Tropical Dry Forest Diversity, Climatic Response, and Resilience in a Changing Climate



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
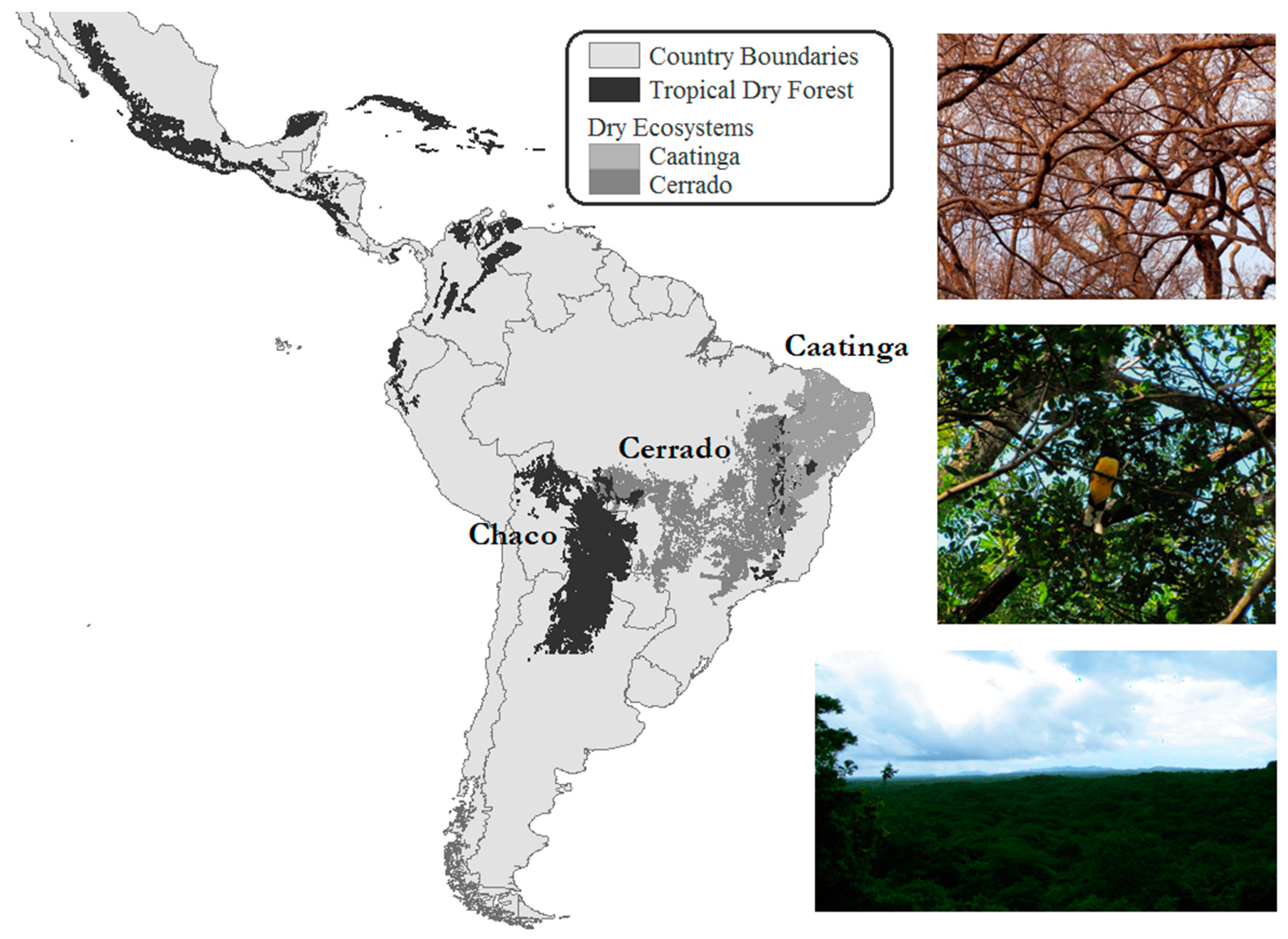
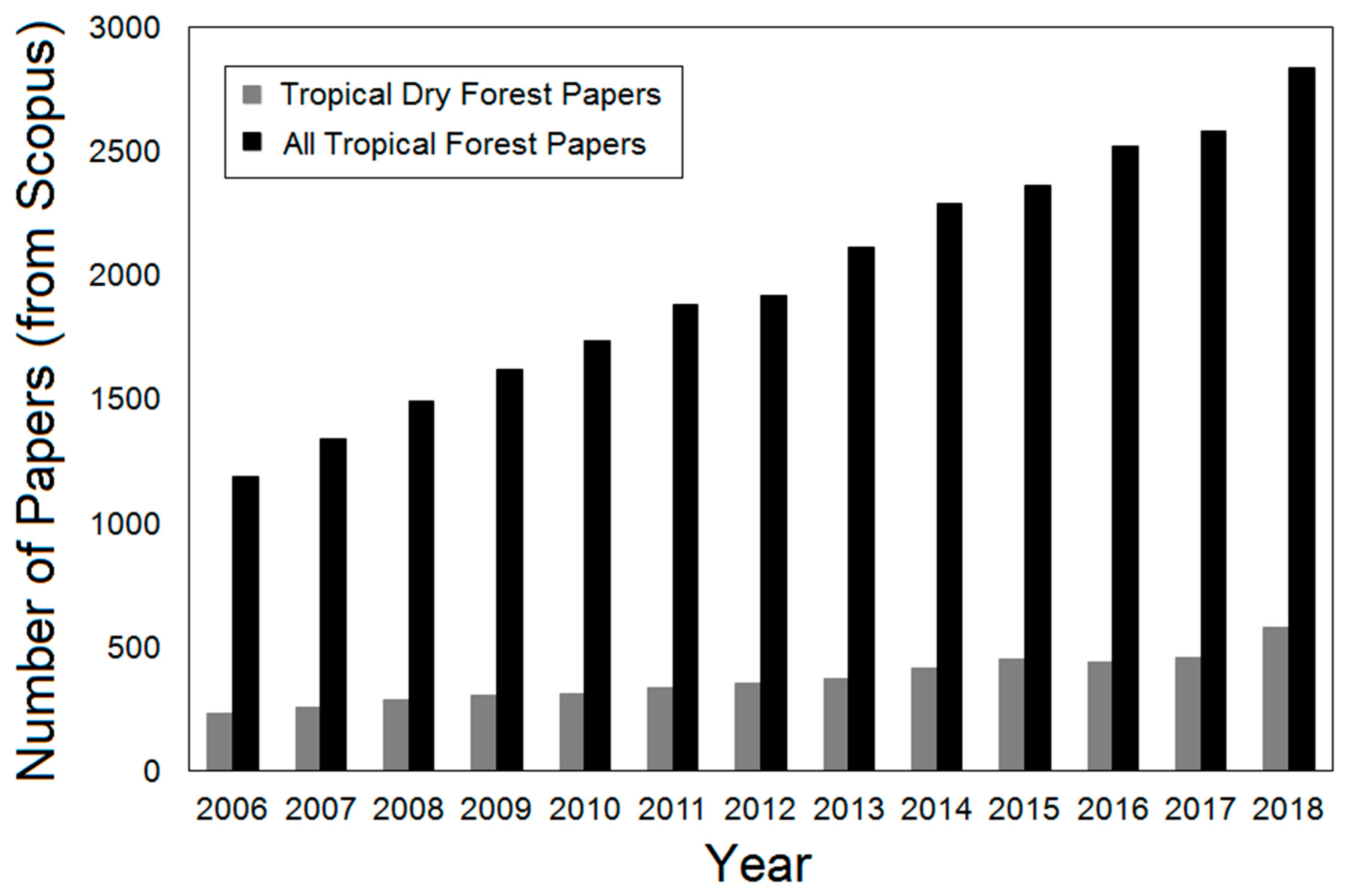
:1. Introduction
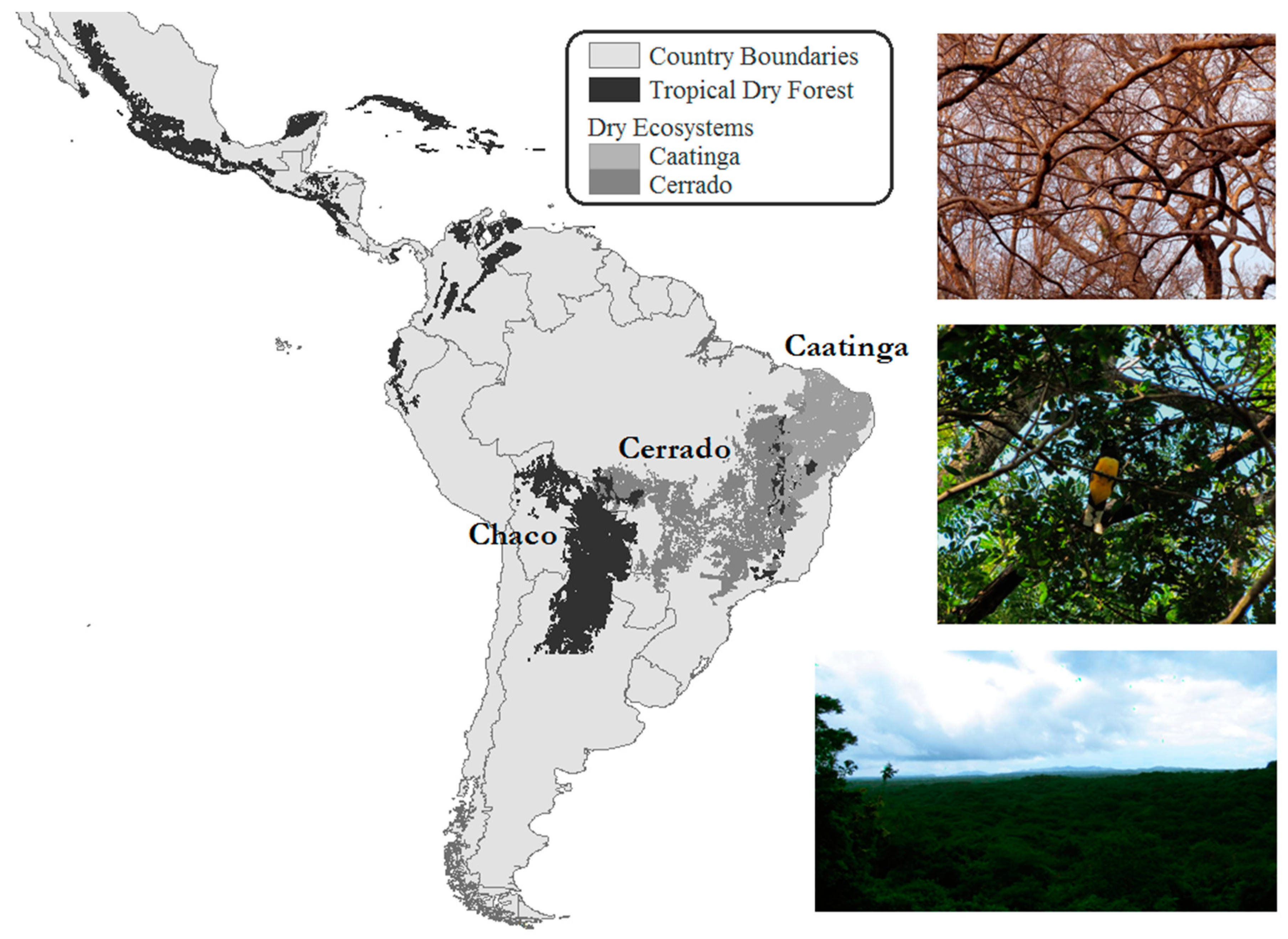
2. Components of Tropical Dry Forests
2.1. Forest Dynamics
2.2. Forest Structure
2.3. Phenological Cycles
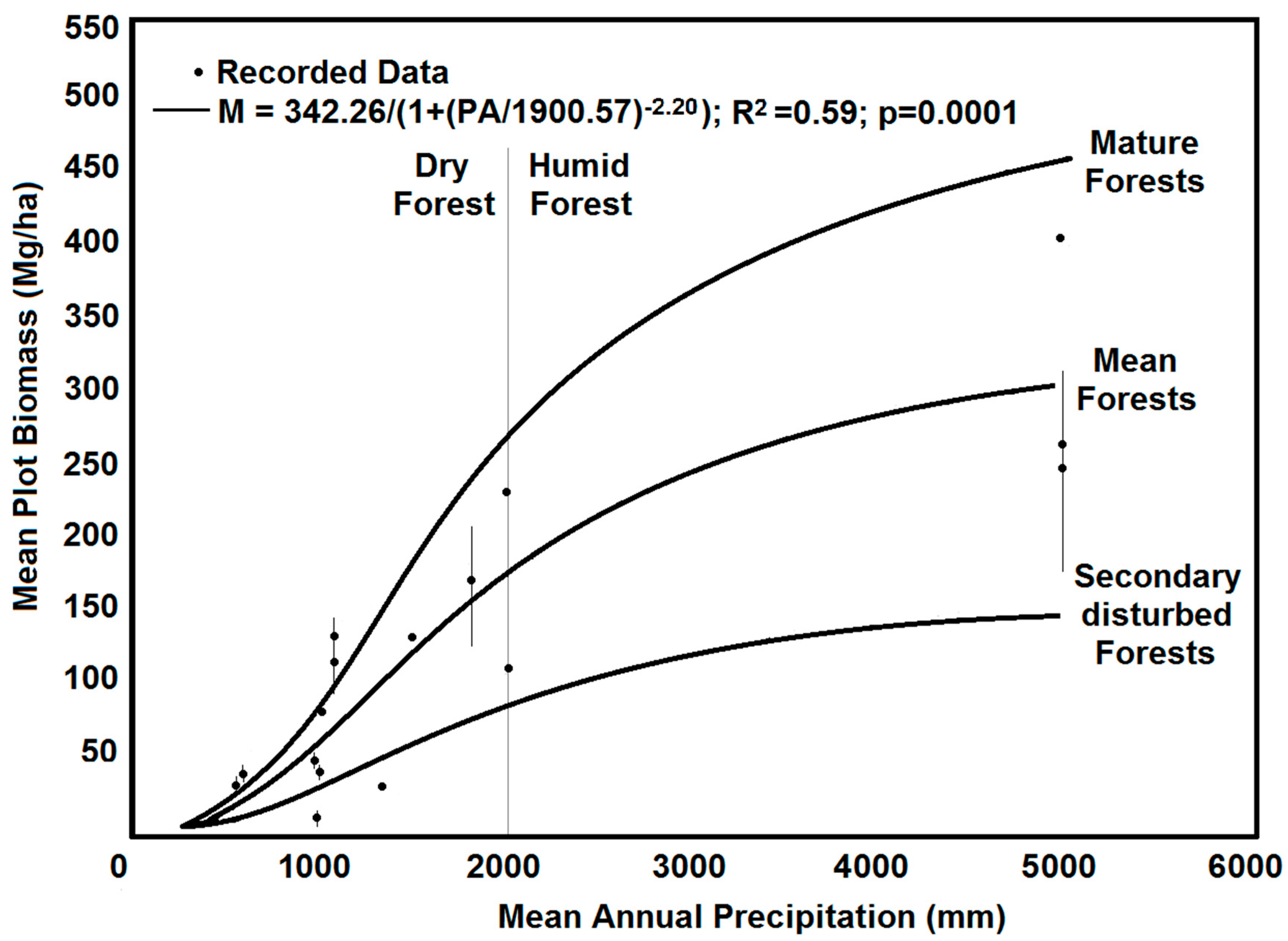
2.4. Above-Ground Biomass (AGB)
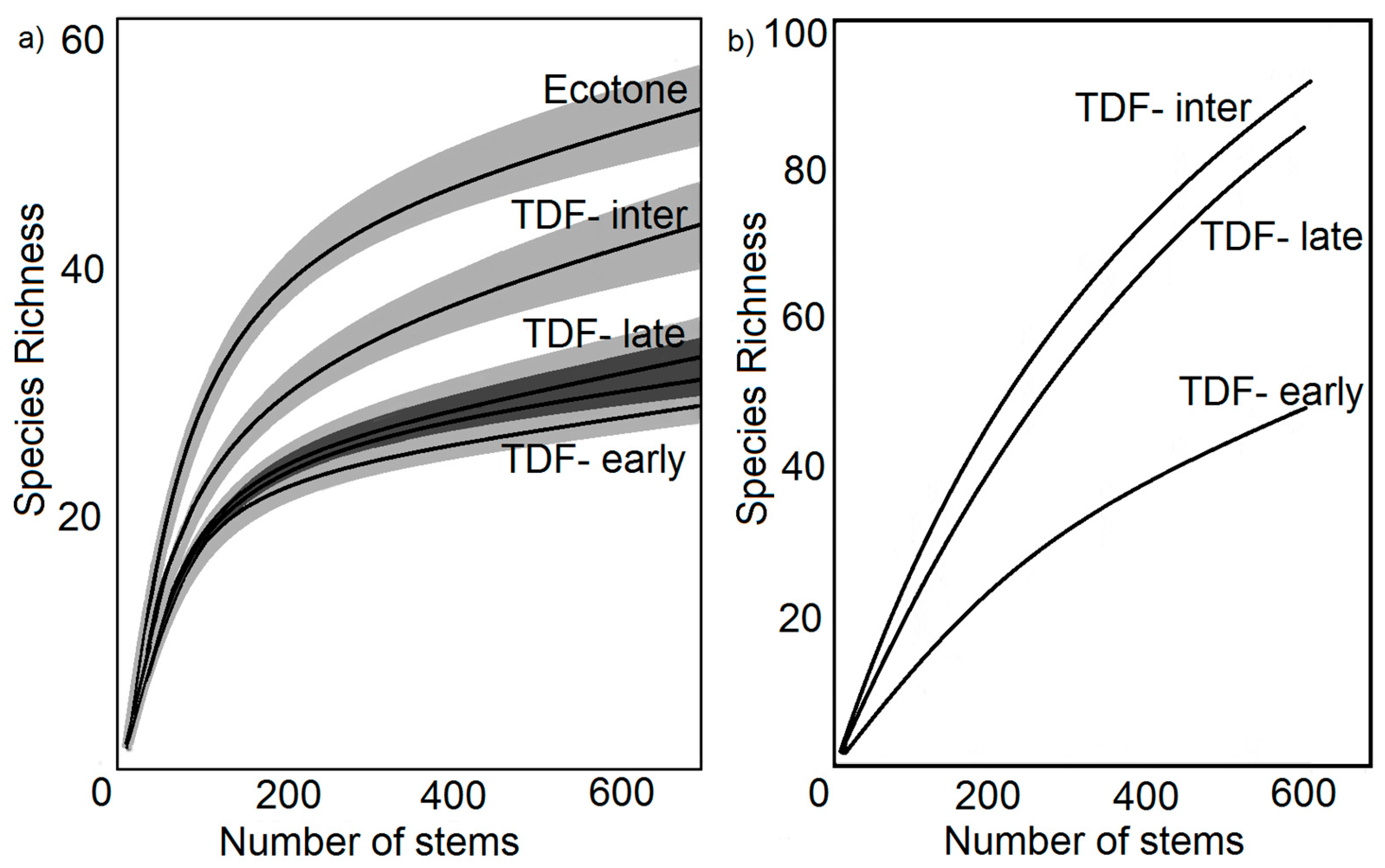
2.5. Biodiversity
3. Tropical Dry Forest Response to Variations in Climate
3.1. Temperature
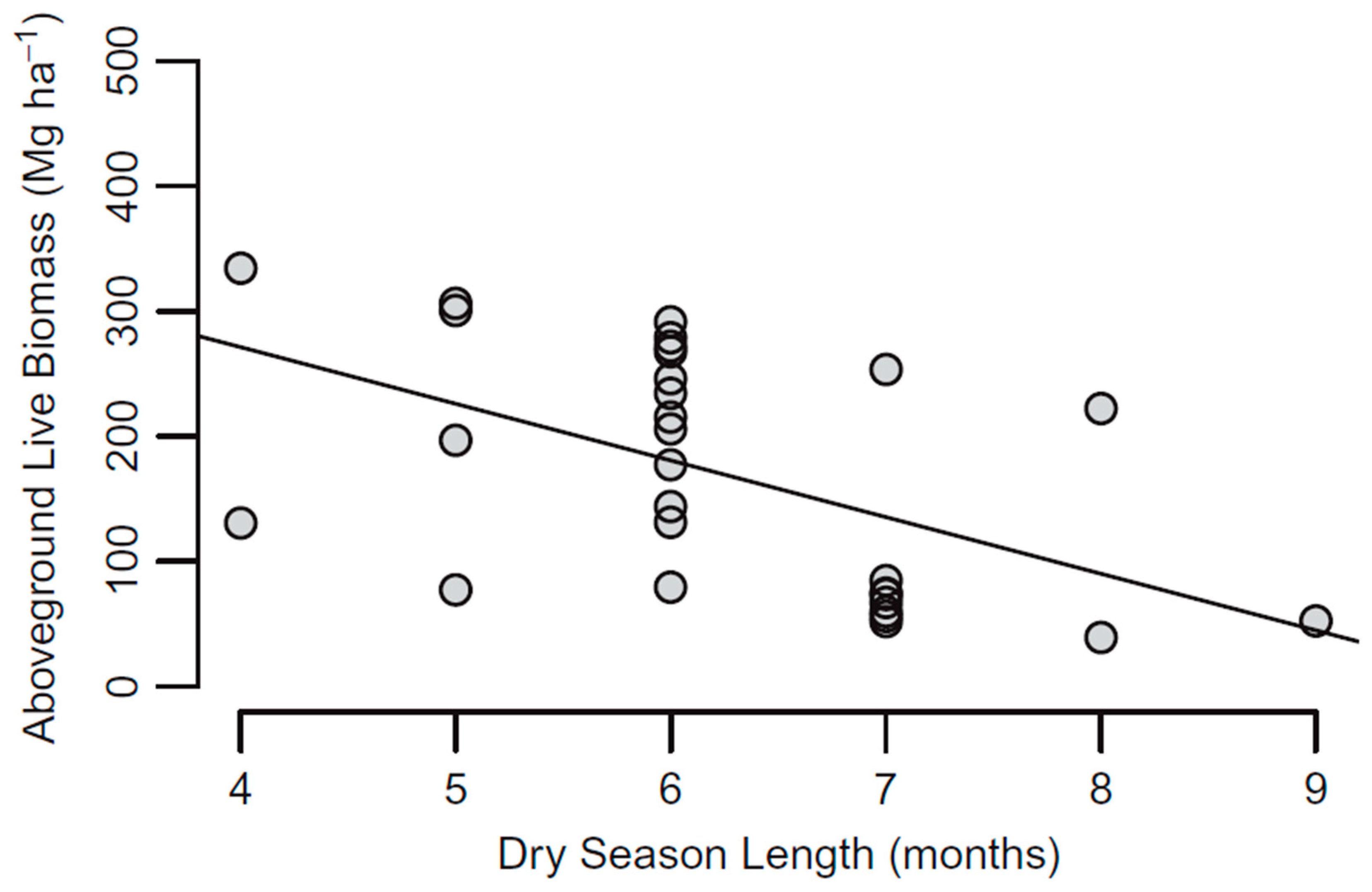
3.2. Response to Variation in Precipitation
3.3. Natural Disturbances
3.3.1. Tropical Storms
3.3.2. Fire
3.3.3. Drought
4. Resilience under Climatic Change
5. Conclusions
Funding
Acknowledgments
Conflicts of Interest
References
- Magrin, G.; Marengo, J.; Boulanger, J.P.; Buckeridge, M.S.; Castellanos, E.; Poveda, G.; Scarano, F.R.; Vicuña, S. Central and South America. In Climate Change 2014: Impacts, Adaptation and Vulnerability. Contribution of Working Group II to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Barros, V.R., Field, C.B., Dokken, D.J., Mastrandrea, M.D., Mach, K.J., Bilir, T.E., Chatterjee, M., Ebi, K.L., Estrada, Y.O., Genova, R.C., Eds.; Cambridge University Press: Cambridge, UK, 2014. [Google Scholar]
- Yamamoto, L.; Serraglio, D.A.; Cavedon-Capdeville, F.S. Human mobility in the context of climate change and disasters: A South American approach. Int. J. Clim. Chang. Strateg. Manag. 2018, 10, 65–85. [Google Scholar] [CrossRef]
- Thiede, B.; Gray, C.; Mueller, V. Climate variability and inter-provincial migration in South America, 1970-2011. Glob. Environ. Chang. 2016, 41, 228–240. [Google Scholar] [CrossRef]
