Abstract
Research Highlights: We demonstrate that ectomycorrhizal (ECM) fungi improve plant aluminum (Al)-tolerance in the field and Laccaria bicolor S238A is a promising ECM isolate. Furthermore, we interpret the underlying nutritional mechanism that ECM inoculation facilitates aboveground biomass production as well as nutrients accumulation and translocation. Background and Objectives: Al toxicity is a primary limiting factor for plants growing in acidic soils. Hydroponic/sand culture studies have shown that some ECM fungi could enhance plant Al-tolerance. However, the underlying mechanisms of ECM fungi in improving plant Al-tolerance in the field are still unknown. To fill this knowledge gap, the present study aimed to examine roles of ECM inoculation in biomass production, accumulation and translocation of nutrients and Al in the host plant grown in the field under Al treatment. Materials and Methods: 4-week-old Pinus massoniana seedlings were inoculated with three Laccaria bicolor isolates (L. bicolor 270, L. bicolor S238A or L. bicolor S238N) and grown in an acidic yellow soil under 1.0 mM Al treatment for 12 weeks in the field. Biomass production, accumulation and translocation of P, Ca, Mg, and Al were investigated in these 16-week-old P. massoniana seedlings. Results: All three of these L. bicolor isolates improved biomass production as well as P, Ca and Mg accumulation in P. massoniana seedlings. Moreover, the three ECM isolates facilitated the translocation of P, Ca, and Mg to aboveground in response to Al treatment, particularly when seedlings were inoculated with L. bicolor S238A. In addition, both L. bicolor 270 and L. bicolor S238A had no apparent effects on Al accumulation, while enhanced Al translocation to aboveground. In contrast, L. bicolor S238N decreased Al accumulation but had no significant effect on Al translocation. Conclusions: ECM fungi in the field improved plant Al-resistance by increasing nutrient uptake, and this was mostly due to translocation of P, Ca, and Mg to aboveground, not by decreasing the uptake and translocation of Al.
1. Introduction
Aluminum (Al) toxicity is a primary limiting factor for plants grown in acidic soils, which account for >40% of the world’s arable land [1]. Al toxicity limits plant growth mainly by inhibiting the uptake of nutrient elements, particularly phosphorus (P) and calcium (Ca), and other metabolic processes [2]. The primary phytotoxicity from active Al is the inhibition of root elongation [3], which reduces the root contacting area with soils and nutrients [4]. In addition, Al3+ reduces the amounts of available nutrients, especially P, in the soil solution by combining with PO43−, HPO42−, and H2PO4− to form insoluble AlPO4 [2]. Moreover, Al3+ competes for the membrane binding sites with divalent cations such as Ca2+ and Mg2+ [5], which reduces their uptakes [3] while increasing Al3+ uptake by plants from soil [6].
Ectomycorrhizal (ECM) fungi can protect its host against injury in soils contaminated with metals including Al, Cu, Ni, Pb, Mn, Cd, and Cr [7]. Previous studies have reported that ECM fungi can improve Al tolerance in their host plants. For example, three Pisolithus ecotypes improved Al tolerance in Eucalyptus [8], Pisolithus tinctorius alleviated Al toxicity in Pinus densiflora [9], and Paxillus involutus significantly enhanced the growth of Populus deltoides and Picea abies under Al stress [10,11]. These results indicated that some plant species could be inoculated with ECM fungi to alleviate Al toxicity when grown in acidic soils [12]. ECM fungi could improve plant Al resistance by expanding the scope and amounts of nutrients uptake and/or by immobilizing Al3+ in numerous hyphae and mycelial cords [12,13]. ECM fungi may mobilize nutrients from clays or rocks and make them available by secreting organic acids, protons, and saccharides [14,15]. These secretions can also immobilize Al3+ through chelation as well as Al3+ adsorption by ECM fungi [16,17]. P, which is easily deposited and immobilized by Al3+, may be released by specific excretions such as phosphatases and organic acids from ECM fungi [18].
Our previous experiments had shown that the growth response of Pinus massoniana Lamb. (Masson pine) seedlings inoculated with three individual isolates of Laccaria bicolor (Maire) P.D. Orton showed an increase of at least 30% [19], whereas these isolates were all Al sensitive in vitro because their biomass decreased at least 20% in response to 1.0 mM Al stress [20]. This indicated that there might be different mechanisms of Al resistance between in ECM fungi in vitro and in ECM seedlings in the field. According to the increased biomass and uptake of P, Ca, and Mg in ECM seedlings grown in an acidic yellow soil [19], we hypothesize that ECM inoculation can improve Al resistance in its host through facilitating nutrient accumulation and translocation to aboveground portions of the plants. The objectives of the present study were thus to examine Al resistance and the accumulation and translocation to aboveground portions of P, Ca, Mg, and Al in Pinus massoniana seedlings inoculated with three different L. bicolor isolates after planting in the field in an acidic yellow soil under Al treatment. In so doing, we address the following three hypotheses: (1) The response to Al treatment is not similar between ECM fungi in vitro and their symbionts in the field, (2) an ECM fungus inoculation can affect the accumulation, and particularly translocation, of P, Ca, Mg, and Al in Pinus massoniana seedlings, and (3) which isolate can confer the best Al resistance in P. massoniana seedlings among three of these L. bicolor isolates.
2. Materials and Methods
2.1. Site Description
The experimental site is located in the Southwest University Farm (29°48′39″ N, 106°24′54″ E, 255 m above the sea level), 28 km of north Chongqing, China. This region has a subtropical humid monsoon climate, with an annual temperature of 18.9 °C, annual rainfall of 1152 mm, and monthly mean warmest and coldest temperatures of 28.6 °C (July–August) and 8.7 °C (January–February), respectively. The yellow soil (Haplic Alisols, ST, pH 4.64) under a Pinus massoniana stand on Jigong Mountain (29°48′54″ N, 106°26′36″ E, 534 m above sea level) in Chongqing, China, has developed from sandstones formed during the Jurassic Period. The basic macronutrient and Al concentrations of this soil at 0–20 cm depth are: 0.99 g total N kg−1, 0.35 g total P kg−1, 21.68 g total K kg−1, 72.95 mg available N kg−1, 3.93 mg available P (extracted by 0.05 M HCl and 0.025 M H2SO4) kg−1, 62.71 mg available K kg−1, 46.82 g total Al kg−1, and 1.23 g labile Al kg−1 [19]. Soils were sterilized with 2% formaldehyde prior to sowing the germinated pine seeds and were then loaded into concrete chambers (2.0 m × 6.0 m × 0.5 m = length × width × depth) at the experimental field site.
2.2. Fungal Isolates and Production of ECM Plant Seedlings
Three Laccaria bicolor isolates (L. bicolor 270, L. bicolor S238A, and L. bicolor S238N) were used in this experiment. L. bicolor 270 was isolated from a Pseudotsuga menziesii forest in Canada, while both L. bicolor S238A and L. bicolor S238N were originally obtained from a Pinus taeda stand in France. Four agar mycelia discs (3 × 3 mm) for inoculation were sub-cultured in each 250 mL flask with 50 mL Pachlewski liquid medium (pH 5.5) for 21 d at 25 °C in the dark in the Silviculture Lab at Southwest University, Chongqing, China.
