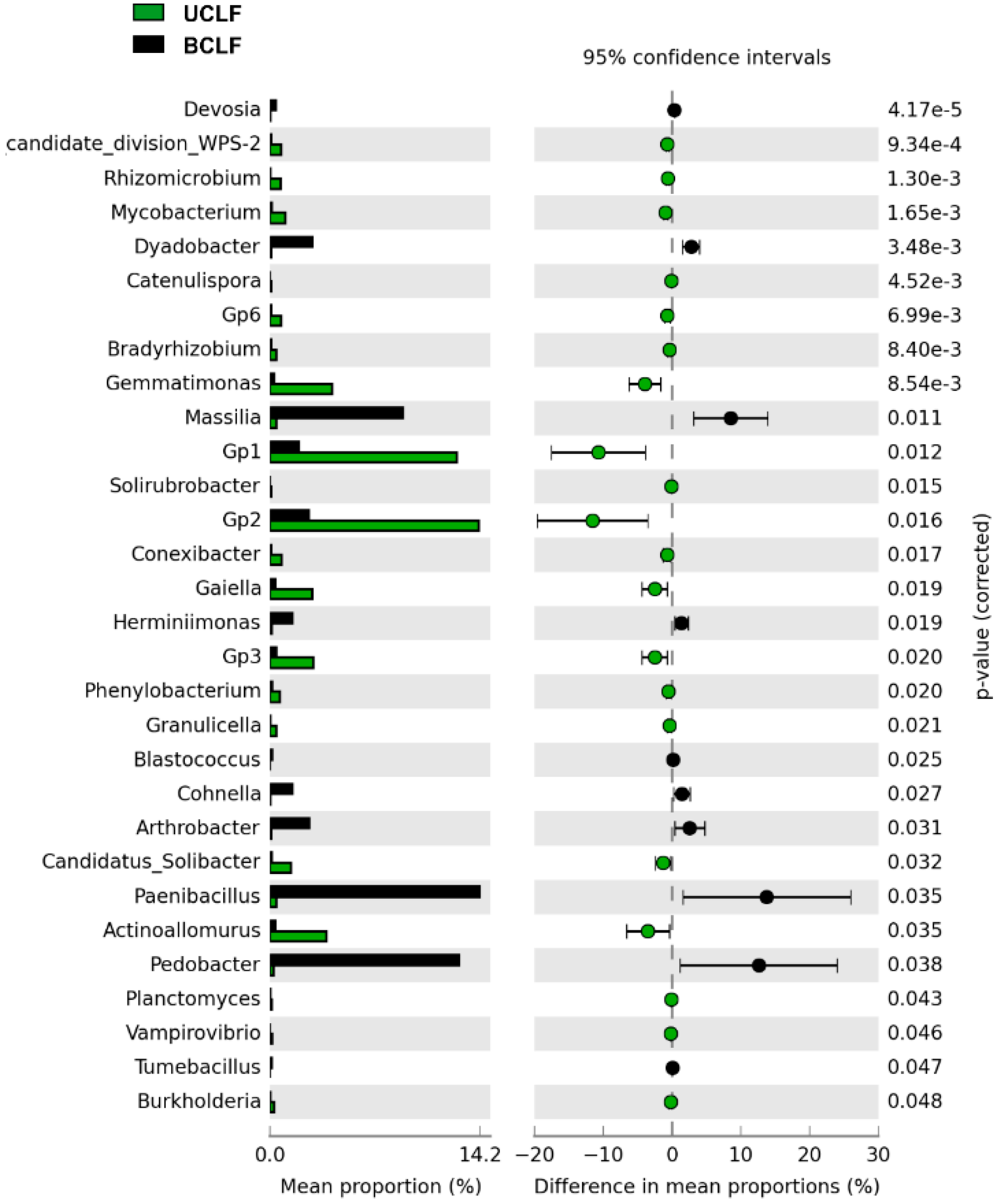
3.4. Effect of a Wildfire on the Soil Microbiome in the Crest Laurel Forest
3.5. Influence of Environmental Variables on Soil Communities
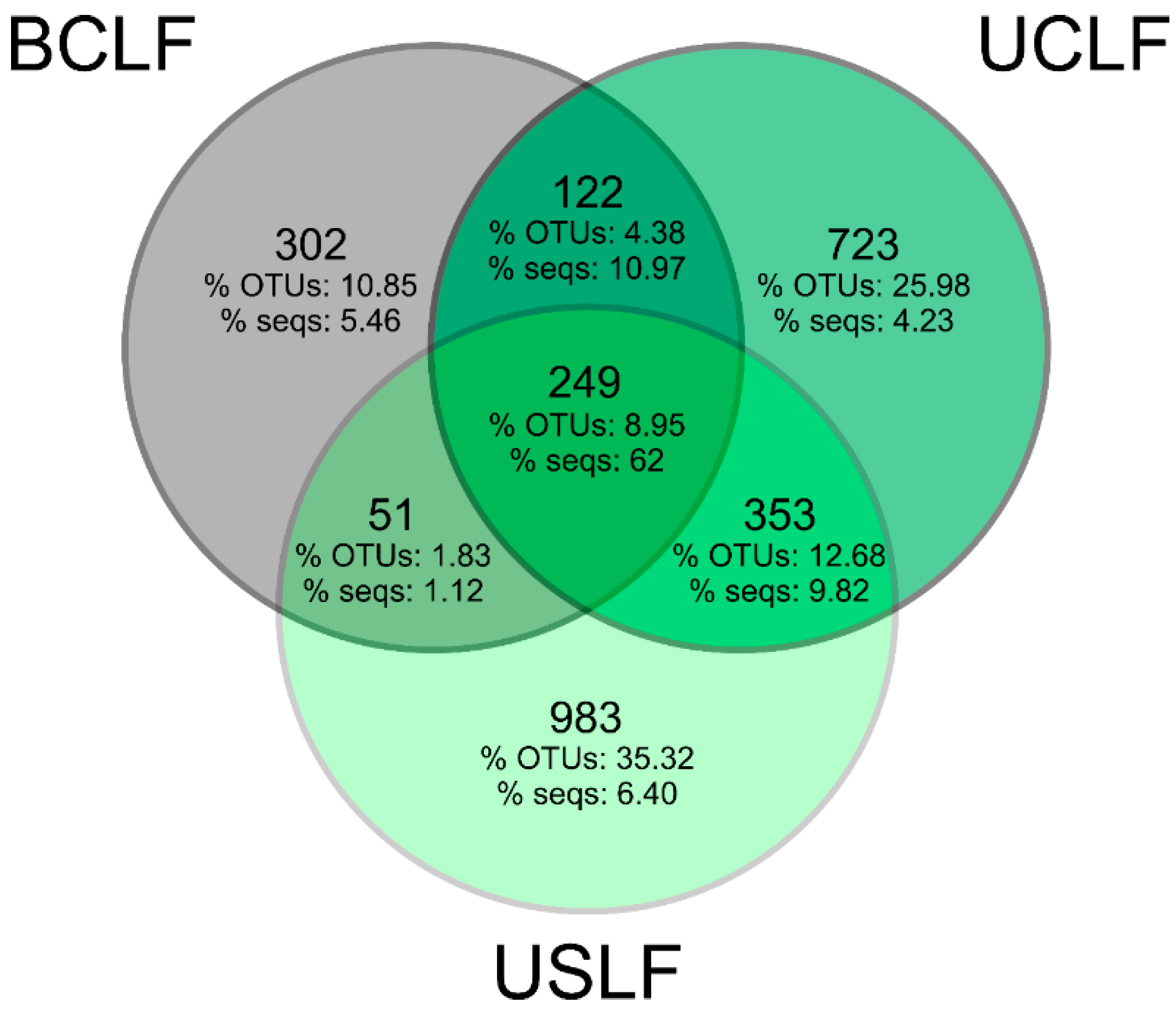
3.6. The Core Microbiome
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Data Availability
References
- Fernández-Palacios, J.M.; de Nascimento, L.; Otto, R.; Delgado, J.D.; García-del-Rey, E.; Arévalo, J.R.; Whittaker, R.J. A reconstruction of Palaeo-Macaronesia with particular reference to the long-term biogeography of the Atlantic island laurel forest. J. Biogeogr. 2011, 38, 226–246. [Google Scholar] [CrossRef]
- Del Arco, M.J.; Wilpret, T.W.; Pérez de Paz, P.L.; Rodríguez-Delgado, O.; Acebes-Ginovés, J.R.; García-Gallo, A.; Martín-Osorio, V.E.; Reyes-Bentancort, A.; Cabrera-Lacalzada, M.V.; García-Avila, S. Mapa de Vegetación de Canarias; GRAFCAN: Tenerife, Spain, 2006. [Google Scholar]
- Notario, J.; Rodríguez, A.R.; Arbelo, A.D.; Rodríguez, A.R.; Fernández, A.; Gómez, L. Burned soils at La Gomera wildfire: A preliminary GIS analysis. Flamma 2015, 6, 95–97. [Google Scholar]
- Certini, G. Effects of fire on properties of forest soils: A review. Oecologia 2005, 143, 1–10. [Google Scholar] [CrossRef]
- Thomaz, E.L.; Antoneli, V.; Doerr, S.H. Effects of fire on the physicochemical properties of soil in a slash-and burn agriculture. Catena 2014, 122, 209–215. [Google Scholar] [CrossRef]
- Hobley, E.U.; Le Gay-Bereton, A.J.; Wilson, B. Forest burning affects quality and quantity of soil organic matter. Sci. Total Environ. 2017, 575, 41–49. [Google Scholar] [CrossRef] [PubMed]
- Aznar, J.M.; González, J.A.; Badía, D.; Martí, C. At what depth are the properties of gypseous forest soil affected by fire? Land Degrad. Dev. 2013, 27, 1344–1353. [Google Scholar] [CrossRef]
- Fritze, H.; Pennanen, T.; Pietikainen, J. Recovery of soil microbial biomass & activity from prescribed burning. Can. J. Res. 1993, 23, 1286–1290. [Google Scholar]
- Acea, M.J.; Carballas, T. Changes in physiological groups of microorganisms in soil following wildfire. FEMS Microbiol. Ecol. 1996, 20, 33–39. [Google Scholar] [CrossRef]
- Hernández, T.; García, C.; Reinhardt, I. Short-term effect of wildfire on the chemical, biochemical & microbiological properties of Mediterranean pine forest soils. Biol. Fertil. Soils 1997, 25, 109–116. [Google Scholar]
- Yeager, C.M.; Kornosky, J.L.; Housman, D.C.; Grote, E.E.; Belnap, J.; Kusk, C.R. Diazotrophic Community structure & function in two successional stages of biological soil crusts from the Colorado Plateau & Chihuahuan desert. Appl. Environ. Microbiol. 2004, 70, 973–983. [Google Scholar]