- Pivetta, M. 50 Anos de Calamidades na América do Sul. Revista Pesquisa FAPESP 2016, 241, 62–65. [Google Scholar]
- Bender, M.A.; Knutson, T.R.; Tuleya, R.E.; Sirutis, J.J.; Vecchi, G.A.; Garner, S.T.; Held, I.M. Modeled Impact of Anthropogenic Warming on the Frequency of Intense Atlantic Hurricanes. Science 2010, 327, 454–458. [Google Scholar] [CrossRef] [Green Version]
- Cai, W.; Santoso, A.; Wang, G.; Yeh, S.-W.; An, S.-I.; Cobb, K.M.; Collins, M.; Guilyardi, E.; Jin, F.-F.; Kug, J.-S. ENSO and greenhouse warming. Nat. Clim. Chang. 2015, 5, 849. [Google Scholar] [CrossRef]
- Wright, S.J. Tropical forests in a changing environment. Trends Ecol. Evol. 2005, 20, 553–560. [Google Scholar] [CrossRef]
- Miles, L.; Newton, A.C.; DeFries, R.S.; Ravilious, C.; May, I.; Blyth, S.; Kapos, V.; Gordon, J.E. A global overview of the conservation status of tropical dry forests. J. Biogeogr. 2006, 33, 491–505. [Google Scholar] [CrossRef]
- Quesada, M.; Sanchez-Azofeifa, G.A.; Alvarez-Añorve, M.; Stoner, K.E.; Avila-Cabadilla, L.; Calvo-Alvarado, J.; Castillo, A.; Espírito-Santo, M.M.; Fagundes, M.; Fernandes, G.W.; et al. Succession and management of tropical dry forests in the Americas: Review and new perspectives. For. Ecol. Manag. 2009, 258, 1014–1024. [Google Scholar] [CrossRef]
- Buzzard, V.; Hulshof, C.M.; Birt, T.; Violle, C.; Enquist, B.J. Re-growing a tropical dry forest: Functional plant trait composition and community assumble during succession. Funct. Ecol. 2016, 30, 1006–1013. [Google Scholar] [CrossRef]