After 21 days of subculture and removal of the growth medium, the fungal mycelia were disrupted in a blender (Philips HR 2024, Philips Inco., Japan) for 1 min. A total of 200 germinating Pinus massoniana seeds inoculated with five flasks of mixed fungal mycelia were sown on 1.0 m2 of sterilized soil (0.5 cm depth) to develop ECM fungal associations (+ECM). The control seedlings (–ECM) were not inoculated with fungal mycelia. Prior to the fungal culture, uniform seeds of P. massoniana from the Chongqing Tree Seeds and Seedlings Service, Chongqing, China were sterilized with 0.3% formaldehyde solution for 30 min, rinsed with sterilized water, and then germinated on moistened paper towels at 25 °C in the dark.
2.3. Aluminum Treatment
After four weeks of growth, the seedlings were watered weekly with Hoagland solution (pH 3.8) that contained 5 mM Ca(NO3)2·4H2O, 5 mM∙KNO3, 2 mM MgSO4·7H2O, 1 mM KH2PO4, 0.02 mM FeC4H4O6, and either 0.0 (−Al) or 1.0 mM Al3+ (+Al) solution as Al2(SO4)3·18H2O for 12 weeks. The total Al added to the soil was 81 mg kg−1. Seedlings with or without mycelia inoculation or Al treatment in a 1.0 m2 plot were randomly arranged in three replicates. The total number of 1.0 m2 plots was 24 (4 ECM inoculations × 2 Al-treatments × 3 replicates).
2.4. Sampling and Analysis
After 12 weeks of Al-treatment, 25 of the 16-week-old seedlings in each plot were selectively harvested, and the percentage of roots showing ECM fungus colonization were examined using a dissecting microscope (XSZ-Hs7, Chongqing Photoelectric Instrument Co. LTD, China). Root ECM colonization ranged from 95% to 100% in +ECM seedlings, but no obvious root colonization was observed in –ECM seedlings. All seedlings were washed with deionized water, subdivided into belowground and aboveground parts, oven-dried at 70 °C for 48 h, weighed, ground using a commercially-available cereal mill grinder (BJ-150, Baijie Inc., China) at 25,000 r min−1 for 1 min, and digested in a nitric acid/perchloric acid solution (HNO3/HClO4 = 5:1, v/v) for P, Ca, Mg, and Al analyses [21]. The accumulation of P, Ca, Mg, or Al was calculated as the concentration of each ion × the amount of biomass.
The rhizosphere soil, collected after shaking the plant roots, was ground using a wooden rolling pin and sieved through a 1-mm sieve after the removal of litter and debris, and then oven-dried at 110 °C. Following Bao’s method [21], soil pH (1:2.5 = soil:water) was measured with a pH meter (pH-3C, Leici Co., China), soil available P, exchangeable Al, exchangeable Ca, and exchangeable Mg were determined after extraction with 0.05 M HCl and 0.025 M H2SO4, 1M KCl, and 1M CH3COONH4 (pH 7.0), respectively. All extraction procedures were performed in three replicates, and the concentrations of P, Al, Ca, and Mg were measured spectrometrically by the Mo-Sb (P), aluminon (Al), and atomic absorption (Ca and Mg) methods [21].
The shoot/root ratio, an indicator to evaluate the capacity of a plant to allocate carbon (C) between shoots and roots [22], was calculated as the ratio of aboveground to belowground biomass. The translocation factor, an indicator to evaluate the capacity of a plant to transport elements from the belowground to aboveground parts, was calculated as the ratio of aboveground to belowground element accumulation as described by Yuan et al. [23].
2.5. Statistical Analysis
Using the SPSS software version 21.0 (SPSS Inc., Chicago, IL, USA), the normal distribution and homogeneity variances of data (means ± SD, n = 3) were tested using the Shapiro–Wilk test and Levene’s test, respectively. The data were then analyzed using a two-way ANOVA to test the treatment (ECM inoculation, Al treatment, and ECM inoculation × Al treatment) effects on the plant biomass production, accumulation and translocation to aboveground portions of P, Ca, Mg, and Al. Significant differences among treatments were compared with Fisher’s LSD test at p < 0.05. The relationships between the biomass production and accumulations of P, Ca, Mg, and Al were assessed by linear regression analyses. Graphs were produced using GraphPad Prism version 5.01 (GraphPad Software, Inc., San Diego, CA, USA).
3. Results
3.1. Biomass of P. massoniana Seedlings
The biomass production in 16-week-old P. massoniana seedlings was similar between the two Al treatments, and was significantly higher in +ECM seedlings than in –ECM counterparts (p < 0.05, Table 1 and Figure 1). In addition, there was a significant interaction for the biomass production between the Al treatments and ECM inoculations (p < 0.05). In the −Al treatment, the biomass production increased more (~74%) in both the L. bicolor 270 and L. bicolor S238N inoculations than in the L. bicolor S238A inoculation (49%). On the contrary, the increase of plant biomass in the +Al treatment was higher (69%) under the L. bicolor S238A inoculation than under both the L. bicolor 270 and L. bicolor S238N inoculations (26% and 37%, respectively). In addition, the biomass production in the +Al treatment was significantly higher than in the −Al treatment for the L. bicolor S238A-inoculated seedlings, but was lower for both the L. bicolor 270 and L. bicolor S238N-inoculated seedlings.

Table 1.
Two-way ANOVA results for plant biomass production, accumulation and translocation factor of P, Ca, Mg, and Al in 16-week-old Pinus massoniana Lamb. seedlings grown in an acidic yellow soil, subjected to ectomycorrhizal inoculation and Al treatment.
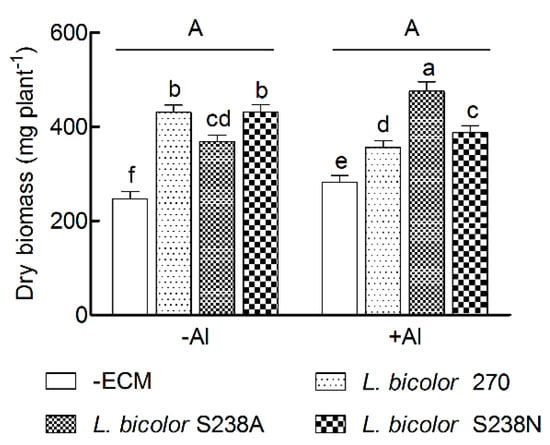
Figure 1.