- Yeager, C.M.; Northup, D.E.; Grow, C.C.; Barns, S.M.; Kuske, C.R. Changes in nitrogen-fixing and ammonia-oxidizing bacterial communities in soil of a mixed conifer forest after wildfire. Appl. Environ. Microbiol. 2005, 71, 2713–2722. [Google Scholar] [CrossRef] [PubMed]
- Smith, N.R.; Kishchuk, B.E.; Mohn, W. Effects of wildfire and harvest disturbances on forest soil bacterial communities. Appl. Environ. Microbiol. 2008, 74, 216–224. [Google Scholar] [CrossRef] [PubMed]
- Covington, W.W.; Sackett, S.S. Soil mineral nitrogen changes following prescribed burning in ponderosa pine. Ecol. Manag. 1992, 54, 175–191. [Google Scholar] [CrossRef]
- Kovacic, D.A.; Swift, D.M.; Ellis, J.E.; Hakonson, T.E. Immediate effects of prescribed burning on mineral soil nitrogen in ponderosa pine of New Mexico. Soil Sci. 1986, 141, 71–76. [Google Scholar] [CrossRef]
- Hobbs, N.T.; Schimel, D.S. Fire Effects on Nitrogen Mineralization and Fixation in Mountain Shrub and Grassl and Communities. J. Range Manag. 1984, 37, 402–406. [Google Scholar] [CrossRef]
- Isobe, K.; Otsuka, S.; Sudiana, I.; Nurkanto, A.; Senoo, K. Community composition of soil bacteria nearly a decade after a fire in a tropical rainforest in East Kalimantan, Indonesia. J. Gen. Appl. Microbiol. 2009, 55, 329–337. [Google Scholar] [CrossRef]
- Xiang, X.; Shi, Y.; Yang, J.; Kong, J.; Lin, X.; Zhang, H.; Zeng, J.; Chu, H. Rapid recovery of soil bacterial communities after wildfire in a Chinese boreal forest. Sci. Rep. 2014, 4, 3829. [Google Scholar] [CrossRef]
- Cobo-Díaz, J.F.; Fernández-González, A.J.; Villadas, P.J.; Robles, A.B.; Toro, N.; Fernández-López, M. Metagenomic Assessment of the potential microbial nitrogen pathways of a Mediterranean forest after a wildfire. Microb. Ecol. 2015, 69, 895–904. [Google Scholar] [CrossRef]
- Shen, J.; Chen, C.; Lewis, T. Long term repeated fire disturbance alters soil bacterial diversity but not abundance in an Australian wet sclerophyll forest. Sci. Rep. 2016, 6, 19639. [Google Scholar] [CrossRef]
- Fernández-González, A.J.; Martínez-Hidalgo, P.; Cobo-Díaz, J.F.; Villadas, P.J.; Martínez-Molina, E.; Toro, N.; Tringe, S.G.; Fernández-López, M. The rhizosphere microbiome of burned holm-oak: Potential role of genus Arthrobacter in the recovery of burned soils. Sci. Rep. 2017, 7, 6008. [Google Scholar] [CrossRef]
- Neris, J.; Tejedor, M.; Rodríguez, M.; Fuentes, J.; Jiménez, C. Effects of forest floor characteristics on water repellency, infiltration, runoff and soil loss in Andisols of Tenerife (Canary Islands, Spain). Catena 2013, 108, 50–57. [Google Scholar] [CrossRef]
- Dorta-Almenar, I.; Navarro, F.; Arbelo, C.; Rodríguez, A.; Notario, J. The temporal distribution of water-soluble nutrients from high mountain soils following a wildfire within legume scrubland of Tenerife, Canary Islands, Spain. Catena 2015, 135, 393–400. [Google Scholar] [CrossRef]