- Banda, R.K.; Delgado-Salinas, A.; Dexter, K.G.; Linares-Palomino, R.; Oliveira-Filho, A.; Prado, D.; Pullan, M.; Quintana, C.; Riina, R.; Rodriguez, M.G.M.; et al. Plant diversity patterns in neotropical dry forests and their conservation implications. Science 2016, 353, 1383–1387. [Google Scholar] [CrossRef] [Green Version]
- Golicher, D.J.; Cayuela, L.; Newton, A.C. Effects of climate change on the potential species richness of mesoamerican forests. Biotropica 2012, 44, 284–293. [Google Scholar] [CrossRef]
- Powers, J.S.; Becknell, J.M.; Irving, J.; Pèrez-Aviles, D. Diversity and structure of regenerating tropical dry forests in Costa Rica: Geographic patterns and environmental drivers. For. Ecol. Manag. 2009, 258, 959–970. [Google Scholar] [CrossRef]
- Portillo-Quintero, C.A.; Sánchez-Azofeifa, G.A. Extent and conservation of tropical dry forests in the Americas. Biol. Conserv. 2010, 143, 144–155. [Google Scholar] [CrossRef]
- Hesketh, M.; Sánchez-Azofeifa, A. A review of remote sensing of tropical dry forests. In Tropical Dry Forests in the Americas: Ecology, Conservation, and Management; Sánchez-Azofeifa, A., Powers, J.S., Fernandes, G.W., Quesada, M., Eds.; CRC Press: Boca Raton, FL, USA, 2013; pp. 80–98. [Google Scholar] [CrossRef]
- Sánchez-Azofeifa, G.A.; Quesada, M.; Rodríguez, J.P.; Nassar, J.M.; Stoner, K.E.; Castillo, A.; Garvin, T.; Zent, E.L.; Calvo-Alvarado, J.C.; Kalacska, M.E.R.; et al. Research Priorities for Neotropical Dry Forests. Biotropica 2005, 37, 477–485. [Google Scholar] [CrossRef]
- Kennard, D.K. Secondary forest succession in a tropical dry forest: Patterns of development across a 50-year chronosequence in lowland Bolivia. J. Trop. Ecol. 2002, 18, 53–66. [Google Scholar] [CrossRef]