Biomass production in 16-week-old Pinus massoniana Lamb. seedlings grown in an acidic yellow soil in response to Al treatment. “−Al” and “+Al” indicated that seedlings were watered once weekly with Hoagland solution (pH 3.8) containing 0.0 mM Al3+ or 1.0 mM Al3+, respectively. Data (means ± SD, n = 3) followed by different uppercase or lowercase letters indicate significant differences between Al treatments or between ectomycorrhizal treatments compared with Fisher’s LSD test at p < 0.05 level.
3.2. Accumulation of P, Ca, Mg, and Al in P. massoniana Seedlings
The +Al treatment significantly increased the accumulation of P, Mg, and Al in 16-week-old P. massoniana seedlings by 27%, 8%, and 11%, respectively, compared to the −Al treatment (p < 0.05, Table 1 and Figure 2), but had no significant effects on the Ca accumulation (p > 0.05). The accumulations of P, Ca, Mg, and Al were significantly higher in the ECM seedlings than in the –ECM counterparts (p < 0.05). Meanwhile, a significant interaction was observed between the Al treatments and ECM inoculations on the accumulations of P, Ca, Mg, and Al (p < 0.05).
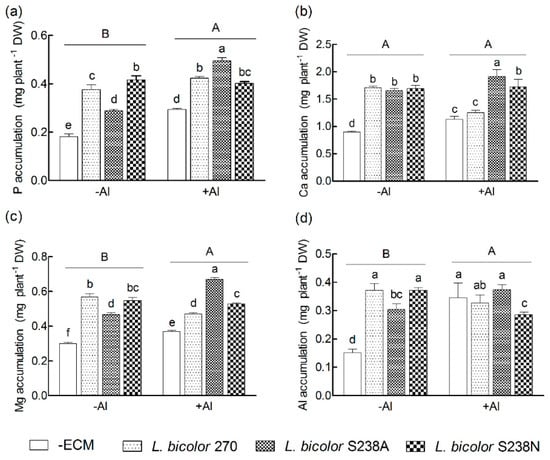
Figure 2.
Accumulation of P (a), Ca (b), Mg (c), and Al (d) in 16-week-old Pinus massoniana Lamb. seedlings grown in an acidic yellow soil in the absence (−Al) or the presence (+Al) of 1.0 mM Al3+. Data (means ± SD, n = 3) followed by different uppercase or lowercase letters indicate significant differences between Al treatments or between ectomycorrhizal treatments compared with Fisher’s LSD test at p < 0.05 level.
In the −Al treatment, both the L. bicolor 270 and L. bicolor S238N inoculations had significantly higher increase (107% and 129%, respectively) on the P accumulation than the L. bicolor S238A inoculation (59%), compared to the –ECM inoculation (p < 0.05, Figure 2a). On the contrary, in the +Al treatment the L. bicolor S238A inoculation had higher increase (67%) than both the L. bicolor 270 and L. bicolor S238N inoculations (44% and 37%, respectively). The P accumulation in the +Al treatment was significantly higher than in the −Al treatment for the same ECM inoculated seedlings; the L. bicolor S238N-inoculated seedlings were the exception, in which no significant difference was observed between the two Al treatments.
In the −Al treatment, the Ca accumulation had no obvious differences among three of these ECM seedlings, but was ~88% higher in the ECM than in the –ECM seedlings (p < 0.05, Figure 2b). On the contrary, compared to the –ECM inoculation in the +Al treatment, the Ca accumulation increased
Compared to the –ECM inoculation, both the L. bicolor 270 and L. bicolor S238N inoculations had significantly higher increases (~86%) in Mg accumulation compared to the L. bicolor S238A inoculation (56%) in the −Al treatment, and significantly lower increase (27% and 43%, respectively) than the L. bicolor S238A inoculation (80%) in the +Al treatment (p < 0.05, Figure 2c). The Mg accumulation in the +Al treatment was significantly higher than in the −Al treatment for the L. bicolor S238A-inoculated seedlings, but was lower for the L. bicolor 270-inoculated seedlings. There were no significant differences in the L. bicolor S238N-inoculated seedlings between the two Al treatments.
The effect of ECM inoculations on the Al accumulation varied in response to the Al treatment. In the −Al treatment, the Al accumulation in the ECM seedlings was significantly higher than in the –ECM seedlings, following a higher increase (~144%) in both the L. bicolor 270 and L. bicolor S238N-inoculated seedlings than in the L. bicolor S238A-inoculated seedlings (101%). Conversely, the Al accumulation in the +Al treatment was not affected under both the L. bicolor 270 and L. bicolor S238A inoculations, whilst was significantly decreased by 22% under the L. bicolor S238N inoculation, compared to the –ECM inoculation (p < 0.05, Figure 2d). The Al accumulation in the +Al treatment was significantly higher than in the −Al treatment for the L. bicolor S238A-inoculated seedlings, but was significantly lower for the L. bicolor S238N-inoculated seedlings. There were no significant differences in Al accumulation in the L. bicolor 270-inoculated seedlings between the two Al treatments.
3.3. Shoot/Root Biomass Ratio in P. massoniana Seedlings
The shoot/root ratios in 16-week-old P. massoniana seedlings significantly increased in both the Al treatments and the ECM inoculations (p < 0.05, Table 1 and Figure 3). In addition, there was a significant interaction between the Al treatments and ECM inoculations (p < 0.05). No significant differences were observed between the +ECM and the –ECM seedlings in the −Al treatment, although the shoot/root ratio was significantly higher in the L. bicolor 270-inoculated than in the L. bicolor S238A-inoculated seedlings. Conversely, the ECM inoculation had an evident improvement on the shoot/root ratio in the +Al treatment, with a higher increase (69%) in L. bicolor S238A-inoculated seedlings than in both the L. bicolor 270 and L. bicolor S238N-inoculated seedlings (24% and 39%, respectively). The shoot/root ratio was significantly higher in the +Al treatment than in the −Al treatment for the L. bicolor S238A-inoculated seedlings, while was similar for both the L. bicolor S238N and L. bicolor 270-inoculated seedlings.
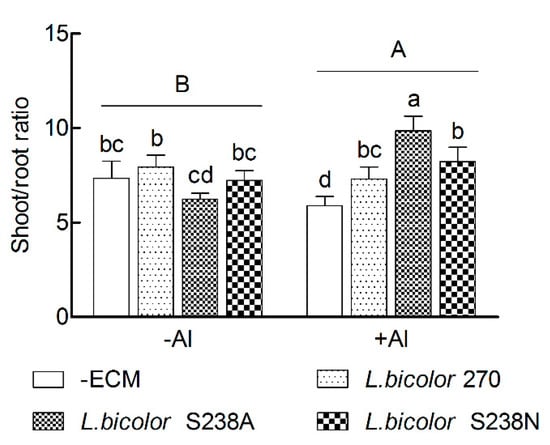
Figure 3.
Shoot/root ratio in 16-week-old Pinus massoniana Lamb. seedlings grown in an acidic yellow soil with (+Al) or without (−Al) added Al. Data (means ± SD, n = 3) followed by different uppercase or lowercase letters indicate significant differences between Al treatments or between ectomycorrhizal treatments compared with Fisher’s LSD test at p < 0.05 level.