- Durán, J.; Gallardo-Cano, A.; Fernández-Palacios, J.M. Efecto del fuego sobre la biomasa microbiana del suelo de ecosistemas de Pino canario (Pinus canariensis). Cuad. Soc. Esp. Sci. 2004, 20, 181–186. [Google Scholar]
- Rodríguez, J.; Turmero, A.; Blánquez, A.; González-Pérez, J.A.; González-Vila, F.J.; Rodríguez Rodríguez, A.; Pérez Leblic, M.I.; Arias Fernández, M.E. Effect of wildfires on the genetic microbial diversity in forest soils from Canary Islands (Spain). Flamma 2014, 5, 42–44. [Google Scholar]
- Nelson, D.W.; Sommers, L.E. Total carbon, organic carbon, and organic matter. In Methods of Soil Analysis. Part 2. Chemical and Microbiological Properties; Page, A.L., Ed.; SSSA, ASA: Madison, WI, USA, 1982; pp. 539–597. [Google Scholar]
- Olsen, S.; Cole, C.; Watanabe, F.; Dean, L. Estimation of available Phosphorus. In Soils by Extraction with Sodium Bicarbonate; USDA: Washington, DC, USA, 1954; pp. 1–19. [Google Scholar]
- Murphy, J.; Riley, J.P. A modified single solution method for the determination of phosphate in natural waters. Anal. Chim. Acta 1962, 27, 31–36. [Google Scholar] [CrossRef]
- Watanabe, F.S.; Olsen, S.R. Test of an ascorbic method for determining phosphorus in water & NaHCO3 extracts from soils. Soil Sci. Soc. Am. J. 1965, 29, 677–678. [Google Scholar]
- Gee, G.W.; Bauder, J.W. Particle-size analysis. In Methods of Soil Analysis. Part 1. Physical & Mineralogical Properties; SSSA, ASA: Madison, WI, USA, 1986; pp. 383–409. [Google Scholar]
- Klute, A. Water Retention: Laboratory Methods. In Methods of Soil Analysis. Part 1. Physical and Mineralogical Properties; SSSA, ASA: Madison, WI, USA, 1986; pp. 635–660. [Google Scholar]
- Doerr, S.H. On standardizing the “water drop penetration time” and the “molarity of an ethanol droplet” techniques to classify soil hydrophobicity: A case study using medium textured soils. Earth Surf. Proc. Land 1998, 23, 663–668. [Google Scholar] [CrossRef]
- Bisdom, E.B.; Dekker, L.W.; Schoute, J.F. Water repellency of sieve fractions from sandy soils and relationships with organic material and structure. Geoderma 1993, 56, 105–118. [Google Scholar] [CrossRef]
- Baker, G.C.; Smith, J.J.; Cowan, D.A. Review and re-analysis of domain-specific 16S primers. J. Microbiol. Meth. 2003, 55, 541–555. [Google Scholar] [CrossRef]
- Binladen, J.M.; Bollback, T.P.G.P.; Panitz, F.; Bendixen, C.; Nielsen, R.; Willerslev, E. The use of coded PCR primers enables high-throughput sequencing of multiple homolog amplification products by 454 parallel sequencing. PLoS ONE 2007, 2, e197. [Google Scholar] [CrossRef]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J.; et al. Introducing mothur: Open-source, platform-independent, community-supported software for describing & comparing microbial communities. App. Environ. Microbiol. 2009, 75, 7537–7541. [Google Scholar]