- Hansen, M.C.; Stehman, S.V.; Potapov, P.V. Quantification of global gross forest cover loss. Proc. Natl. Acad. Sci. USA 2010, 107, 8650–8655. [Google Scholar] [CrossRef] [Green Version]
- Coelho, P.A.; Santos, P.F.; de Paiva Paula, E.; Apgaua, D.M.G.; Madeira, B.G.; Menino, G.C.d.O.; Nunes, Y.R.F.; Santos, R.M.; Tng, D.Y.P. Tree succession across a seasonally dry tropical forest and forest-savanna ecotone in northern Minas Gerais, Brazil. J. Plant Ecol. 2016, 10, 858–868. [Google Scholar] [CrossRef]
- Fajardo, L.; Gonzalez, V.; Nassar, J.M.; Lacabana, P.; Portillo, Q.C.A.; Carrasquel, F.; Rodriguez, J.P. Tropical Dry Forests of Venezuela: Characterization and Current Conservation Status1. Biotropica 2005, 37, 531–546. [Google Scholar] [CrossRef]
- Portillo-Quintero, C.; Sanchez-Azofeifa, A.; Calvo-Alvarado, J.; Quesada, M.; Do Espirito Santo, M.M. The role of tropical dry forests for biodiversity, carbon and water conservation in the neotropics: Lessons learned and opportunities for its sustainable management. Reg. Environ. Chang. 2015, 15, 1039–1049. [Google Scholar] [CrossRef]
- Sanchez-Azofeifa, G.A.; Kalacska, M.; Quesada, M.; Calvo-Alvarado, J.C.; Nassar, J.M.; Rodriguez, J.P. Need for Integrated Research for a Sustainable Future in Tropical Dry Forests. Conserv. Biol. 2005, 19, 285–286. [Google Scholar] [CrossRef]
- Calvo-Rodriguez, S.; Sanchez-Azofeifa, A.G.; Duran, S.M.; Espirito-Santo, M.M. Assessing ecosystem services in Neotropical dry forests: A systematic review. Environ. Conserv. 2017, 44, 34–43. [Google Scholar] [CrossRef]
- Garaglio, M.A.; Sampaio, E.V.d.S.B.; Cestaro, L.A.; Kageyama, P.Y. Uso Sustentável e Conservação dos Recursos Florestais da Caatinga; Serviço Florestal Brasileiro: Brasilia, Brazil, 2010; p. 368.