3.4. Translocation of P, Ca, Mg, and Al in P. massoniana Seedlings
Translocation factors of P, Ca, Mg, and Al in 16-week-old P. massoniana seedlings were significantly higher in the +Al treatment than in the −Al treatment (p < 0.05, Table 1 and Figure 4). In general, the ECM inoculations significantly increased the translocation factors of P, Ca, and Mg, but decreased the Al translocation factor, compared to the –ECM inoculation (p < 0.05), with an exception of Ca translocation factor under L. bicolor S238A, Mg translocation factor under L. bicolor S238N, and Al translocation factor under both the L. bicolor S238A and L. bicolor 270. In addition, significant interactions existed between the Al treatments and ECM inoculations (p < 0.05).
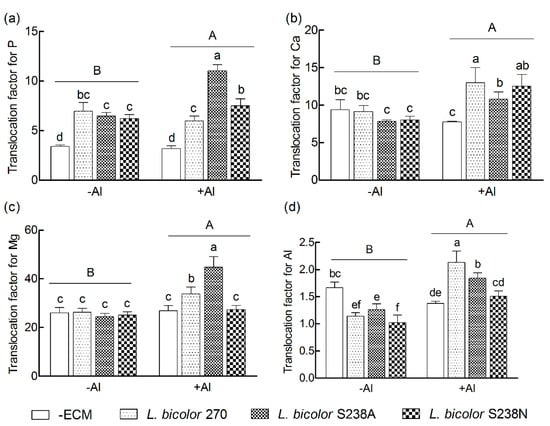
Figure 4.
Translocation factors for P (a), Ca (b), Mg (c), and Al (d) in 16-week-old Pinus massoniana Lamb. seedlings grown in an acidic yellow soil with (+Al) or without (−Al) added Al. Data (means ± SD, n = 3) followed by different uppercase or lowercase letters indicate significant differences between Al treatments or between ectomycorrhizal treatments compared with Fisher’s LSD test at p < 0.05 level.
The P translocation factors were not obviously different among the three ECM seedlings in the −Al treatment, but were significantly ~92% higher in the ECM compared to the –ECM seedlings (p < 0.05, Figure 4a). Conversely, the P translocation factors were significantly higher in the L. bicolor S238A-inoculated seedlings than in the both L. bicolor 270 and L. bicolor S238N-inoculated seedlings in the +Al treatment, with the increase of 248%, 137%, and 89%, respectively, compared to the –ECM seedlings. The P translocation factors were significantly higher in the +Al treatment than in the −Al treatment for both the L. bicolor S238A and L. bicolor S238N-inoculated seedlings, but was similar for the L. bicolor 270-inoculated seedlings.
No significant differences in the Ca translocation factor were observed among the three ECM seedlings and between the ECM and the –ECM seedlings in the −Al treatment. But in the +Al treatment, the ECM inoculation significantly increased the Ca translocation factor compared to the –ECM seedlings and the L. bicolor 270-inoculated seedlings has a significantly higher Ca translocation factor than the L. bicolor S238A-inoculated seedlings (p < 0.05, Figure 4b). The Ca translocation factor was significantly higher in the +Al treatment than in the −Al treatment for the three classes of ECM-inoculated seedlings.
There were no significant differences in the Mg translocation factor among the three ECM seedlings and between the +ECM and –ECM seedlings in the −Al treatment. However, the Mg translocation factor significantly increased in both L. bicolor S238A and L. bicolor 270 inoculations and ranked in the order L. bicolor S238A > L. bicolor 270 in the +Al treatment (p < 0.05, Figure 4c). The Mg translocation factor was significantly higher in the +Al treatment than in the −Al control for both the L. bicolor 270 and L. bicolor S238A-inoculated seedlings, but was similar for the L. bicolor S238N-inoculated seedlings.
The +ECM seedlings had significantly lower Al translocation factors than the –ECM seedlings in the −Al treatment, with significantly higher Al translocation factor in the L. bicolor S238A- than in the L. bicolor S238N-inoculated seedlings (p < 0.05, Figure 4d). On the contrary, compared to the –ECM seedlings, the Al translocation factors significantly increased in both the L. bicolor 270 and L. bicolor S238A inoculations in the +Al treatment and ranked in the order L. bicolor 270 > L. bicolor S238A. The L. bicolor S238N inoculation had no obvious effects on the Al translocation factor in the +Al treatment. The Al translocation factors were significantly higher in the +Al treatment than in the −Al treatment for the pine seedlings inoculated with all three strains of L. bicolor (highest under L. bicolor 270).
3.5. Contributions of P, Ca, Mg, and Al to the Growth in P. massoniana Seedlings
Plant biomass production was significantly positively correlated with the accumulations of P, Ca, Mg, and Al in 16-week-old P. massoniana seedlings in the −Al treatment (R2 = 0.99, p < 0.05, Table 2) and also in the +Al treatment (R2 = 0.99, p < 0.05). With an order Mg (β = 0.61) > P (β = 0.24) > Ca (β = 0.17), the accumulations of P, Ca, and Mg were significant for plant biomass production in the +Al treatment (p < 0.05). In contrast, neither of the accumulations of P, Ca nor Mg were significant for the biomass production in the −Al treatment (p > 0.05).
Table 2.
Relationships between biomass production and accumulations of P, Ca, Mg, and Al in 16-week-old Pinus massoniana Lamb. seedlings grown in an acidic yellow soil with (+Al) or without (−Al) Al treatment (n = 12).
4. Discussion
4.1. Aluminum Resistance and Biomass Production in ECM P. massoniana Seedlings
In this study, all three ECM isolates showed a remarkable ability to improve the growth and Al tolerance of 16-week-old P. massoniana seedlings grown in an acidic yellow soil in the field, even under +Al treatment (Figure 1), which verified that ECM inoculation could improve Al resistance in its host. Similar results were observed for other ECM fungi, including Pisolithus tinctorius and Paxillus involutus, which could improve the Al tolerance of their host plants Eucalyptus [8], Pinus densiflora [9], Populus deltoides [11], and Picea abies [10]. Moreover, Al tolerance conferred by ECM inoculation differed among the ECM isolates, and L. bicolor S238A demonstrated the best Al resistance in P. massoniana seedlings (Figure 1). Seedlings inoculated with L. bicolor S238A were Al resistant, while both L. bicolor 270- and L. bicolor S238N-inoculated seedlings were Al sensitive, although Gu and Huang [20] reported that three of these L. bicolor isolates (270, S238A and S238N) in vitro were sensitive to 1.0 mM Al treatment, and that L. bicolor S238A was the most susceptible isolate. Thompson and Medve [24] found that the responses of three Pisolithus tinctorius isolates to Al in vitro were not consistent with field observations, while Egerton-Warburton [8] observed an agreement between the in vitro Al tolerance of three Pisolithus ecotypes and their performance in symbiosis with Eucalyptus in an acidic mine soil. These results suggested that Al resistance in ECM plants was affected by numerous factors, including ECM fungus species or isolates, host plant species, and growth conditions. Bucking et al. [25] suggested that ectomycorrhizae could alter the function of both the host and fungus, while Costa et al. [26] found that the growth of Eucalyptus urophylla benefited from an association with Pisolithus sp. Our results indicated that the symbiosis benefited the growth and function of both L. bicolor S238A and P. massoniana seedlings, but the underlying mechanisms need further study.