- Calleja-Cervantes, M.E.; Aparicio-Tejo, P.M.; Villadas, P.J.; Irigoyen, I.; Irañeta, J.J.B.; Fernández-González, A.J.; Fernández-López, M.; Menéndez, S. Rational application of treated sewage sludge with urea increases GHG mitigation opportunities in Mediterranean soils. Agric. Ecosyst. Environ. 2017, 238, 114–127. [Google Scholar] [CrossRef]
- Parks, D.H.; Beiko, R.G. Identifying biologically relevant differences between metagenomic communities. Bioinformatics 2010, 26, 715–721. [Google Scholar] [CrossRef] [PubMed]
- DeCáceres, M.; Font, X.; Oliva, F.; Vives, S.; Ginkgo, A. Program for Non-Standard Multivariate Fuzzy Analysis. Adv. Fuzzy Sets Syst. 2007, 2, 41–56. [Google Scholar]
- Ter-Braak, C.J.F.; Smilauer, P. Canoco for Windows Version 4.5; Biometris-Plant Research International: Wageningen, The Netherland, 2002. [Google Scholar]
- Clarke, K.R.; Ainsworth, M. A method of linking multivariate community structure to environmental variables. Mar. Ecol. Prog. Ser. 1993, 92, 205–219. [Google Scholar] [CrossRef]
- Heberle, H.; Meirelles, G.V.; Da Silva, F.R.; Telles, G.P.; Minghim, R. InteractiVenn: A web-based tool for the analysis of sets through Venn diagrams. BMC Bioinform. 2015, 16, 169. [Google Scholar] [CrossRef]
- Rodríguez Rodríguez, A.; Arbelo, C.D.; Notario, J.S.; Mora, J.L.; Guerra, J.A.; Armas, C.M. Los suelos del Parque Nacional de Garajonay. In Parque Nacional de Garajonay. Patrimonio Mundial; Fernández, A.B., Turquesa, S.L., Eds.; O.A.P.N.: Madrid, Spain, 2009; pp. 63–91. [Google Scholar]
- Stoof, C.R.; Wesseling, J.G.; Ritsema, C.J. Effects of fire and ash on soil water retention. Geoderma 2010, 159, 276–285. [Google Scholar] [CrossRef]
- Li, W.; Niu, S.; Liu, X.; Wang, J. Short-term response of the soil bacterial community to differing wildfire severity in Pinus tabulaeformis stands. Sci. Rep. 2019, 9, 1148. [Google Scholar] [CrossRef]
- Giovannini, G.; Lucchesi, S.; Giachetti, M. Effects of heating on some physical and chemical parameters related to soil fertility and plant growth. Soil Sci. 1990, 149, 344–350. [Google Scholar] [CrossRef]
- Knicker, H. How Does Fire Affect the Nature and Stability of Soil Organic Nitrogen and Carbon? A Review. Biogeochemistry 2007, 85, 91–118. [Google Scholar] [CrossRef]