- Robalino, J.; Pfaff, A.; Villalobos, L. Heterogeneous Local Spillovers from Protected Areas in Costa Rica. J. Assoc. Environ. Resour. Econ. 2017, 4, 795–820. [Google Scholar] [CrossRef]
- Stan, K.; Sanchez-Azofeifa, A. Deforestation and secondary growth in Costa Rica along the path of development. Reg. Environ. Chang. 2019, 19, 587–597. [Google Scholar] [CrossRef]
- Porter-Bolland, L.; Ellis, E.A.; Guariguata, M.R.; Ruiz-Mallén, I.; Negrete-Yankelevich, S.; Reyes-García, V. Community managed forests and forest protected areas: An assessment of their conservation effectiveness across the tropics. For. Ecol. Manag. 2012, 268, 6–17. [Google Scholar] [CrossRef]
- Becknell, J.M.; Kissing Kucek, L.; Powers, J.S. Aboveground biomass in mature and secondary seasonally dry tropical forests: A literature review and global synthesis. For. Ecol. Manag. 2012, 276, 88–95. [Google Scholar] [CrossRef]
- Allen, K.; Dupuy, J.M.; Gei, M.; Hulshof, C.; Medvigy, D.; Pizano, C.; Salgado Negret, B.; Smith, C.; Trierweiler, A.; Van Bloem, S.; et al. Will seasonally dry tropical forests be sensitive or resistant to future changes in rainfall regimes? Environ. Res. Lett. 2017, 12, 023001. [Google Scholar] [CrossRef] [Green Version]
- Grebner, D.L.; Bettinger, P.; Siry, J.P. Chapter 10—Forest Dynamics. In Introduction to Forestry and Natural Resources; Grebner, D.L., Bettinger, P., Siry, J.P., Eds.; Academic Press: Cambridge, MA, USA, 2013; pp. 243–254. [Google Scholar] [CrossRef]
- Ma, L.; Lian, J.; Lin, G.; Cao, H.; Huang, Z.; Guan, D. Forest dynamics and its driving forces of sub-tropical forest in South China. Sci. Rep. 2016, 6, 22561. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Cao, S.; Campos-Vargas, C.; Sanchez-Azofeifa, A. Identifying tropical dry forests extent and succession via the use of machine learning techniques. Int. J. Appl. Earth Observ. Geoinf. 2017, 63, 196–205. [Google Scholar] [CrossRef]
- Cao, S.; Sanchez-Azofeifa, A. Modeling seasonal surface temperature variations in secondary tropical dry forests. Int. J. Appl. Earth Observ. Geoinf. 2017, 62, 122–134. [Google Scholar] [CrossRef]
- Johnson, E.A.; Miyanishi, K. Testing the assumptions of chronosequences in succession. Ecol. Lett. 2008, 11, 419–431. [Google Scholar] [CrossRef]
- Madeira, B.G.; Espírito-Santo, M.M.; Neto, S.D.Â.; Nunes, Y.R.F.; Arturo Sánchez Azofeifa, G.; Wilson Fernandes, G.; Quesada, M. Changes in tree and liana communities along a successional gradient in a tropical dry forest in south-eastern Brazil. Plant Ecol. 2009, 201, 291–304. [Google Scholar] [CrossRef]
- Kalacska, M.E.R.; Sánchez-Azofeifa, G.A.; Calvo-Alvarado, J.C.; Rivard, B.; Quesada, M. Effects of Season and Successional Stage on Leaf Area Index and Spectral Vegetation Indices in Three Mesoamerican Tropical Dry Forests. Biotropica 2005, 37, 486–496. [Google Scholar] [CrossRef]
- Chazdon, R.L.; Letcher, S.G.; Van Breugel, M.; Martínez-Ramos, M.; Bongers, F.; Finegan, B. Rates of change in tree communities of secondary Neotropical forests following major disturbances. Philos. Trans. R. Soc. B Biol. Sci. 2007, 362, 273–289. [Google Scholar] [CrossRef]
- Martínez-Ramos, M.; Balvanera, P.; Arreola Villa, F.; Mora, F.; Maass, J.M.; Maza-Villalobos Méndez, S. Effects of long-term inter-annual rainfall variation on the dynamics of regenerative communities during the old-field succession of a neotropical dry forest. For. Ecol. Manag. 2018, 426, 91–100. [Google Scholar] [CrossRef]
- Hernández-Ramírez, A.M.; García-Méndez, S. Diversidad, estructura y regeneración de la selva tropical estacionalmente seca de la Península de Yucatán, México. Revista de Biología Tropical 2015, 63, 603–616. [Google Scholar] [CrossRef]
- Derroire, G.; Balvanera, P.; Castellanos-Castro, C.; Decocq, G.; Kennard, D.K.; Lebrija-Trejos, E.; Leiva, J.A.; Odén, P.-C.; Powers, J.S.; Rico-Gray, V.; et al. Resilience of tropical dry forests—A meta-analysis of changes in species diversity and composition during secondary succession. Oikos 2016, 125, 1386–1397. [Google Scholar] [CrossRef]
- Singh, K.; Kushwaha, C. Emerging paradigms of tree phenology in dry tropics. Curr. Sci. 2005, 89, 964–975. [Google Scholar]
- Wright, S.J.; Calderón, O. Seasonal, El Niño and longer term changes in flower and seed production in a moist tropical forest. Ecol. Lett. 2006, 9, 35–44. [Google Scholar] [CrossRef]
- Lebrija-Trejos, E.; Bongers, F.; Pérez-García, E.A.; Meave, J.A. Successional change and resilience of a very dry tropical deciduous forest following shifting agriculture. Biotropica 2008, 40, 422–431. [Google Scholar] [CrossRef]
- Brienen, R.J.W.; Lebrija-Trejos, E.; Zuidema, P.A.; Martínez-Ramos, M. Climate-growth analysis for a Mexican dry forest tree shows strong impact of sea surface temperatures and predicts future growth declines. Glob. Chang. Biol. 2010, 16, 2001–2012. [Google Scholar] [CrossRef]
- Kalacska, M.; Sanchez-Azofeifa, G.A.; Calvo-Alvarado, J.C.; Quesada, M.; Rivard, B.; Janzen, D.H. Species composition, similarity and diversity in three successional stages of a seasonally dry tropical forest. For. Ecol. Manag. 2004, 200, 227–247. [Google Scholar] [CrossRef]
- Castillo-Núñez, M.; Sánchez-Azofeifa, G.A.; Croitoru, A.; Rivard, B.; Calvo-Alvarado, J.; Dubayah, R.O. Delineation of secondary succession mechanisms for tropical dry forests using LiDAR. Remote Sens. Environ. 2011, 115, 2217–2231. [Google Scholar] [CrossRef]
- Hilje, B.; Calvo-Alvarado, J.; Jiménez-Rodríguez, C.; Sánchez-Azofeifa, A. Tree Species Composition, Breeding Systems, and Pollination and Dispersal Syndromes in Three Forest Successional Stages in a Tropical Dry Forest in Mesoamerica. Trop. Conserv. Sci. 2015, 8, 76–94. [Google Scholar] [CrossRef]
- Jaramillo, V.J.; Kauffman, J.B.; Renteria-Rodriguez, L.; Cummings, D.L.; Ellingson, L.J. Biomass, Carbon, and Nitrogen Pools in Mexican Tropical Dry Forest Landscapes. Ecosystems 2003, 6, 609–629. [Google Scholar] [CrossRef]