All three of these L. bicolor isolates significantly improved the shoot/root ratios of P. massoniana seedlings in the +Al treatment, but generally not in the −Al treatment (Figure 3), which indicated that ECM inoculations facilitated aboveground biomass production. In general, the Al-tolerant ECM seedlings (L. bicolor S238A-inoculated) had a significantly higher aboveground biomass production than did the Al-sensitive ECM seedlings (L. bicolor 270- and L. bicolor S238N-inoculated) in the +Al treatment (Figure 3). Schier and McQuattie [27] found that shoot growth, but not root growth, was significantly improved in Pisolithus tinctorius-inoculated Pinus strobus seedlings grown in sand across 0–100mg L−1Al concentrations (pH 3.8), while Huang and Lapeyrie [28] observed that L. bicolor S238N had no evident effects on the shoot/root ratio of Pseudotsuga menziesii seedlings in the −Al treatment. Results from the present study agreed with these results of the previous studies. According to Shinde et al. [22], the shoot/root ratio could be used to evaluate the capacity of a plant to allocate carbon between roots and shoot. In their experiment, Populus tremuloides seedlings inoculated with L. bicolor S238N allocated less C to the shoots, while more C was allocated to the roots grown under P limitation. The results of the present study did not agree with their results since ECM inoculation allocated more C to the shoots in the +Al treatment (Figure 3). Choi et al. [29] and Makita et al. [30] have suggested that C allocated to the belowground parts under P limitation is for hyphal growth, fungal respiration, and other fungal metabolic demands, which could greatly increase the nutrient capturing capability of the plant [31]. Although ~20% of the carbon assimilated by the plant was consumed by the fungal symbiont [32], ECM fungi associations might prevent the formation of root hairs and thus inhibit root growth [33,34,35]. This might result in relatively lower C in the belowground parts, and thus more C was allocated aboveground [36].
4.2. Accumulation and Translocation of P, Ca, and Mg in ECM P. massoniana Seedlings
Our experiments showed that L. bicolor-inoculated P. massoniana seedlings accumulated more P, Ca, and Mg when grown in in an acidic yellow soil whether or not they were treated with Al (Figure 2a–c), and the accumulations of Mg, P, and Ca contributed positively to plant biomass production (Table 2). Furthermore, the Al-tolerant ECM seedlings took up more P, Ca, and Mg from the soil than the Al-sensitive ECM seedlings did in the +Al treatment (Figure 2a–c). Various ECM fungi, such as Pisolithus ecotypes, Pisolithus tinctorius, Laccaria bicolor, and Paxillus involutus could improve plant P concentrations [8,37,38]. Considering plant Ca and Mg uptake, the role of ECM fungi differed with respect to fungal species or ecotypes, host plant species, and growth conditions. For example, an Al-tolerant Pisolithus inoculum, Paxillus involutus, and Pisolithus tinctorius (strain Pt KACC) improved Ca and Mg concentration in Eucalyptus growing in acidic mine spoil [8], Populus deltoides in Melin’s liquid medium containing Al [11], and Populus growing in mine tailings [38]. However, Scholl et al. [6] found that ECM colonization had no significant effects on concentrations of Ca and Mg in Pinus sylvestris seedlings grown in a semi-hydroponic system with or without Al treatment, and there was even less Ca in ECM Pinus sylvestris seedlings cultured in perlite [39]. Both Smits et al. [40] and Remiszewski et al. [15] suggested that ECM fungi increased rock weathering and mineral decomposition and thus released more elemental nutrients (Ca, Mg, and P) into the soil. Gu et al. [41] found that L. bicolor-inoculated P. massoniana seedlings mobilized more inorganic P (Fe-P, Ca-P, and Al-P) than did the –ECM seedlings grown in an acidic yellow soil. In our study, the increased accumulation of P, Ca, and Mg in ECM seedlings (Figure 2a–c) demonstrated that ECM fungi could facilitate nutrient release from the soil. The decreased rhizosphere soil pH (Table S1) in the +Al treatment indicated that L. bicolor inoculation might stimulate proton secretion from ECM-inoculated seedlings, thus facilitating the release of nutrients such as P, Ca, and Mg. In addition, the decreased available P, exchangeable Ca, and exchangeable Mg (Table S1) in rhizosphere soils in the +Al treatment suggested that ECM inoculation facilitated the uptake of available nutrients by plants. However, additional work is needed to understand how ECM inoculation mediates the movement of nutrients from the soil to the plant.
Results from our study showed that ECM inoculation facilitated P translocation, reduced Al translocation, and had no significant effects on Ca and Mg translocation from the roots to the shoot in the −Al treatment (Figure 4a–c). Moreover, there were higher translocation factors for P, Ca, and Mg in the ECM seedlings under +Al than under −Al treatment (Figure 4a–c). Laccaria bicolor S238A showed the most promising contribution to translocation of P and Mg in P. massoniana seedlings in the +Al treatment. Numerous studies had shown that ECM fungi caused plants to take up more nutrients and Al [20,42,43]. Increased P and Al uptake also occurred in Pisolithus tinctorius-colonized seedlings of Pinus strobus [27] and Hebeloma crustuliniforme-colonized Picea abies seedlings [44]. In this study, L. bicolor inoculation improved the accumulation of P, Ca, Mg, and Al in P. massoniana seedlings in the −Al treatment (Figure 2a–d). Aluminum may combine with PO43−, HPO42−, H2PO4- to form insoluble AlPO4 not only in the soil solution but also in the root apoplast [2,4]. Also, Al3+ could replace Ca2+ and Mg2+ in the root cortex and compete for their binding sites on the membrane [5]. Thus, Al might prevent P, Ca, and Mg from being transported upward to the aboveground parts of the plant [37]. However, our findings suggested that the transport route for P, Ca, and Mg might differ from that of Al in the ECM seedlings, or that the ECM seedlings had more specific P transporters [18,37], and that ECM inoculation ameliorated the negative effects of Al toxicity on nutrient transportation in P. massoniana seedlings and improved the transport of P, Ca, and Mg in seedlings even in the +Al treatment.