- Ferrenberg, S.; O’Neill, S.P.; Knelman, J.E.; Todd, B.; Duggan, S.; Bradley, D.; Robinson, T.; Schmidt, S.K.; Townsend, A.R.; Williams, M.W.; et al. Changes in assembly processes in soil bacterial communities following a wildfire disturbance. ISME J. 2013, 7, 1102–1111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Emerson, W.W. Water retention, organic carbon and soil texture. Aust. J. Soil Res. 1995, 33, 241–251. [Google Scholar] [CrossRef]
- Rawls, W.J.; Pachepsky, Y.A.; Ritchie, J.C.; Sobecki, T.M.; Bloodworth, H. Effect of soil organic carbon on soil water retention. Geoderma 2003, 116, 61–76. [Google Scholar] [CrossRef]
- Reichert, J.M.; Albuquerque, J.A.; Kaiser, D.R.; Reinert, D.J.; Urach, F.L.; Carlesso, R. Estimation of water retention and availability for Rio Grande do Sul soils. R. Bras. Ci. Solo 2009, 33, 1547–1560. [Google Scholar] [CrossRef] [Green Version]
- Janssen, P.H. Identifying the dominant soil bacterial taxa in libraries of 16S rRNA and 16S rRNA genes. Appl. Environ. Microbiol. 2006, 72, 1719–1728. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kielak, A.; Barreto, C.; Kowalchuk, G.A.; van Veen, J.A.; Kuramae, E. The Ecology of Acidobacteria: Moving beyond Genes and Genomes. Front. Microbiol. 2016, 7, 744. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fierer, N.; Bradford, M.A.; Jackson, R.B. Toward an ecological classification of soil bacteria. Ecology 2007, 88, 1354–1364. [Google Scholar] [CrossRef]
- Jones, R.T.; Robeson, M.S.; Lauber, C.L.; Hamady, M.; Knight, R.; Fierer, N. A comprehensive survey of soil acidobacterial diversity using pyrosequencing & clone library analyses. ISME J. 2009, 3, 442–453. [Google Scholar]
- Faoro, H.; Alves, A.C.; Souza, E.M.; Rigo, L.U.; Cruz, L.M.; Al-Janabi, S.M.; Monteiro, R.A.; Baura, V.A.; Pedrosa, F.O. Influence of soil characteristics on the diversity of Bacteria in the southern Brazilian Atlantic forest. Appl. Environ. Microbiol. 2010, 76, 4744–4749. [Google Scholar] [CrossRef] [Green Version]
- Navarrete, A.; Kuramae, E.E.; de Hollander, M.; Pijl, A.S.; van Veen, J.A.; Tsai, S.M. Acidobacterial community responses to agricultural management of soybean in Amazon forest soils. FEMS Microbiol. Ecol. 2013, 83, 607–621. [Google Scholar] [CrossRef] [Green Version]
- Sile, J.A.; Margesin, R. Abundance and Diversity of Bacterial, Archaeal, and Fungal communities Along an Altitudinal Gradient in Alpine Forest Soils: What Are the Driving Factors? Microb. Ecol. 2016, 72, 207–220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lauber, C.L.; Hamady, M.; Knight, R.; Fierer, N. Pyrosequencing-based assessment of soil pH as a predictor of soil bacterial community structure at the continental scale. Appl. Environ. Microbiol. 2009, 75, 5111–5120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Männistö, M.K.; Tiirola, M.; Haggblom, M.M. Bacterial communities in Arctic fields of Finnish Lapland are stable but highly pH-dependent. FEMS Microbiol. Ecol. 2007, 59, 452–465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sait, M.; Davis, K.E.; Janssen, P.H. Effect of pH on isolation and distribution of members of subdivision 1 of the phylum Acidobacteria occurring in soil. Appl. Environ. Microbiol. 2006, 72, 1852–1857. [Google Scholar] [CrossRef] [Green Version]