- Vilanova, E.; Ramirez-Angulo, H.; Torres-Lezama, A.; Aymard, G.; Gamez, L.; Duran, C.; Hernandez, L.; Herrera, R.; van der Heijden, G.; Phillips, O.L.; et al. Environmental drivers of forest structure and stem turnover across Venezuelan tropical forests. PLoS ONE 2018, 13, e0198489. [Google Scholar] [CrossRef] [PubMed]
- Peña-Claros, M.; Poorter, L.; Alarcón, A.; Blate, G.; Choque, U.; Fredericksen, T.S.; Justiniano, M.J.; Leaño, C.; Licona, J.C.; Pariona, W.; et al. Soil Effects on Forest Structure and Diversity in a Moist and a Dry Tropical Forest. Biotropica 2012, 44, 276–283. [Google Scholar] [CrossRef]
- Durán, S.M.; Sánchez-Azofeifa, G.A.; Rios, R.S.; Gianoli, E. The relative importance of climate, stand variables and liana abundance for carbon storage in tropical forests. Glob. Ecol. Biogeogr. 2015, 24, 939–949. [Google Scholar] [CrossRef]
- Schnitzer, S.A.; Bongers, F. Increasing liana abundance and biomass in tropical forests: Emerging patterns and putative mechanisms. Ecol. Lett. 2011, 14, 397–406. [Google Scholar] [CrossRef] [PubMed]
- Kalacska, M.; Sanchez-Azofeifa, G.A.; Rivard, B.; Calvo-Alvarado, J.C.; Quesada, M. Baseline assessment for environmental services payments from satellite imagery: A case study from Costa Rica and Mexico. J. Environ. Manag. 2008, 88, 348–359. [Google Scholar] [CrossRef]
- Dewalt, S.J.; Schnitzer, S.A.; Chave, J.; Bongers, F.; Burnham, R.J.; Cai, Z.; Chuyong, G.; Clark, D.B.; Ewango, C.E.N.; Gerwing, J.J.; et al. Annual Rainfall and Seasonality Predict Pan-tropical Patterns of Liana Density and Basal Area. Biotropica 2010, 42, 309–317. [Google Scholar] [CrossRef]
- Schnitzer, S.A. A mechanistic explanation for global patterns of liana abundance and distribution. Am. Nat. 2005, 166, 262–276. [Google Scholar] [CrossRef]
- Van Der Heijden, G.M.F.; Phillips, O.L. What controls liana success in Neotropical forests? Glob. Ecol. Biogeogr. 2008, 17, 372–383. [Google Scholar] [CrossRef]
- Reich, P.B.; Borchert, R. Water stress and tree phenology in a tropical dry forest in the lowlands of Costa Rica. J. Ecol. 1994, 72, 61–74. [Google Scholar] [CrossRef]
- Rankine, C.; Sánchez-Azofeifa, G.A.; Guzmán, J.A.; Espirito-Santo, M.M.; Sharp, I. Comparing MODIS and near-surface vegetation indexes for monitoring tropical dry forest phenology along a successional gradient using optical phenology towers. Environ. Res. Lett. 2017, 12. [Google Scholar] [CrossRef]
- Elliott, S.; Baker, P.J.; Borchert, R. Leaf flushing during the dry season: The paradox of Asian monsoon forests. Glob. Ecol. Biogeogr. 2006, 15, 248–257. [Google Scholar] [CrossRef]
- Hulshof, C.M.; Martínez-Yrízar, A.; Burquez, A.; Boyle, B.; Enquist, B.J. Plant functional trait variation in tropical dry forests: A review and synthesis. In Tropical Dry Forests in the Americas: Ecology, Conservation, and Management; Sánchez-Azofeifa, A., Powers, J.S., Fernandes, G.W., Quesada, M., Eds.; CRC Press: Boca Raton, FL, USA, 2013; pp. 129–140. [Google Scholar] [CrossRef]
- Lopezaraiza-Mikel, M.; Quesada, M.; Álvarez-Añorve, M.; Ávila-Cabadilla, L.; Martén-Rodríguez, S.; Calvo-Alvarado, J.; do Espírito-Santo, M.M.; Fernandes, G.W.; Sánchez-Azofeifa, A.; de Jesús Aguilar-Aguilar, M. Phenological patterns of tropical dry forests along latitudinal and successional gradients in the neotropics. In Tropical Dry Forests in the Americas: Ecology, Conservation, and Management; Sánchez-Azofeifa, A., Powers, J.S., Fernandes, G.W., Quesada, M., Eds.; CRC Press: Boca Raton, FL, USA, 2013; pp. 113–140. [Google Scholar] [CrossRef]
- Castro, S.M.; Sanchez-Azofeifa, G.A.; Sato, H. Effect of drought on productivity in a Costa Rican tropical dry forest. Environ. Res. Lett. 2018, 13. [Google Scholar] [CrossRef]
- Pezzini, F.F.; Ranieri, B.D.; Brandão, D.O.; Fernandes, G.W.; Quesada, M.; Espírito-Santo, M.M.; Jacobi, C.M. Changes in tree phenology along natural regeneration in a seasonally dry tropical forest. Plant Biosyst. Int. J. Deal. Asp. Plant Biol. 2014, 148, 965–974. [Google Scholar] [CrossRef]
- Ravindrath, N.H.; Ostwald, M. Carbon Inventory Methods: Handbook for Greenhouse Gas Inventory, Carbon Mitigation and Roundwood Production Projects; Beninston, M., Ed.; Springer: New York, NY, USA, 2008; p. 315. [Google Scholar] [CrossRef]
- Poorter, L.; Bongers, F.; Aide, T.M.; Almeyda Zambrano, A.M.; Balvanera, P.; Becknell, J.M.; Boukili, V.; Brancalion, P.H.; Broadbent, E.N.; Chazdon, R.L.; et al. Biomass resilience of Neotropical secondary forests. Nature 2016, 530, 211–214. [Google Scholar] [CrossRef]
- Rozendaal, D.M.A.; Chazdon, R.L.; Arreola-Villa, F.; Balvanera, P.; Bentos, T.V.; Dupuy, J.M.; Hernández-Stefanoni, J.L.; Jakovac, C.C.; Lebrija-Trejos, E.E.; Lohbeck, M.; et al. Demographic Drivers of Aboveground Biomass Dynamics During Secondary Succession in Neotropical Dry and Wet Forests. Ecosystems 2016, 20, 340–353. [Google Scholar] [CrossRef]
- Navar Chaidez, J.D.J. The Spatial Distribution of Aboveground Biomass in Tropical Forests of Mexico. Trop. Subtrop. Agroecosyst. 2010, 13, 10. [Google Scholar]
- De Jong, B.H.; Gutiérrez, L.I.; de la Rosa, J.A.A. Advances of Mexico in Preparing for REDD: UNFCCC Workshop on Methodological Issues Relating to Reducing Emissions from Deforestation and Forest Degradation in Developing Countries; UNFCCC: Tokyo, Japan, 2008. [Google Scholar]
- Houghton, R.A. The annual net flux of carbon to the atmosphere from changes in land use 1850–1990. Tellus B 1999, 51, 298–313. [Google Scholar] [CrossRef]
- Defries, R.S.; Houghton, R.A.; Hansen, M.C.; Field, C.B.; Skole, D.; Townshend, J. Carbon emissions from tropical deforestation and regrowth based on satellite observations for the 1980s and 1990s. Proc. Natl. Acad. Sci. USA 2002, 99, 14256–14261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, S. Estimating Biomass and Biomass Change of Tropical Forests: A Primer; Food & Agriculture Org.: Rome, Italy, 1997; Volume 134. [Google Scholar]