4.3. Accumulation and Translocation of Al in ECM P. massoniana Seedlings
Ectomycorrhizae could ameliorate metal toxicity, partly because these fungi could prevent plants from absorbing metals from the soil [8,11,45,46]. Results from our study provided partial support for this statement (Figure 2d). Only L. bicolor S238N decreased Al uptake, whilst both L. bicolor 270 and L. bicolor S238A had no significant effects on plant Al accumulation in the +Al treatment. In contrast, all three isolates improved Al accumulation in the −Al treatment (Figure 2d). These results suggested that inoculation with L. bicolor S238N could reduce Al uptake and thus improved Al tolerance in the host in the +Al treatment. Tang et al. [47] found that inoculation with Cenococcum geophilum, Pisolithus tinctorius, Laccaria laccata, or Hebeloma vinosophyllus accelerated the absorption of metals (Cu and Cd) by Pinus thunbergii. Schier et al. [27], Tang et al. [47] and Aguirre et al. [48] considered that ectomycorrhizae could ameliorate metal toxicity, which mainly resulted from enhanced nutrient uptake and growth promotion, rather than a decrease in metal ion uptake. Based on seedling growth (Figure 1) and accumulation of P, Ca, and Mg in seedlings (Figure 2a–c), both L. bicolor S238A and L. bicolor 270 might use a similar pathway to increase nutrient uptake but not decrease Al uptake, thus improving Al tolerance in P. massoniana seedlings. In addition, we found that L. bicolor S238N could decrease Al uptake in the +Al treatment and enhance nutrient uptake.
Previous studies suggested that ECM plants could prevent upward transport of metals, thus ameliorating metal toxicity [49,50]. Our experimental data (Figure 4d) supported this in P. massoniana seedlings grown in the −Al treatment. All three of these ECM isolates decreased the translocation of Al from roots to the shoot in the −Al treatment (Figure 4d). Moyer-Henry et al. [51] observed that large amounts of Al accumulated at the fungal mantle, in areas with the Hartig net, and inside hyphae of Pisolithus tinctorius colonizing Pinus taeda seedlings. Vaario et al. [52] found that mushrooms of Tricholoma matsutake, and ECM fungus, were enriched in Al. Pisolithus tinctorius increased Cu accumulation in Pinus tabulaeformis and the majority of the Cu remained in the roots [53]. These results indicated that ECM fungi might limit Al3+ movement toward the inner root tissues and then inhibit Al3+ translocation to the shoot. Gu et al. [54] demonstrated that Al was mainly distributed in the roots, while this study found a decreased Al in the shoots of ECM P. massoniana seedlings in the −Al treatment (Figure 4d). Nevertheless, the Al translocation factor increased in seedlings that were inoculated with both L. bicolor 270 and L. bicolor S238A in the +Al treatment (Figure 4d). Similar results were found as a high root-to-shoot translocation rate for both Zn and Mn in metal-tolerant birch growing in heavy-metal polluted soils [50]. According to Liu and Liu [55], P. massoniana is a moderately Al-resistant plant and can be stimulated by <0.15 mM Al; L. bicolor inoculation may improve its capacity to adapt to higher concentrations of Al in the soil, or increase the threshold of active Al that the host is able to tolerate [51]. Nevertheless, further field-based studies are needed to determine the mechanisms that control Al transport in ECM seedlings under Al stress.
5. Conclusions
The three tested ECM isolates (L. bicolor 270, L. bicolor S238A and L. bicolor S238N), particularly L. bicolor S238A, improved Al tolerance in 16-week-old P. massoniana seedlings in the field in response to Al exposure. All three of these ECM isolates were Al-sensitive in vitro, but L. bicolor S238A became Al resistant when associated with P. massoniana seedlings. The ECM inoculation improved seedling growth and the accumulation of P, Ca, and Mg. Furthermore, the ECM fungi facilitated the translocation of P, Ca, and Mg to the aboveground parts of the seedlings in the +Al treatment, particularly when the seedlings had been inoculated with L. bicolor S238A. In addition, only L. bicolor S238N reduced Al accumulation, while both L. bicolor 270 and L. bicolor S238A increased Al translocation. Therefore, the ECM inoculation improved Al tolerance of P. massoniana seedlings in the field in response to Al exposure mainly by increasing nutrient uptake, and particularly by facilitating nutrient translocation to the aboveground parts of the plant rather than by decreasing the uptake and/or translocation of Al. Further research should focus on the transport processes and transporters of nutrients and Al from the soil to the plant or in ECM seedlings exposed to Al in the field.
Supplementary Materials
The following are available online at https://www.mdpi.com/1999-4907/10/12/1153/s1, Table S1: Rhizosphere soil pH, available P, exchangeable Ca, exchangeable Mg, and exchangeable Al of 16-week-old Pinus massoniana seedlings grown in acidic yellow soil with (+Al) or without (−Al) Al treatment.
Author Contributions
Conceptualization, X.G.; investigation, X.W. and J.L.; writing—original draft, X.G.; writing—review and editing, X.H.; funding acquisition, X.G. and X.H. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Natural Science Foundation of China (31570599 and 31100464) and Biological Science Research Center at the Southwest University, China.
Acknowledgments
The author thanks Jianguo Huang and Ling Yuan for supplying the ECM isolates. We would like to thank the native English-speaking scientists of Elixigen Company (Huntington Beach, California) for editing our manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Moustaka, J.; Ouzounidou, G.; Baycu, G.; Moustakas, M. Aluminum resistance in wheat involves maintenance of leaf Ca2+ and Mg2+ content, decreased lipid peroxidation and Al accumulation, and low photosystem II excitation pressure. BioMetals 2016, 29, 611–623. [Google Scholar] [CrossRef]
- Han, G.; Deng, X.; Jiang, C.; Gu, X. The research progress on the cause and control of aluminum toxicity to plants. J. Fujian For. Sci. Technol. 2007, 34, 174–179. [Google Scholar]