- Nacke, H.; Thürmer, A.; Wollherr, A.; Will, C.; Hodac, L.; Herold, N.; Schöning, I.; Schrumpf, M.; Daniel, R. Pyrosequencing-Based assessment of bacterial community structure along different management types in German forest and grassland soils. PLoS ONE 2011, 6, e17000. [Google Scholar] [CrossRef] [Green Version]
- Yun, Y.; Wang, H.; Man, B.; Xiang, X.; Zhou, J.; Qiu, X.; Duan, Y.; Engel, A.S. The Relationship between pH and bacterial communities in a single Karst ecosystem & its implication for soil acidification. Front. Microbiol. 2016, 7, 1955. [Google Scholar]
- Weber, C.F.; Lockhart, J.S.; Charaska, E.; Aho, K.; Lose, K.A. Bacterial composition of soils in ponderosa pine and mixed conifer forests exposed to different wildfire burn severity. Soil Biol. Biochem. 2014, 69, 242–250. [Google Scholar] [CrossRef]
- Vila-Escalé, M.; Vegas-Vilarrúbia, T.; Prat, N. Release of polycyclic aromatic compounds into a Mediterranean creek (Catalonia, NE Spain) after a forest fire. Water Res. 2007, 41, 2171–2179. [Google Scholar] [CrossRef]
- Bodour, A.A.; Wang, J.M.; Brusseau, M.L.; Maier, R.M. Temporal change in culturable phenanthrene degraders in response to long-term exposure to phenanthrene in a soil column system. Environ. Microbiol. 2003, 5, 888–895. [Google Scholar] [CrossRef]
- Wang, H.; Lou, J.; Gu, H.; Luo, X.; Yang, L.; Wu, L.; Liu, Y.; Wu, J.; Xu, J. Efficient biodegradation of phenanthrene by a novel strain Massilia sp. WF1 isolated from a PAH-contaminated soil. Environ. Sci. Pollut. Res. Int. 2016, 23, 1378–1388. [Google Scholar] [CrossRef]
- Ofek, M.; Hadar, Y.; Minz, D. Ecology of Root Colonizing Massilia (Oxalobacteraceae). PLoS ONE 2012, 7, e40117. [Google Scholar] [CrossRef] [PubMed]
- Willumsen, P.A.; Johansen, J.E.; Karlson, U.; Hansen, B.M. Isolation & taxonomic affiliation of N-heterocyclic aromatic hydrocarbon-transforming bacteria. Appl. Microbiol. Biotechnol. 2005, 67, 420–428. [Google Scholar] [PubMed]
- Kwon, S.W.; Son, J.A.; Kim, S.J.; Kim, Y.S.; Park, I.C.; Bok, J.I.; Weon, H.Y. Pedobacter rhizosphaerae sp. nov. and Pedobacter soli sp. nov., isolated from rhizosphere soil of Chinese cabbage (Brassica campestris). Int. J. Syst. Evol. Microbiol. 2011, 61, 2874–2879. [Google Scholar] [CrossRef]