- Achard, F.; Eva, H.D.; Mayaux, P.; Stibig, H.-J.; Belward, A. Improved estimates of net carbon emissions from land cover change in the tropics for the 1990s. Glob. Biogeochem. Cycles 2004, 18, GB2008. [Google Scholar] [CrossRef]
- IPCC. IPCC Guidelines for National Greenhouse Gas Inventories—A Primer; Institute for Global Environmental Strategires: Hayama, Japan, 2006. [Google Scholar]
- Corona-Núñez, R.O.; Mendoza-Ponce, A.; López-Martínez, R. Model selection changes the spatial heterogeneity and total potential carbon in a tropical dry forest. For. Ecol. Manag. 2017, 405, 69–80. [Google Scholar] [CrossRef]
- Montoro-Girona, M.; Morin, H.; Lussier, J.M.; Walsh, D. Radial growth response of black spruce stands ten years after experimental shelterwoods and seed-tree cuttings in borel forest. Forests 2016, 7, 20. [Google Scholar] [CrossRef]
- Montoro-Girona, M.; Rossi, S.; Lussier, J.M.; Walsh, D.; Morin, H. Understanding tree growth responses after partial cuttings: A new approach. PLoS ONE 2016, 12, e0172653. [Google Scholar] [CrossRef] [PubMed]
- Rito, K.F.; Arroyo-Rodríguez, V.; Queiroz, R.T.; Leal, I.R.; Tabarelli, M.; Norden, N. Precipitation mediates the effect of human disturbance on the Brazilian Caatinga vegetation. J. Ecol. 2017, 105, 828–838. [Google Scholar] [CrossRef] [Green Version]
- Trejo, I.; Dirzo, R. Floristic diversity of Mexican seasonally dry tropical forests. Biodivers. Conserv. 2002, 11, 2063–2084. [Google Scholar] [CrossRef]
- Segura, G.; Balvanera, P.; Durán, E.; Pérez, A. Tree community structure and stem mortality along a water availability gradient in a Mexican tropical dry forest. Plant Ecol. 2002, 169, 259–271. [Google Scholar] [CrossRef]
- Vargas-Rodriguez, Y.L.; Vazquez-Garcıa, J.A.; Williamson, G.B. Environmental correlates of tree and seedling-sapling distributions in a Mexican tropical dry forest. Plant Ecol. 2005, 180, 117–134. [Google Scholar] [CrossRef]
- Saiter, F.Z.; Eisenlohr, P.V.; Barbosa, M.R.V.; Thomas, W.W.; Oliveira-Filho, A.T. From evergreen to deciduous tropical forests: How energy–water balance, temperature, and space influence the tree species composition in a high diversity region. Plant Ecol. Divers. 2015, 9, 45–54. [Google Scholar] [CrossRef]
- Power, M.J.; Whitney, B.S.; Mayle, F.E.; Neves, D.M.; de Boer, E.J.; Maclean, K.S. Fire, climate and vegetation linkages in the Bolivian Chiquitano seasonally dry tropical forest. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2016, 371. [Google Scholar] [CrossRef] [PubMed]
- Easterling, D.R.; Meehl, G.A.; Parmesan, C.; Changnon, S.A.; Karl, T.R.; Mearns, L.O. Climate Extremes: Observations, Modeling, and Impacts. Science 2000, 289, 2068–2074. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walther, G.R. Community and ecosystem responses to recent climate change. Philos. Trans. R. Soc. B Biol. Sci. 2010, 365, 2019–2024. [Google Scholar] [CrossRef]
- Mendivelso, H.A.; Camarero, J.J.; Gutiérrez, E.; Zuidema, P.A. Time-dependent effects of climate and drought on tree growth in a Neotropical dry forest: Short-term tolerance vs. long-term sensitivity. Agric. For. Meteorol. 2014, 188, 13–23. [Google Scholar] [CrossRef] [Green Version]
- Álvarez-Dávila, E.; Cayuela, L.; González-Caro, S.; Aldana, A.M.; Stevenson, P.R.; Phillips, O.; Cogollo, Á.; Peñuela, M.C.; Von Hildebrand, P.; Jiménez, E.; Melo, O.; et al. Forest biomass density across large climate gradients in northern South America is related to water availability but not with temperature. PLoS ONE 2017, 12, e0171072. [Google Scholar] [CrossRef] [PubMed]
- Slot, M.; Winter, K. In situ temperature response of photosynthesis of 42 tree and liana species in the canopy of two Panamanian lowland tropical forests with contrasting rainfall regimes. New Phytol. 2017, 214, 1103–1117. [Google Scholar] [CrossRef] [PubMed]
- Haselhorst, D.S.; Tcheng, D.K.; Moreno, J.E.; Punyasena, S.W. The effects of seasonal and long-term climatic variability on Neotropical flowering phenology: An ecoinformatic analysis of aerial pollen data. Ecol. Inform. 2017, 41, 54–63. [Google Scholar] [CrossRef]
- Pau, S.; Wolkovich, E.M.; Cook, B.I.; Nytch, C.J.; Regetz, J.; Zimmerman, J.K.; Joseph Wright, S. Clouds and temperature drive dynamic changes in tropical flower production. Nat. Clim. Chang. 2013, 3, 838–842. [Google Scholar] [CrossRef]
- Lebrija-Trejos, E.; Pérez-García, E.A.; Meave, J.A.; Poorter, L.; Bongers, F. Environmental changes during secondary succession in a tropical dry forest in Mexico. J. Trop. Ecol. 2011, 27, 477–489. [Google Scholar] [CrossRef] [Green Version]
- McLaren, K.P.; McDonald, M.A. Seedling dynamics after different intensities of human disturbance in a tropical dry limestone forest in Jamaica. J. Trop. Ecol. 2003, 19, 567–578. [Google Scholar] [CrossRef]
- Markesteijn, L.; Poorter, L.; Bongers, F.; Paz, H.; Sack, L. Hydraulics and life history of tropical dry forest tree species: Coordination of species’ drought and shade tolerance. New Phytol. 2011, 191, 480–495. [Google Scholar] [CrossRef]
- Seiler, C.; Hutjes, R.W.A.; Kruijt, B.; Quispe, J.; Añez, S.; Arora, V.K.; Melton, J.R.; Hickler, T.; Kabat, P. Modeling forest dynamics along climate gradients in Bolivia. J. Geophys. Res. Biogeosci. 2014, 119, 758–775. [Google Scholar] [CrossRef]
- Navarro, L.; Morin, H.; Bergeron, Y.; Girona, M.M. Changes in spatiotemporal patterns of 20th century spruce budworm outbreaks in Eastern Canadian Boreal Forests. Front. Plant Sci. 2018, 9, 1905. [Google Scholar] [CrossRef]
- Seidl, R.; Schelhaas, M.J.; Rammer, W.; Verkerk, P.J. Increasing forest disturbances in Europe and their impact on carbon storage. Nat. Clim. Chang. 2014, 4, 806. [Google Scholar] [CrossRef] [PubMed]