- Hirano, Y.; Isomura, A.; Kaneko, S. Root morphology and nutritional status of Japanese red cedar saplings subjected to in situ levels of aluminum in forest soil solution. J. For. Res. 2003, 8, 209–214. [Google Scholar] [CrossRef]
- Poschenrieder, C.; Gunsé, B.; Corrales, I.; Barceló, J. A glance into aluminum toxicity and resistance in plants. Sci. Total Environ. 2008, 400, 356–368. [Google Scholar] [CrossRef]
- Godbold, D.L.; Fritz, E.; Hüttermann, A. Aluminum toxicity and forest decline. Proc. Natl. Acad. Sci. USA 1988, 85, 3888–3892. [Google Scholar] [CrossRef]
- Scholl, L.; Keltjens, W.G.; Hoffland, E.; Breemen, N. Effect of ectomycorrhizal colonization on the uptake of Ca, Mg and Al by Pinus sylvestris under aluminium toxicity. For. Ecol. Manag. 2005, 215, 352–360. [Google Scholar] [CrossRef]
- Ray, P.; Tiwari, R.; Reddy, U.G.; Adholeya, A. Detecting the heavy metal tolerance level in ectomycorrhizal fungi in vitro. World J. Microbiol. Biotechnol. 2005, 21, 309–315. [Google Scholar] [CrossRef]
- Egerton-Warburton, L. Aluminum-tolerant Pisolithus ectomycorrhizas confer increased growth, mineral nutrition, and metal tolerance to Eucalyptus in acidic mine spoil. Appl. Environ. Soil Sci. 2015, 2015, 803821. [Google Scholar] [CrossRef][Green Version]
- Tahara, K.; Norisada, M.; Tange, T.; Yagi, H.; Kojima, K. Ectomycorrhizal association enhances Al tolerance by inducing citrate secretion in Pinus densiflora. Soil Sci. Plant Nutr. 2005, 51, 397–403. [Google Scholar] [CrossRef]
- Hentschel, E.; Godbold, D.L.; Marschner, P.; Schlegel, H.; Jentschke, G. The effect of Paxillus involutus Fr. on aluminum sensitivity of Norway spruce seedlings. Tree Physiol. 1993, 12, 379–390. [Google Scholar] [CrossRef]
- Khosla, B.; Kaur, H.; Reddy, M.S. Influence of ectomycorrhizal colonization on the growth and mineral nutrition of Populus deltoides under Aluminum toxicity. J. Plant Interact. 2009, 4, 93–99. [Google Scholar] [CrossRef]
- Gu, X.; Liang, G.; Huang, J. Mechanism on increasing plant aluminum resistance by ectomycorrhizae. Chin. Agric. Sci. Bull. 2005, 21, 218–221. [Google Scholar]
- Desai, S.; Naik, D.; Cumming, J.R. The influence of phosphorus availability and Laccaria bicolor symbiosis on phosphate acquisition, antioxidant enzyme activity, and rhizosphere carbon flux in Populus tremuloides. Micorrhiza 2014, 24, 369–382. [Google Scholar] [CrossRef]
- Scholl, L.; Kuyper, T.W.; Smits, M.M.; Landeweert, R.; Hoffland, E.; Breemen, N. Rock-eating mycorrhizas: Their role in plant nutrition and biogeochemical cycles. Plant Soil 2008, 303, 35–47. [Google Scholar] [CrossRef]
- Remiszewski, K.A.; Bryce, J.G.; Fahnestock, M.F.; Pettitt, E.A.; Blichert-Toft, J.; Vadeboncoeur, M.A.; Bailey, S.W. Elemental and isotopic perspectives on the impact of arbuscular mycorrhizal and ectomycorrhizal fungi on mineral weathering across imposed geologic gradients. Chem. Geol. 2016, 445, 164–171. [Google Scholar] [CrossRef]
- Bellion, M.; Courbot, M.; Jacob, C.; Blaudez, D.; Chalot, M. Extracellular and cellular mechanisms sustaining metal tolerance in ectomycorrhizal fungi. FEMS Microbiol. Lett. 2006, 254, 173–181. [Google Scholar] [CrossRef]
- Adeleke, R.; Nwangburuka, C.; Oboirien, B. Origins, roles and fate of organic acids in soils: A review. S. Afr. J. Bot. 2017, 108, 393–406. [Google Scholar] [CrossRef]
- Becquer, A.; Trap, J.; Irshad, U.; Ali, M.A.; Claude, P. From soil to plant, the journey of P through trophic relationships and ectomycorrhizal association. Front. Plant Sci. 2014, 5, 1–7. [Google Scholar] [CrossRef]
- Gu, X.; Liang, G.; Yang, S.; Chen, C.; Huang, J. Influences of Laccaria bicolor on the growth, nutrient uptake and aluminum resistance of Pinus massoniana seedlings. Sci. Silvae Sin. 2005, 41, 199–203. [Google Scholar]
- Gu, X.; Huang, J. Effect of aluminum on growth, oxalate exudation, and uptake of aluminum, phosphorus and potassium by ectomycorrhizal fungi in vitro. Acta Ecol. Sin. 2010, 30, 0357–0363. [Google Scholar]
- Bao, S.D. Soil and Agricultural Chemistry Analysis, 3rd ed.; Chinese Agricultural Press: Beijing, China, 2000; pp. 74–165. [Google Scholar]
- Shinde, S.; Naik, D.; Cumming, J.R. Carbon allocation and partitioning in Populus tremuloides are modulated by ectomycorrhizal fungi under phosphorus limitation. Tree Physiol. 2018, 38, 52–65. [Google Scholar] [CrossRef] [PubMed]
- Yuan, H.; Sun, L.; Tai, P.; Liu, W.; Li, X.; Hao, L. Effects of grafting on root-to-shoot cadmium translocation in plants of eggplant (Solanum melongena) and tomato (Solanum lycopersicum). Sci. Total Environ. 2019, 652, 989–995. [Google Scholar] [CrossRef] [PubMed]
- Thompson, G.W.; Medve, R.J. Effects of aluminum and manganese on the growth of ectomycorrhizal fungi. Appl. Environ. Microbiol. 1984, 48, 556–560. [Google Scholar]
- Bucking, H.; Beckmann, S.; Heyser, W.; Kottke, I. Elemental contents in vacuolar granules of ectomycorrhizal fungi measured by EELS and EDXS. A comparison of different methods and preparation techniques. Micron 1998, 29, 53–61. [Google Scholar] [CrossRef]
- Costa, L.S.; Grazziotti, P.H.; Grazziotti, D.C.F.S.; Silva, A.C.; Rossi, M.J.; Silva, E.B.; Costa, V.H.D.; Gomes, A.L.F. In vitro evaluation of Eucalyptus ectomycorrhizae on substrate with phosphorus doses for fungal pre-selection. Rev. Árvore 2015, 39, 127–136. [Google Scholar] [CrossRef]
- Schier, G.A.; McQuattie, C.J. Effect of aluminum on the growth, anatomy, and nutrient content of ectomycorrhizal and nonmycorrhizal eastern white pine seedlings. Can. J. For. Res. 1995, 25, 1252–1262. [Google Scholar] [CrossRef]
- Huang, J.; Lapeyrie, F. Ability of ectomycorrhizal fungus Laccaria bicolor S238N to increase the growth of Douglas fir seedlings and their phosphorus and potassium uptake. Pedosphere 1996, 6, 217–224. [Google Scholar]
- Choi, D.S.; Jin, H.O.; Chung, D.J.; Sasa, K.; Koike, T. Growth and physiological activity in Larix kaempferi seedlings inoculated with ectomycorrhizae as affected by soil acidification. Trees 2008, 22, 729–735. [Google Scholar] [CrossRef]