- Yang, D.J.; Hong, J.K. Pedobacter solisilvae sp. nov., isolated from forest soil. Int. J. Syst. Evol. Microbiol. 2017, 67, 4814–4819. [Google Scholar] [CrossRef] [PubMed]
- Seldin, L. Paenibacillus, Nitrogen Fixation and Soil Fertility. In Endospore-Forming Soil Bacteria; Logan, N., Vos, P., Eds.; Springer: Berlin/Heidelberg, Germany, 2011; pp. 287–307. [Google Scholar]
- Wang, L.Y.; Chen, S.F.; Wang, L.; Zhou, Y.G.; Liu, H.C. Cohnella plantaginis sp. nov., a novel nitrogen-fixing species isolated from plantain rhizosphere soil. Anton. Leeuw. 2012, 102, 83–89. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.Y.; Wang, T.S.; Chen, S.F. Cohnella capsici sp. nov., a novel nitrogen-fixing species isolated from Capsicum annuum rhizosphere soil, and emended description of Cohnella plantaginis. Anton. Leeuw. 2015, 107, 133–139. [Google Scholar] [CrossRef]
- Pizarro-Tobías, P.; Fernández, M.; Niqui, J.L.; Solano, J.; Duque, E.; Ramos, J.L.; Roca, A. Restoration of Mediterranean forest after a fire: Bioremediation and rhizoremediation field-scale trial. Microb. Biotecnol. 2014, 8, 77–92. [Google Scholar] [CrossRef]
- Sellstedt, A.; Richau, K.H. Aspects of nitrogen-fixing Actinobacteria, in particular free-living and symbiotic Frankia. FEMS Microbiol. Lett. 2013, 342, 179–186. [Google Scholar] [CrossRef] [Green Version]
- Westerberg, K.; Elvang, A.M.; Stackebrandt, E.; Jansson, J.K. Arthrobacter chlorophenolicus sp. nov., a new species capable of degrading high concentrations of 4-chlorophenol. Int. J. Syst. Evol. Microbiol. 2000, 50, 2083–2092. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Soil Sample | |||
|---|---|---|---|
| Parameter | USLF | UCLF | BCLF a |
| SOC (g kg−1) | 105 | 155 | 83 (±13.26) |
| Total N (g kg−1) | 6.6 | 10.5 | 8.7 (±0.09) |
| C/N | 15.9 | 14.8 | 9.5 (±3.02) |
| pH (H2O) | 5.8 | 5.8 | 6.3 (±0.75) |
| Available P (mg kg−1) | 13.7 | 15.0 | 16.0 (±3.25) |
| Available K (cmolckg−1) | 0.5 | 0.8 | 1.5 (±0.4) |
| Available Ca (cmolckg−1) | 5.9 | 9.4 | 10.6 (±6.48) |
| Available Mg (cmolckg−1) | 2.0 | 4.4 | 2.2 (±0.22) |
| Clay (%) | 15.8 | 10.4 | 11.1 (±3.35) |
| Silt (%) | 50.1 | 47.8 | 65.7 (±5.74) |
| Sand (%) | 34.1 | 41.8 | 23.2 (±6.19) |
| Coarse fragments (g kg−1) | 150 | 150 | 233 (±32.3) |
| Texture | Silty loam | Loamy | Silty loam |
| Water retention b | |||
| 33 kPa | 626 | 867 | 403(±56.0) |
| 1500 kPa | 262 | 345 | 236 (±38.0) |
| Available water | 364 | 522 | 167 (±23.8) |
| Water repellency c | |||
| MED | Wettable | Slightly w.r. | Slightly w.r. |
| WDPT | Slightly w.r. | Wettable | Wettable |
| Group | OTUs | Chao | InvSimpson | Simpson | Shannon | Equity | Coverage |
|---|---|---|---|---|---|---|---|
| USLF A | 624.63 | 963.82 | 86.61 | 0.01 | 5.27 | 0.8180 | 0.9388 |