- Gavito, M.E.; Sandoval-Pérez, A.L.; del Castillo, K.; Cohen-Salgado, D.; Colarte-Avilés, M.E.; Mora, F.; Santibáñez-Rentería, A.; Siddique, I.; Urquijo-Ramos, C. Resilience of soil nutrient availability and organic matter decomposition to hurricane impact in a tropical dry forest ecosystem. For. Ecol. Manag. 2018, 426, 81–90. [Google Scholar] [CrossRef]
- Xie, Y.; Wang, X.; Silander, J.A. Deciduous forest responses to temperature, precipitation, and drought imply complex climate change impacts. Proc. Natl. Acad. Sci. USA 2015, 112, 13585–13590. [Google Scholar] [CrossRef] [Green Version]
- Kanniah, K.D.; Beringer, J.; Hutley, L.B. Response of savanna gross primary productivity to interannual variability in rainfall: Results of a remote sensing based light use efficiency model. Prog. Phys. Geogr. 2013, 37, 642–663. [Google Scholar] [CrossRef]
- Tapia-Palacios, M.A.; García-Suárez, O.; Sotomayor-Bonilla, J.; Silva-Magaña, M.A.; Pérez-Ortíz, G.; Espinosa-García, A.C.; Ortega-Huerta, M.A.; Díaz-Ávalos, C.; Suzán, G.; Mazari-Hiriart, M. Abiotic and biotic changes at the basin scale in a tropical dry forest landscape after Hurricanes Jova and Patricia in Jalisco, Mexico. For. Ecol. Manag. 2018, 426, 18–26. [Google Scholar] [CrossRef]
- Renton, K.; Salinas-Melgoza, A.; Rueda-Hernández, R.; Vázquez-Reyes, L.D. Differential resilience to extreme climate events of tree phenology and cavity resources in tropical dry forest: Cascading effects on a threatened species. For. Ecol. Manag. 2018, 426, 164–175. [Google Scholar] [CrossRef]
- Jimenez-Rodríguez, D.L.; Alvarez-Añorve, M.Y.; Pineda-Cortes, M.; Flores-Puerto, J.I.; Benítez-Malvido, J.; Oyama, K.; Avila-Cabadilla, L.D. Structural and functional traits predict short term response of tropical dry forests to a high intensity hurricane. For. Ecol. Manag. 2018, 426, 101–114. [Google Scholar] [CrossRef]
- Holm, J.A.; Van Bloem, S.J.; Larocque, G.R.; Shugart, H.H. Shifts in biomass and productivity for a subtropical dry forest in response to simulated elevated hurricane disturbances. Environ. Res. Lett. 2017, 12. [Google Scholar] [CrossRef]
- Middleton, B.A.; Sanchez-Rojas, E.; Suedmeyer, B.; Michels, A. Fire in a Tropical Dry Forest of Central America: A Natural Part of the Disturbance Regime? Biotropica 1997, 29, 515–517. [Google Scholar] [CrossRef]
- Otterstrom, S.M.; Schwartz, M.W.; Velázquez-Rocha, I. Responses to Fire in Selected Tropical Dry Forest Trees. Biotropica 2006, 38, 592–598. [Google Scholar] [CrossRef]
- Montoro-Girona, M.; Navarro, L.; Morin, H. A secret hidden in the sediments: Lepidoptera scales. Front. Ecol. Evol. 2018, 6. [Google Scholar] [CrossRef]
- Schwalm, C.R.; Anderegg, W.R.; Michalak, A.M.; Fisher, J.B.; Biondi, F.; Koch, G.; Litvak, M.; Ogle, K.; Shaw, J.D.; Wolf, A. Global patterns of drought recovery. Nature 2017, 548, 202. [Google Scholar] [CrossRef]
- Valdes, M.; Asbjornsen, H.; Gomez-Cardenas, M.; Juarez, M.; Vogt, K.A. Drought effects on fine-root and ectomycorrhizal-root biomass in managed Pinus oaxacana Mirov stands in Oaxaca, Mexico. Mycorrhiza 2006, 16, 117–124. [Google Scholar] [CrossRef] [PubMed]
- Pau, S.; Okin, G.S.; Gillespie, T.W. Asynchronous response of tropical forest leaf phenology to seasonal and el Nino-driven drought. PLoS ONE 2010, 5, e11325. [Google Scholar] [CrossRef]
- Bretfeld, M.; Ewers, B.E.; Hall, J.S. Plant water use responses along secondary forest succession during the 2015–2016 El Nino drought in Panama. New Phytol. 2018, 219, 885–899. [Google Scholar] [CrossRef]
- Keenan, T.F.; Hollinger, D.Y.; Bohrer, G.; Dragoni, D.; Munger, J.W.; Schmid, H.P.; Richardson, A.D. Increase in forest water-use efficiency as atmospheric carbon dioxide concentrations rise. Nature 2013, 499, 324–327. [Google Scholar] [CrossRef]
- Konings, A.G.; Gentine, P. Global variations in ecosystem-scale isohydricity. Glob. Chang. Biol. 2017, 23, 891–905. [Google Scholar] [CrossRef]
- Taylor, M.A.; Centella, A.; Charlery, J.; Bezanilla, A.; Campbell, J.; Borrajero, I.; Stephenson, T.; Nurmohamed, R. The Precis Caribbean Story: Lessons and Legacies. Bull. Am. Meteorol. Soc. 2013, 94, 1065–1073. [Google Scholar] [CrossRef]
- Coe, M.T.; Brando, P.M.; Deegan, L.A.; Macedo, M.N.; Neill, C.; Silvério, D.V. The Forests of the Amazon and Cerrado Moderate Regional Climate and Are the Key to the Future. Trop. Conserv. Sci. 2017, 10. [Google Scholar] [CrossRef] [Green Version]
- Chadwick, R.; Good, P.; Martin, G.; Rowell, D.P. Large rainfall changes consistently projected over substantial areas of tropical land. Nat. Clim. Chang. 2015, 6, 177–181. [Google Scholar] [CrossRef]
- Huang, J.; Yu, H.; Guan, X.; Wang, G.; Guo, R. Accelerated dryland expansion under climate change. Nat. Clim. Chang. 2015, 6, 166–171. [Google Scholar] [CrossRef]
- Salazar, A.; Katzfey, J.; Thatcher, M.; Syktus, J.; Wong, K.; McAlpine, C. Deforestation changes land–atmosphere interactions across South American biomes. Glob. Planet. Chang. 2016, 139, 97–108. [Google Scholar] [CrossRef]
- Esquivel-Hernández, G.; Sánchez-Murillo, R.; Birkel, C.; Good, S.P.; Boll, J. Hydroclimatic and ecohydrological resistance/resilience conditions across tropical biomes of Costa Rica. Ecohydrology 2017, 10. [Google Scholar] [CrossRef]




© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stan, K.; Sanchez-Azofeifa, A. Tropical Dry Forest Diversity, Climatic Response, and Resilience in a Changing Climate. Forests 2019, 10, 443. https://doi.org/10.3390/f10050443
Stan K, Sanchez-Azofeifa A. Tropical Dry Forest Diversity, Climatic Response, and Resilience in a Changing Climate. Forests. 2019; 10(5):443. https://doi.org/10.3390/f10050443
Chicago/Turabian StyleStan, Kayla, and Arturo Sanchez-Azofeifa. 2019. "Tropical Dry Forest Diversity, Climatic Response, and Resilience in a Changing Climate" Forests 10, no. 5: 443. https://doi.org/10.3390/f10050443
APA StyleStan, K., & Sanchez-Azofeifa, A. (2019). Tropical Dry Forest Diversity, Climatic Response, and Resilience in a Changing Climate. Forests, 10(5), 443. https://doi.org/10.3390/f10050443