- Makita, N.; Hirano, Y.; Yamanaka, T.; Yoshimura, K.; Kosugi, Y. Ectomycorrhizal-fungal colonization induces physio-morphological changes in Quercus serrata leaves and roots. J. Plant Nutr. Soil Sci. 2012, 175, 900–906. [Google Scholar] [CrossRef]
- Fransson, P.M.A.; Anderson, I.C.; Alexander, I.J. Does carbon partitioning in ectomycorrhizal pine seedlings under elevated CO2 vary with fungal species? Plant Soil 2007, 291, 323–333. [Google Scholar] [CrossRef]
- Cairney, J.W.G. Extramatrical mycelia of ectomycorrhizal fungi as moderators of carbon dynamics in forest soil. Soil. Biol. Biochem. 2012, 47, 198–208. [Google Scholar] [CrossRef]
- Felten, J.; Martin, F.; Legué, V. Signaling in ectomycorrhizal symbiosis. In Signaling and Communication in Plants; Baluška, F., Vivanco, J., Eds.; Springer: Berlin, Germany, 2012; pp. 123–142. [Google Scholar]
- Szuba, A. Ectomycorrhiza of Populus. For. Ecol. Manag. 2015, 347, 156–169. [Google Scholar] [CrossRef]
- Szuba, A.; Karlinski, L.; Krzeslowska, M.; Hazubska-Przybyl, T. Inoculation with a Pb-tolerant strain of Paxillus involutus improves growth and Pb tolerance of Populus × canescens under in vitro conditions. Plant Soil 2017, 412, 253–266. [Google Scholar] [CrossRef]
- Verlinden, M.S.; Verbruggen, E.; Janssens, I.A.; Wallander, H.; Vicca, S. Favorable effect of mycorrhizae on biomass production efficiency exceeds their carbon cost in a fertilization experiment. Ecology 2018, 99, 2525–2534. [Google Scholar] [CrossRef] [PubMed]
- Cairney, J.W.G. Ectomycorrhizal fungi: The symbiotic route to the root for phosphorus in forest soils. Plant Soil 2011, 344, 51–71. [Google Scholar] [CrossRef]
- Aggangan, N.S.; Moon, H.K.; Han, S.H. Growth response of Acacia mangium Willd. seedlings to arbuscular mycorrhizal fungi and four isolates of the ectomycorrhizal fungus Pisolithus tinctorius (Pers.) Coker and Couch. New For. 2010, 39, 215–230. [Google Scholar] [CrossRef]
- Hobbie, E.A.; Hoff, C.J.; Bryce, J.G.; Colpaert, J.V.; Hallett, R.A. Nutrient supply rate and mycorrhizal colonization control patterns of element distribution in ectomycorrhizal pine. Commun. Soil Sci. Plant Anal. 2009, 40, 3503–3523. [Google Scholar] [CrossRef]
- Smits, M.M.; Bonneville, S.; Benning, L.G.; Banwart, A.; Leake, J.R. Plant-driven weathering of apatite-the role of an ectomycorrhizal fungus. Geobiology 2012, 10, 445–456. [Google Scholar] [CrossRef]
- Gu, X.; Ni, Y.; Jiang, Y.; Jia, H.; He, X. Effect of Laccaria bicolor inoculation on contents of inorganic phosphorus and labile aluminum in the rhizosphere soil of Pinus massoniana saplings. Acta Pedol. Sin. 2018, 55, 1179–1189. [Google Scholar]
- Browning, M.H.R.; Hutchinson, T.C. The effects of aluminum and calcium on the growth and nutrition of selected ectomycorrhizal fungi of jack pine. Can. J. Bot. 1991, 69, 1691–1699. [Google Scholar] [CrossRef]
- Wu, X.; Zhou, Q.; Gu, X.; Wan, Y.; Mao, W.; Song, Y.; Zeng, Q. Resistance and nutritional response to aluminum toxicity of three ectomycorrhizal fungi from forest soils in Southwest China. J. Southwest Univ. Nat. Sci. Ed. 2015, 37, 13–19. [Google Scholar]
- Heim, A.; Brunner, I.; Frossard, E.; Luster, J. Aluminum effects on Picea abies at low solution concentrations. Soil Sci. Soc. Am. J. 2003, 67, 895–898. [Google Scholar] [CrossRef]
- Kayama, M.; Qu, L.; Koike, T. Elements and ectomycorrhizal symbiosis affecting the growth of Japanese larch seedlings regenerated on slopes of an active volcano in northern Japan. Trees Struct. Funct. 2015, 29, 1567–1579. [Google Scholar] [CrossRef]
- Fernández-Fuego, D.; Keunen, E.; Cuypers, A.; Bertrand, A.; González, A. Mycorrhization protects Betula pubescens Ehr. from metal-induced oxidative stress increasing its tolerance to grow in an industrial polluted soil. J. Hazard. Mater. 2017, 336, 119–127. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Shi, L.; Zhong, K.; Shen, Z.; Chen, Y. Ectomycorrhizal fungi may not act as a barrier inhibiting host plant absorption of heavy metals. Chemosphere 2019, 215, 115–123. [Google Scholar] [CrossRef] [PubMed]
- Aguirre, C.F.V.; Páez, F.A.R.; Vargas, S.E. Effect of arbuscular mycorrhizae and mercury on Lactuca sativa (Asteraceae) seedling morpho—Histology. Environ. Exp. Bot. 2018, 156, 197–202. [Google Scholar] [CrossRef]
- Phanthavongsa, P.; Chalot, M.; Papin, A.; Lacercat-Didier, L.; Roy, S.; Blaudez, D.; Bert, V. Effect of mycorrhizal inoculation on metal accumulation by poplar leaves at phytomanaged sites. Environ. Exp. Bot. 2017, 143, 72–81. [Google Scholar] [CrossRef]
- Fernández-Fuego, D.; Bertrand, A.; González, A. Metal accumulation and detoxification mechanisms in mycorrhizal Betula pubescens. Environ. Pollut. 2017, 231, 1153–1162. [Google Scholar] [CrossRef]
- Moyer-Henry, K.; Macfall, J.; Johannes, E.; Allen, N.; Goldfarb, B.; Rufty, T. Accumulation and localization of aluminum in root tips of loblolly pine seedlings and the associated ectomycorrhizal Pisolithus tinctorius. Plant Cell Environ. 2005, 28, 111–120. [Google Scholar] [CrossRef]
- Vaario, L.; Pennanen, T.; Lu, J.; Palmen, J.; Stenman, J.; Leveinen, J.; Kilpelainen, P.; Kitunen, V. Tricholoma matsutake can absorb and accumulate trace elements directly from rock fragments in the shiro. Micorrhiza 2015, 25, 325–334. [Google Scholar] [CrossRef]
- Wen, Z.; Shi, L.; Tang, Y.; Shen, Z.; Xia, Y.; Chen, Y. Effects of Pisolithus tinctorius and Cenococcum geophilum inoculation on pine in copper-contaminated soil to enhance phytoremediation. Int. J. Phytoremediation 2017, 19, 387–394. [Google Scholar] [CrossRef] [PubMed]
- Gu, X.; Ni, Y.; Jiang, Y.; Jia, H. Effects of Laccaria bicolor on growth, uptake and distribution of nutrients and aluminum of Pinus massoniana seedlings under acid aluminum exposure. Sci. Silvae Sin. 2018, 54, 170–178. [Google Scholar]
- Liu, R.; Liu, H. Effect of acidity and aluminum on the growth of Pinus massoniana seedlings. Acta Bot. Sin. 1995, 37, 154–158. [Google Scholar]




© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).