| USLF B | 853.64 | 1482.89 | 152.87 | 0.01 | 5.77 | 0.8551 | 0.9048 |
| USLF C | 958.09 | 1566.26 | 152.51 | 0.01 | 5.90 | 0.8599 | 0.8931 |
| average | 812.12 (170.56) a | 1337.66 (326.43) a | 130.66(38.15) a | 0.01 (0.00) a | 5.65 (0.33) a | 0.8443 (0.0229) a | 0.9122 (0.0237) a |
| UCLF A | 660.23 | 1006.26 | 84.99 | 0.01 | 5.31 | 0.8181 | 0.9335 |
| UCLF B | 704.29 | 1026.83 | 59.37 | 0.02 | 5.29 | 0.8061 | 0.9313 |
| UCLF C | 761.70 | 1076.88 | 96.81 | 0.01 | 5.54 | 0.8356 | 0.9274 |
| average | 708.74 (50.88) a | 1036.66 (36.32) ab | 80.39 (19.14) a | 0.01 (0.01) a | 5.38 (0.14) a | 0.8199 (0.0148) a | 0.9307 (0.0031) ab |
| BCLF A | 182.84 | 241.45 | 13.39 | 0.07 | 3.36 | 0.6442 | 0.9868 |
| BCLF B | 514.64 | 820.26 | 12.14 | 0.08 | 4.17 | 0.6676 | 0.9464 |
| BCLF C | 258.00 | 313.12 | 26.98 | 0.04 | 4.02 | 0.7244 | 0.9831 |
| average | 318.49 (173.98) b | 458.28 (315.53) b | 17.50 (8.23) b | 0.06 (0.02) b | 3.85 (0.43) b | 0.6787 (0.0412) b | 0.9721 (0.0223) b |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Villadas, P.J.; Díaz-Díaz, S.; Rodríguez-Rodríguez, A.; del Arco-Aguilar, M.; Fernández-González, A.J.; Pérez-Yépez, J.; Arbelo, C.; González-Mancebo, J.M.; Fernández-López, M.; León-Barrios, M. The Soil Microbiome of the Laurel Forest in Garajonay National Park (La Gomera, Canary Islands): Comparing Unburned and Burned Habitats after a Wildfire. Forests 2019, 10, 1051. https://doi.org/10.3390/f10121051
Villadas PJ, Díaz-Díaz S, Rodríguez-Rodríguez A, del Arco-Aguilar M, Fernández-González AJ, Pérez-Yépez J, Arbelo C, González-Mancebo JM, Fernández-López M, León-Barrios M. The Soil Microbiome of the Laurel Forest in Garajonay National Park (La Gomera, Canary Islands): Comparing Unburned and Burned Habitats after a Wildfire. Forests. 2019; 10(12):1051. https://doi.org/10.3390/f10121051
Chicago/Turabian StyleVilladas, Pablo J., Sara Díaz-Díaz, Antonio Rodríguez-Rodríguez, Marcelino del Arco-Aguilar, Antonio J. Fernández-González, Juan Pérez-Yépez, Carmen Arbelo, Juana M. González-Mancebo, Manuel Fernández-López, and Milagros León-Barrios. 2019. "The Soil Microbiome of the Laurel Forest in Garajonay National Park (La Gomera, Canary Islands): Comparing Unburned and Burned Habitats after a Wildfire" Forests 10, no. 12: 1051. https://doi.org/10.3390/f10121051
APA StyleVilladas, P. J., Díaz-Díaz, S., Rodríguez-Rodríguez, A., del Arco-Aguilar, M., Fernández-González, A. J., Pérez-Yépez, J., Arbelo, C., González-Mancebo, J. M., Fernández-López, M., & León-Barrios, M. (2019). The Soil Microbiome of the Laurel Forest in Garajonay National Park (La Gomera, Canary Islands): Comparing Unburned and Burned Habitats after a Wildfire. Forests, 10(12), 1051. https://doi.org/10.3390/f10121051